Introduction
One of the most commonly asked questions is ‘Where are the aliens?’ If there are so many planets and the Universe is 14.3 billion years old, why have no aliens popped over to the Earth to investigate it or to colonize it (Jasty, Reference Jasty2014)? A simple solution might be that the time that has been available thus far is not sufficient to allow the development of complex species capable of space travel (Loeb et al., Reference Loeb, Batista and Sloan2016). More complex solutions involve the nature of planets upon which life can evolve. For example, Lingam and Loeb have modelled the habitability of planets that are tidally locked to their parent stars (Lingam and Loeb, Reference Lingam and Loeb2017). In this analysis, the authors show that when atmospheric erosion is taken into account, both the TRAPPIST-1 planets and Proxima b are in fact relatively poor candidates for life. Other studies suggest much the same at least for a proportion of planets in the habitable zones of their red dwarf hosts (Irwin et al., Reference Irwin, Charbonneau, Nutzman and Falco2008; Luger and Barnes, Reference Luger and Barnes2015; Ribas et al., Reference Ribas, Bolmont, Selsis, Reiners, Leconte, Raymond, Engle, Guinan, Morin, Turbet, Forget, Velasco and Anglada-Escudé2016; Wheatley et al., Reference Wheatley, Louden, Bourrier, Ehrenreich and Gillon2016; Barnes, Reference Barnes2017).
In this paper, we also consider the habitability of tidally locked planets and show that a key limiting factor is the functional surface area upon which life can evolve. This functional space is not the total surface area of the planet, but the area over which climatic and other factors are constrained by illumination angle and continental surface. The habitable space can be defined in terms of a hypervolume – a term which allows a simple integration of the principle factors controlling biological diversity on Earth (Gatti, Reference Gatti2016a, Reference Gatti2016b; Blonder, Reference Blonder2017).
In essence, while tidally locked planets may be habitable in terms of their atmospheric structure, density, temperature and availability of water, they are far more limited in terms of their developmental capacity; that is, their ability to host and evolve complex life from simpler forms. By integrating plate tectonics into the ‘construction’ of model worlds, we can see how this process, more than any other, may limit the development of biological complexity.
Methods
We assume that the gross habitable surface corresponds to those regions that are illuminated in the first instance. While this is not a true measure of habitability in terms of all life, it does delineate the area over which complex life is probable, as complex life will require an abundance of energy supplied by the host star. This area is defined by the simple relationship shown in equation 1:
 $$A = 2{\rm \pi }r^2.$$
$$A = 2{\rm \pi }r^2.$$The habitable surface in this instance is half the total surface area of the planet, because half the planet is in perpetual darkness and any biosphere will be subsurface, where geothermal energy supports any food chains. If we use the Earth as an analogue, then we assume that limited microbial food chains will be possible in darkness if other sources of energy are available, but a lack of light, perpetual low temