Introduction
Horse flies and other tabanids (Diptera: Tabanidae) affect the health of livestock throughout the world as mechanical vectors of diseases such as surra, caused by Trypanosoma evansi, and nagana, caused by Trypanosoma vivax (Krinsky, Reference Krinsky1976; Foil, Reference Foil1989; Desquesnes, Reference Desquesnes2004); but their most important impact is often through the act of feeding. Blood loss and irritation can result in significant losses in productivity, or even in death. For example, as few as 66 horse flies feeding per day on heifers in the USA reduced weight gain by 0.1 kg per day (Perich et al., Reference Perich, Wright and McNew1986). The economic impact of tabanids is particularly important in the Sudan where tabanids emerge in extremely high numbers during the rainy season (Mohamed-Ahmed et al., Reference Mohamed-Ahmed, Abdulla, Mohamed, El Rayah and El Amin2007). In the central Sudan along the White Nile (personal observation) and in the Bahr El Arab tsetse belt, Southern Darfur, Western Sudan (Hall et al., Reference Hall, Kheir, Rahman and Noga1984), tabanid attacks on cattle reach 10–50 times the levels typical of temperate environments (Abdel Karim & Benjamin, Reference Abdel Karim and Benjamin1989). The impacts of the high levels of tabanid challenge in the Sudan have rarely been assessed (Rahman, Reference Rahman2005).
The range of options for control of horse flies is limited. Large-scale control is rarely practical, as most tabanids are mobile and widely-distributed (Anderson, Reference Anderson1985). However, on a local scale, removal/kill trapping, especially with the use of baits (Vale & Torr, Reference Vale, Torr, Maudlin, Holmes and Miles2004), has proven to be effective under special circumstances (Wall & Doane, Reference Wall and Doane1980). Although currently used only for tsetse control, insecticide-treated visual decoys and/or cattle show potential for other biting flies, with exploratory studies underway with stable flies (Hogsette et al., Reference Hogsette, Nalli and Foil2008). There is a similar renewed interest in the use of bait technology for mosquito control (Kline, Reference Kline2006; Torr et al., Reference Torr, Della Torre, Calzetta, Costantini and Vale2008).
In the Sudan, there are 74 species of tabanids (Yagi, Reference Yagi1968). Tabanids such as Philoliche spp. (e.g. P. magretti Bezzi) and certain Tabanus spp. (e.g. T. biguttatus Wiedemann) are difficult to catch in traps (Mihok, Reference Mihok2002; Abdulla & El Malik, Reference Abdulla and El Malik2003; Mohamed-Ahmed et al., Reference Mohamed-Ahmed, Abdulla, Mohamed, El Rayah and El Amin2007). Until better traps and baits are developed, insecticide-treated devices and/or hosts may be the only practical control methods for many tabanid species. A pre-requisite to the economical use of this technology is the determination of behavioural responses to hosts and their mimics (Torr et al., Reference Torr, Maudlin and Vale2007a).
This paper describes, for the first time, the alighting sites of Sudanese tabanids on cows, calves and goats, as well as on cylindrical models mimicking hosts. We also address the behaviour of non-biting muscids, which are of interest because of the use of aerial fogging with toxic insecticides in cities for ‘fly’ control. Determination of alighting sites should also be useful for the refinement of traps for monitoring and/or control of all biting flies (Mihok, Reference Mihok2002). An example of an existing practical control device for tabanids is the greenhead box trap (Wall & Doane, Reference Wall and Doane1980). It has been used for many years to control tabanids near Cape Cod, USA (Hayes et al., Reference Hayes, Doane, Sakolsky and Berrick1993). Its design relies on flies entering the bottom of an elevated trap, exploiting the behaviour of Tabanus nigrovittatus Macquart, a species which feeds on the undersides of cattle (Allan et al., Reference Allan, Day and Edman1987).
Materials and methods
Study site
The study was performed over two rainy seasons (June–November 2006–2007) at the fodder plots of the Farm Unit (26 ha) of the College of Veterinary Medicine and Animal Production, Sudan University of Science and Technology, Khartoum (15.5°N, 32.5°E). Irrigation canals provide moist soil for tabanid larvae; and resident farm animals (91 cattle, 21 sheep, 18 goats, 3 camels, 4 horses and 5 donkeys) provide hosts for a limited fauna consisting primarily of Tabanus sufis (Jennicke), Atylotus agrestis (Wiedemann) and Tabanus taeniola (Palisot). Catches in Nzi traps of 50–150 tabanids/trap/day are typical of peak abundance during rainy seasons (Mohamed-Ahmed et al., Reference Mohamed-Ahmed, Abdulla, Mohamed, El Rayah and El Amin2007, unpublished data). Evergreen shade trees and flower and grass beds provide nectar, resting sites and refuges in the extreme climate of this desert environment.
Alighting on models
Tabanids and non-biting muscids (Muscinae, Stomoxys were almost absent) were counted after they were stuck, killed or stunned as they alighted on sticky or electrified cylindrical models mimicking hosts (Vale, Reference Vale1974). Three models were used: small (‘goat’, 73-cm long, 27-cm Ø), medium (‘calf’, 65-cm long, 44-cm Ø) and large (‘cow’, 95-cm long, 58-cm Ø). The large model was 6.01 times the small model by volume, and the medium model was 2.36 times larger. Each model consisted of a plastic drum wrapped in matt-black cotton cloth. A model stood horizontally on four legs of flat iron bars 1.5-cm wide, 0.2-cm thick and 37- or 11-cm high, depending on the experiment. An electrified model was built by attachment of an electrocution grid (Vale, Reference Vale1974) to one half or all of the circumference of the drum, depending on the experiment, excluding the anterior and the posterior faces. It was powered by a high voltage transformer that converted 12 v DC to ~40,000 v interrupted DC (Bonar Industries, Zimbabwe). Stunned or killed flies were collected in shallow brown trays containing soapy water (180-cm long, 120-cm wide and 5-cm deep). Each model was placed in the middle of a tray leaving a clear space on all sides. Sticky models were constructed by completely wrapping the legs and torso of each model in transparent adhesive film used in insect electrocution devices (Stock No. FE26, Rentokil Initial PLC, Felcourt, UK), leaving the faces uncovered as with electrocuting grids. Flies were recovered with forceps, identified and assigned to where they had alighted: legs, upper or lower ‘torso’. When the circumference of the drum was enclosed with an electrocution grid or sticky film, the large model had 2.80 times the ‘catching’ area of the small model, and the medium model had 1.45 times the area, disregarding the small area of the legs (see dimensions of the leg above) covered by adhesive film. Model experiments were performed between 15:00–18:00 h to coincide with the single daily peak of tabanid activity and the second daily peak of muscid activity during the rainy season (Mohamed-Ahmed, unpublished data). Stuck and killed/stunned flies were identified and counted at the end of each daily run.
The use of adhesive film to monitor biting fly behaviour in place of electrocuting grids has only recently been reported. We, therefore, present complimentary data for an additional simultaneous comparison that was not included in table 2 of Mihok et al. (Reference Mihok, Carlson and Ndegwa2007) to provide further information on relative capture efficiency for this approach. At Nguruman, Kenya, a 3-D triangular target with 1×1 m sides was constructed by wrapping blue cloth around three poles. The sides were covered with an electrocuting grid or adhesive film, or an electrocuting grid in front of adhesive film, to catch flies alighting on the target. Equipment and general methods were identical to those used in the Sudan. The target was baited with acetone, cow urine and octenol. It was set for three days with the capture methods rotated among the three sides at the end of each day. Catches were tallied three times a day from sunrise to just after sunset for a total of N=9 per treatment.
Alighting on hosts
A brown cow (300 kg), a black female calf (100 kg) and a black female goat (30 kg) were used in three consecutive experiments run for six days at the same site (mean Nzi trap catch was 35 tabanids per trap per day at this time: Mohamed-Ahmed et al., Reference Mohamed-Ahmed, Abdulla, Mohamed, El Rayah and El Amin2007). The animals were tethered but could feed, groom and move over a radius of 2 m. They were given water and green Durrah stalks (Sorghum spp.). Two observers stood about 3 m away on opposite sides to score alighting insects as well as behaviour between 16:00–18:00 h. Close proximity of humans does not appear to influence the behaviour of African tabanids near animals (Vale & Phelps, Reference Vale and Phelps1974), but effects have been seen with North American species (Hribar et al., Reference Hribar, Leprince and Foil1992). During our observations, only two T. sufis approached or landed on the observers. To score alighting sites, the body was divided into three regions: upper torso (including the head, neck with dewlap, upper flank and back), lower torso (including the lower flank and belly) and legs (including the fore- and hind-quarters). If a single fly was seen to alight more than once, it was counted each time.
Five types of host defensive behaviour were recorded (Ralley et al., Reference Ralley, Galloway and Crow1993): (i) tail lashes (disturbing flies with the tail), (ii) head tosses and licks (using the head or tongue to dislodge flies), (iii) ear flicks (movement of the ear to disturb alighting flies), (iv) skin rippling (the characteristic twitching of the skin in response to attacking flies), and (v) foot stomping (kicking and violent movement of legs and hooves to dislodge flies)
Experimental designs and data analysis
Randomized Latin squares incorporating the effects of treatment, sites and days (3×3) were used for model experiments. Each 3×3 square was replicated twice on consecutive three-day periods within experiments. Means were separated using the Student-Newman-Keuls (SNK) test at P<0.05. Relative catches are expressed as a catch index, the backtransformed mean catch relative to the small model for size comparisons, or relative to the upper torso for area comparisons. The distribution of the landing patterns of flies on different regions of hosts was straightforward and was, therefore, characterized by χ2 analysis.
Results
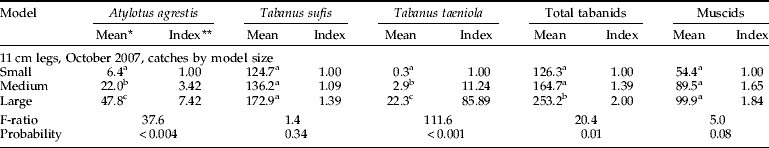
The models caught many female tabanids and non-biting muscids, along with a few male tabanids. Female T. sufis comprised 94.2% (N=231), 95.2% (N=1064), 81.9% (N=477) of the tabanids caught by electrocution in experiments 1–3, and 82.1% (N=3924) of those caught with sticky models in experiment 4. Tabanus taeniola and A. agrestis accounted for the rest of the female tabanid catch. Muscinae accounted for 82.9, 46.8, 62.6 and 29.6% of the total catches. Only one male T. taeniola was caught on the sticky models, versus 22–68 male tabanids on the electrified models (114 of 1886). In experiments 1–3 (table 1a–c), 24, 22 and 68 male tabanids were caught with T. taeniola representing 91, 73 and 92% of the males, respectively. Alighting of male tabanids (F=0.023–1.55; P=0.31–0.98), and Muscinae (table 1a–c and 2) did not vary significantly with model size.
Table 1. Alighting of Tabanidae and non-biting Muscinae on electrified models.

* Back-transformed values; means in the same column with different superscripts are significantly different at P<0.05, SNK test; ** ratio of back-transformed mean relative to the small model in (a) and (b), and relative to the model with the upper half covered in an electrocuting grid in (c).
Alighting on electrified models
In the first experiment, with models standing on legs 37-cm high and the electrocution grid only on the upper half, the large model caught 2.57 times as many alighting T. sufis and 2.74 times as many total tabanids as the small one, respectively (table 1a). Catches were always greatest with the large model, but variability was too high to show whether there was a graduated effect of model size. Many tabanids were seen landing on the legs and may not have been counted by the upper grid, so the second experiment was conducted with shorter legs (11 cm), again with the grid covering only the upper half. With shorter legs, tabanid catches increased by model size, e.g. the large model caught 4.11 and 2.04 times as many T. sufis, and 4.15 and 1.99 times as many total tabanids, as the small model (table 1b). In both experiments, very few tabanids other than T. sufis were caught.
To investigate what proportion of tabanids landing on a model were being electrocuted by a partial electrocution grid, the third experiment was performed using three identical medium-sized models each standing on legs 11-cm high. One model was covered with the grid on the upper half only (upper torso), the second was covered with the grid on the lower half only (lower torso) and the third was covered with the grid over the entire circumference. Table 1c shows that the model with the grid on the upper half only (as in prior experiments) caught significantly fewer T. sufis and total tabanids than when the lower half or the entire circumference was electrified. Catches implied that about two-thirds of the tabanids landing on the medium model were not being counted in the first two experiments, as many flies were landing on the lower torso only. As noted earlier, this result was unique to tabanids, since neither model size nor the extent of the grid had significant effects on muscid catch. This implied that muscids often alighted more than once, sometimes choosing to land on different regions of the models.
Alighting on sticky models
Sticky models were used to better document total numbers and alighting sites, taking into account flies landing on the support legs. With the circumference and all the legs covered with adhesive film, significantly more total tabanids alighted on the large model than on both the small or medium models; and species other than T. sufis were better represented in the catch (table 2). More A. agrestis and T. taeniola landed on large vs. medium vs. small models, and similar numbers of T. sufis landed on all three models.
Table 2. Alighting of Tabanidae and non-biting Muscinae on models with the circumference and legs covered in clear adhesive film.
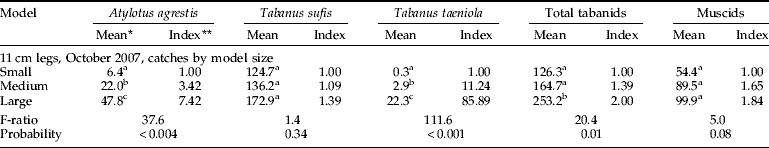
* Back-transformed values; means in the same column with different superscripts are significantly different at P<0.05, SNK test; ** ratio of back-transformed mean relative to the small model.
To identify the origins of different patterns among species, catches on the legs, and the lower or upper torso, were compared separately among models. Altogether, 2315 tabanids landed on the legs of the three models, consisting of 99.3% T. sufis (fig. 1). Landings of T. sufis on the legs of models did not vary with model size (data not shown). In contrast, alighting of tabanids increased significantly with model size for both the lower and upper torso (figs 2 and 3). For the lower torso, significantly more tabanids of all species landed on the large model relative to the small and medium models, with more landings of A. agrestis also on the medium vs. the small model. For the upper torso, significantly more T. sufis and T. taeniola landed on the large model relative to the small and medium models; landings of A. agrestis did not differ by model size. Muscinae strongly preferred to land first on the upper torso (fig. 3 vs. fig. 2), but there was no significant pattern according to model size.

Fig. 1. Flies (mostly Tabanus sufis) stuck to clear adhesive film wrapped around the short flat metal legs of a drum model.

Fig. 2. Distribution of flies on the lower torso of different sizes of three sticky models. Letters within species/groups differentiate significantly different catches, P<0.05, SNK test (□, small; ![]() , medium; ▪, large).
, medium; ▪, large).

Fig. 3. Distribution of flies on the upper torso of different sizes of three sticky models. Letters within species/groups differentiate significantly different catches, P<0.05, SNK test (□, small; ![]() , medium; ▪, large).
, medium; ▪, large).
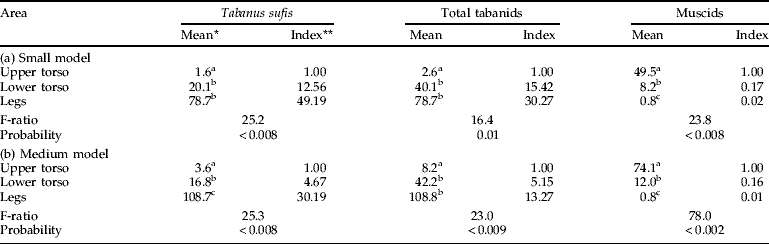
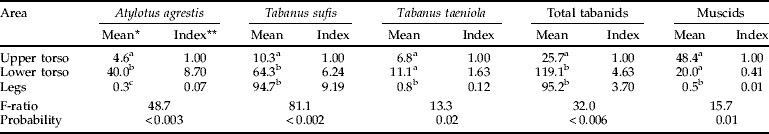
In addition to patterns related to model size, tabanids showed strong preferences for landing on certain regions of any one model. T. sufis preferred to land on the legs (49×, 30×, 9× by increasing model size relative to the upper torso) or on the lower torso (tables 3 and 4). Muscids showed a strong bias for the upper torso, regardless of model size, with hardly any captures on the legs. For the large model where captures of A. agrestis and T. taeniola were adequate for statistical analysis (table 4), there were 133.3 times as many A. agrestis and 13.9 times as many T. taeniola captured on the lower torso than on the legs. A. agrestis also showed a strong preference for the lower vs. the upper torso (8.7×); catches of T. taeniola were similar in these two regions.
Table 3. Alighting of Tabanidae and non-biting Muscinae on different areas of small and medium models with the circumference and legs covered in clear adhesive film (October 2007, 11 cm legs).
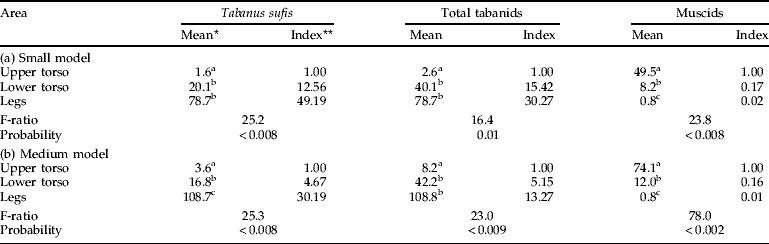
* Back-transformed values; means in the same column with different superscripts are significantly different at P<0.05, SNK test; ** ratio of back-transformed mean relative to the upper torso.
Table 4. Alighting of Tabanidae and non-biting Muscinae on different areas the large model with the circumference and legs covered in clear adhesive film (October 2007, 11 cm legs).
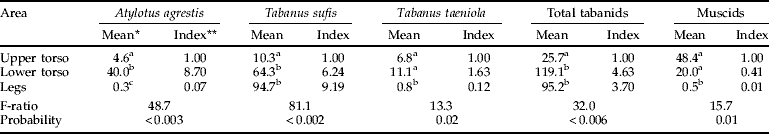
* Back-transformed values; means in the same column with different superscripts are significantly different at P<0.05, SNK test; ** ratio of back-transformed mean relative to the upper torso.
Relative efficiency of sticky materials and electrocuting grids
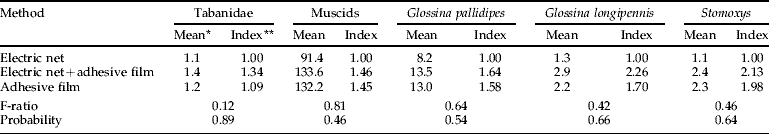
Our attempt to directly compare capture techniques simultaneously for various Diptera in Kenya was only partially successful (table 5). Only muscids and Glossina pallidipes Austen were present in good numbers at the site where this experiment was conducted. Catches of all types of flies were statistically equal among capture methods, but with an indication of higher captures with adhesive film. This trend was present whether the film was used to capture flies or was simply present behind the electrocuting grid. No flies were caught on the adhesive film placed behind the electrocuting grid.
Table 5. Simultaneous comparison of three techniques for enumerating biting flies alighting on the 1×1 m sides of a triangular trap model made from blue cloth wrapped around three poles (June 1996, Nguruman, Kenya).
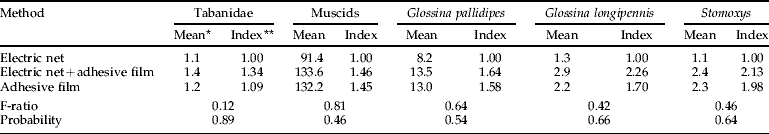
* Back-transformed values; ratio of back-transformed mean relative to an electric net alone.
Alighting on hosts
Results for muscids were straightforward with a strong bias for landing on the upper torso of all hosts (goat: χ2=107.2, df=1, P≪0.0001; calf: χ2=1157.5, df=2, P≪0.0001; cow: χ2=386.6, df=2, P≪0.0001); this pattern was also typical of muscid behaviour towards the equivalent models (table 6). For the largest sample from the cow (N=3135), the landing behaviour of each of the three tabanid species was not random (χ2=126.8–2355.1, df=2, P≪0.0001) but similar. About twice as many tabanids landed on the legs as on the lower torso, with very few landings on the upper torso (table 6). This pattern was typical of T. sufis on all models, but was the reverse of the pattern observed for T. taeniola and A. agrestis on models (preference for landing on the lower torso). Non-random landing of tabanids was also observed on the goat and the calf (goat: χ2=31.2, df=1, P≪0.0001; calf: χ2=132.8, df=2, P≪0.0001), but sample sizes were too small to differentiate between species. Altogether, 90–98% and 94–100% of T. sufis alighted on the legs and the lower torsos of the models and hosts, respectively. Atylotus agrestis and T. taeniola hardly alighted on the legs of models, but 53–100%, 89–100% and 54–69% preferred the legs of the goat, calf and cow, respectively.
Table 6. Percent alighting of Tabanidae and non-biting Muscinae on different areas of live hosts compared with alighting on corresponding sticky models of similar size (2006–2007).

Defensive response of hosts
From the number of landings (table 6), tabanid challenge was 34.9 times greater on the cow than on the calf, and 69.3 times greater on the cow than on the goat. The larger tabanids (T. taeniola, A. agrestis) accounted for a greater proportion of observations on the cow (81.3%) than on the calf (50.0%) or goat (54.5%). Each animal defended itself against alighting flies by one or more actions, such as tail lashes, skin ripples (111.5 per day), foot stomps (99.0 per day), ear flicks (78.4 per day) or head tosses (4.0 per day). Tail lashes were often continuous.
Since very few tabanids landed on the calf or the goat, correlations between the frequency of each response (y) and the total of landing flies (x) were explored for the cow only (N=6). A significant positive correlation was found only between the log-transformed number of foot stomps, and the log-transformed number of alighting tabanids as a whole (log y=–0.3+1.6 log x, R=0.98, df=4, P<0.02).
Discussion
Models
The harassment of equines and camels by tabanids was described by early explorers in Africa. In modern times, several studies of alighting behaviour on livestock have been conducted, particularly in North America (Hollander & Wright, Reference Hollander and Wright1980; Magnarelli & Anderson, Reference Magnarelli and Anderson1980; Zyzak et al., Reference Zyzak, Leprince, Byford and Foil1989; Foil et al., Reference Foil, Leprince and Byford1990; Phelps & Holloway, Reference Phelps and Holloway1990; Hribar et al., Reference Hribar, Leprince and Foil1992; Krčmar & Durbesic, Reference Krčmar and Durbesic2000). The documentation of alighting sites is important for the economical use of residual insecticides on hosts or their mimics, given the paucity of options for control of biting flies (Hall & Wall, Reference Hall, Wall, Maudlin, Holmes and Miles2004).
Our most practical finding was that more tabanids, but not more muscids, landed on larger models when set at a low height (tables 1 and 2). Very many more tabanids also landed on the largest animal (the cow; table 6). This agrees with the attraction of diverse tabanids to large animals/objects (Gibson & Torr, Reference Gibson and Torr1999) although certain species specialize in feeding on other hosts (Ferreira et al., Reference Ferreira, Henriques and Rafael2002; Ferreira & Rafael, Reference Ferreira and Rafael2004). Vale (Reference Vale1974) was the first African researcher to explore the use of a drum model (mimic of an adult warthog standing 37 cm high) to concentrate tsetse flies around an electrocuting grid for behavioural studies. Subsequent tests of different-sized models revealed that tsetse landings increased with increasing model size (Hargrove, Reference Hargrove1980). In our study, this was only partly true for tabanids (table 1a,b) and did not apply to muscids. Muscinae and Stomoxyinae explore the outside and top of objects much more than other flies (Vale, Reference Vale1982a,Reference Valeb). To reduce catches of non-target species, early traps for tsetse were designed with basal entrances and a large cone. Unfortunately, very few insights were obtained for tabanids in these elegant studies.
The numbers of landing tabanids were affected by height over a narrow range (11 vs. 37 cm), with greater catches on larger models unequivocal only at the lower height (table 1b). This appeared to be due to the preference of T. sufis to land on model legs and/or the lower torso. It was the main species captured in the first experiments with a partial electrocuting grid when models with legs of different sizes were tested. For other species, the size of the model was critical for obtaining significant captures, e.g. the large sticky model set at the lower height caught 86 times as many T. taeniola as the small model (table 2).
Estimates of tabanids alighting on models were affected by the capture technique, with significantly different estimates obtained from partial vs. complete electrocution grids (table 1c). Substantially higher captures were obtained with the use of adhesive film. The effects of model size and the distribution of landings also differed between tabanid species. Responses for each species were clear but were not a simple function of model volume or surface area, as with tsetse such as Glossina pallidipes Austen and Glossina morsitans morsitans Westwood (Hargrove, Reference Hargrove1980)
Our experimental design did not provide information on tabanids approaching the models and not landing, a critical factor in the efficacy of insecticide-impregnated devices (Gibson & Torr, Reference Gibson and Torr1999). Circling without landing is common in tsetse; the frequency of landing also varies greatly among species in the presence or absence of host odours (Hargrove, Reference Hargrove1980). This may also occur with certain tabanids, e.g. as shown by the striking increase in catch of Croatian tabanids when a presumably inefficient trap is baited with animal urine (Krčmar et al., Reference Krčmar, Mikuska and Merdić2006). Unfortunately, capture efficiency for tabanids has rarely been measured. Recently, Mohamed-Ahmed et al. (Reference Mohamed-Ahmed, Abdulla, Mohamed, El Rayah and El Amin2007) estimated Nzi trap efficiency with flanking electric nets for three species of Sudanese tabanids at 91% when traps were baited with a blend of phenols and octenol and 45% when unbaited (assuming 100% electrocution efficiency). Past estimates for tabanids caught while testing a historical tsetse trap were very low (1, 2, 11% for the A7C trap for three Tabanus spp. in Zimbabwe). Efficiency estimates were greater (16, 33, 18%, respectively) when a net was placed close to an object like our small/medium model (Vale & Hargrove, Reference Vale and Hargrove1979). Efficiency estimates for tsetse increase as nets are placed closer to objects, complicating comparisons among studies (Mihok et al., Reference Mihok, Carlson and Ndegwa2007).
Unfortunately, the present experiments in the Sudan did not include a simultaneous comparison of electrocution and sticky techniques to differentiate unequivocally the causes of the striking difference in the number of tabanids counted by alternative methods. Altogether, the total catch of tabanids on the adhesive film in one experiment was more than two times the total catch in three experiments using electrified models (3924 vs. 1772); in contrast, catches of muscids were similar across experiments. From seasonal monitoring with traps at the Farm Unit (Mohamed-Ahmed et al., Reference Mohamed-Ahmed, Abdulla, Mohamed, El Rayah and El Amin2007, unpublished data), the striking difference for tabanids was not a simple function of the timing of experiments. The nature of this result, nevertheless, needs to be tested explicitly with a simultaneous comparison of methods, as in our modest attempt to do so in Kenya. In any future comparison, an important issue that must be addressed is the finding that electric grids only stun a proportion of large flies such as tsetse (e.g. ~50%) (Packer & Brady, Reference Packer and Brady1990; Griffiths & Brady, Reference Griffiths and Brady1994; Dransfield & Brightwell, Reference Dransfield and Brightwell2001). This would be consistent with our results for tabanids (large) vs. muscids (small).
Carefully-controlled comparisons of techniques are also needed to rule out the possibility that the adhesive film is attractive to certain species because of the presence of subtle visual cues. This was implied by higher catches of several types of Diptera with adhesive film present in the comparison of techniques in Kenya. Tabanids are particularly sensitive to the specular reflectance of polarized light (Kriska et al., Reference Kriska, Majer, Horváth, Szivák and Horváth2008), and also to ultraviolet reflectance (Hribar et al., Reference Hribar, LePrince and Foil1991). The Rentokil product, like other sticky materials, is shiny and not perfectly transparent; it also contains an ultraviolet absorbent. Simultaneous comparisons of adhesives with different optical properties (IAEA, 2003) vs. ‘invisible’ electrocution grids are also needed. For example, Mihok et al. (Reference Mihok, Carlson and Ndegwa2007) found that transparent ‘enclosures’ of Rentokil adhesive film caught modest numbers of Tabanus quinquevittatus Wiedemann and Stomoxys calcitrans L. in Canada, whereas enclosures were largely ignored by two species of tsetse in Kenya, so long as they were unbaited. In contrast, Stomoxys niger niger Macquart were caught in large numbers, implying straightforward visual detection of the adhesive film.
The near absence of male tabanids in sticky model catches, relative to catches of 6% males on electrified models agrees with similar sticky catch results in Canada (Mihok et al., Reference Mihok, Carlson and Ndegwa2007). The difference with an electrocution approach may have been a function of using trays containing water to retain flies in this hot, desert environment. Male tabanids are attracted to pools of water; a behaviour that can be exploited when objects with special features are set on the ground (Hall et al., Reference Hall, Farkas and Chainey1998; Kriska et al., Reference Kriska, Majer, Horváth, Szivák and Horváth2008). For example, lower catches of males (~2% for T. taeniola and T. pertinens Austen) were obtained with electric nets (1.4×1.5 m) placed next to a baited model in Zimbabwe equipped with underground hoppers instead of water-filled trays (Hargrove & Vale, Reference Hargrove and Vale1978). In contrast, use of several large electric nets (1.5×3.3 m) downwind from a baited trap resulted in catches of many male T. pertinens (42%) but few male T. taeniola (4%) (Vale & Hargrove, Reference Vale and Hargrove1979). Opposite trends (2% male T. pertinens, 63% male T. taeniola) were obtained when ranging tabanids were intercepted far from an odour source by very large electric nets (9.9×1.5 m) (Phelps & Vale, Reference Phelps and Vale1976).
Hosts vs. models
In this initial study, we did not attempt to determine the absolute numbers of flies alighting on hosts. Some flies surely landed repeatedly on each host during the course of a day and were counted more than once. In future work, behaviour around hosts should be characterized more precisely by surrounding animals with an incomplete ring of electric nets (Torr, Reference Torr1994; Baylis, Reference Baylis1997; Torr & Hargrove, Reference Torr and Hargrove1998). Data are also needed on feeding success and spatial transfer among animals (Barros & Foil, Reference Barros and Foil2007), given the key role of interrupted feeding in the mechanical transmission of trypanosomes (Desquesnes & Dia, Reference Desquesnes and Dia2003).
Irrespective of tabanid species or bait type (hosts, sticky or electrified models), more than 80% of horse flies observed landing on animals or models did so on the lower torsos or lower portions (table 6). A preference for low landing sites is common in many, but not all, species of tabanids in Zimbabwe (Phelps & Holloway, Reference Phelps and Holloway1990), in Croatia (Krčmar & Durbesic, Reference Krčmar and Durbesic2000) and in the USA (Foil et al., Reference Foil, Leprince and Byford1990). Here, the smallest species T. sufis (8–11 mm) alighted mainly on the legs of baits, whereas the larger species T. taeniola (16 mm) and A. agrestis (14 mm) landed on the legs, belly, flanks and on the back. This is similar to the behaviour of horse flies in several studies in the USA; e.g. the smallest species most often attack the legs, flanks or lower body, whereas very large species (e.g. Tabanus. americanus Forster, Tabanus atratus Fabricius, 20–27 mm) land almost exclusively on the back (Mullens & Gerhardt, Reference Mullens and Gerhardt1979; Hollander & Wright, Reference Hollander and Wright1980; Hribar et al., Reference Hribar, Leprince and Foil1992). In contrast, deer flies (Chrysops spp.), which are small (5–12 mm), attack the head; this is also found in some smaller horse flies, such as the important pest T. lineola Fabricius (10–15 mm: Mullens & Gerhardt, Reference Mullens and Gerhardt1979). Altogether, the heights at which tabanids alight on hosts appears to be governed mostly by fly size rather than taxonomic relationships (Konstantinov, Reference Konstantinov1993).
One important feature of our results for targeted application of insecticides was the finding that T. sufis landed on the legs of both models and hosts, whereas A. agrestis and T. taeniola scarcely alighted on the legs of models, despite landing on the legs of all sizes of hosts. For these tabanids, differences in behaviour may have been related to the visual aspects of the iron legs, or the absence of host cues such as movement or warmth (Thorsteinson, Reference Thorsteinson1958), or host odours (Gibson & Torr, Reference Gibson and Torr1999). Alternatives beyond the use of nominal ‘legs’ may be worth investigating as large changes in catch could result from small changes in format. Similarly, although we detected few tabanids landing on the heads of cattle, it would be prudent to test models with artificial heads in other settings. Some tabanid species are attracted to the head; this also occurs with some tsetse (Torr, Reference Torr1994).
Within the fly community in our study, there was a straightforward altitudinal partitioning of flies: muscids nearly always at the top, T. taeniola and A. agrestis in the middle, and T. sufis at the bottom and on the legs. Several authors have suggested that competition among blood-sucking flies may lead to partitioning of flies on the host (Mullens & Gerhardt, Reference Mullens and Gerhardt1979; Phelps & Holloway, Reference Phelps and Holloway1990; Hribar et al., Reference Hribar, Leprince and Foil1992). Mullens & Gerhardt (Reference Mullens and Gerhardt1979) and Hribar et al. (Reference Hribar, Leprince and Foil1992) also observed more horse flies at the anterior compared with the posterior of grazing cattle, possibly related to tail lashing. Waage & Davies (Reference Waage and Davies1986) did not agree with a competition-based interpretation as they believed resources such as blood and space are not limiting. This opinion was also expressed by Hollander & Wright (Reference Hollander and Wright1980), who noted that the density of tabanids on hosts did not affect additional landings. Waage & Davies (Reference Waage and Davies1986) proposed that density-dependant grooming could influence partitioning, e.g. by reducing feeding success of timid species in body regions where they overlap with aggressive species. As in other studies, our animals reacted to alighting flies with simple defensive behaviours. High levels of attack may also lead to bunching in herds (Ralley et al., Reference Ralley, Galloway and Crow1993; Torr, Reference Torr1994; Torr et al., Reference Torr, Prior, Wilson and Schofield2007b). In our brief study, we found only one significant correlation between foot stomps of the cow and the frequency of alighting of tabanids. Defensive responses are clearly more complicated, but can only be quantified with a much more sophisticated effort (Schofield & Torr, Reference Schofield and Torr2002).
Conclusions
Our objective was to document the alighting of tabanids and muscids on practical models relative to hosts to guide the design of insecticide-treated devices for biting fly control on a local scale in the Sudan. Large-scale control may never be practical, given the failure of deltamethrin-impregnated, odour-baited, 2-D cloth targets to reduce numbers of tabanids in a major tsetse control trial in Zimbabwe (Vale et al., Reference Vale, Lovemore, Flint and Cockbill1988). If bait technology is to be pursued, e.g. for high-value operations such as periurban dairies supplying cities, pyrethroid-treated, 3-D black drum models would be appropriate based on our results.
An alternative strategy, the use of insecticide-treated cattle, has also been suggested as a practical option for tabanid control (Hall & Wall, Reference Hall, Wall, Maudlin, Holmes and Miles2004). This conclusion has some support in reduced catches of biting flies in a natural setting following the application of cypermethrin pour-on to cattle in Ethiopia (Leak et al., Reference Leak, Mulatu, Rowlands and D‘Ieteren1995). It is popular for tsetse control, but there are serious concerns about its sustainability (Vale et al., Reference Vale, Mutika and Lovemore1999). Over-reliance on this approach has the following problems: (i) insecticide treatment of cattle may interfere with natural resistance to ticks and tick-borne diseases, (ii) grazing and watering practices may limit contact between treated herds and areas of high fly numbers, and (iii) maintaining residual mortality may prove to be uneconomical in bright sun or heavy rain, or simply as a result of animals grazing in tall, wet grass. Issues of insecticide persistence are a major problem in the tropics and are particularly relevant for any effort to control tabanids in the Sudan. Harsh sunlight and high temperatures characterize Sudanese weather; heavy rain also coincides with high fly emergence. During certain weather conditions, insecticide persistence on treated cattle may be as short as five days, making this strategy uneconomical (Vale et al., Reference Vale, Mutika and Lovemore1999).
Although there are clearly pitfalls in using long-lasting insecticides and baits for biting fly control, we believe that sufficient progress has now been made on the behaviour of biting flies to embark on pilot trials based on the lessons learned from tsetse.
Acknowledgements
We wish to thank our field assistants and the undergraduate students of the College of Veterinary Medicine, Sudan University, who performed some of the experiments as part of their graduate programmes. The work was funded by the Sudan University and the Ministry of Higher Education and Scientific Research, Khartoum, Sudan.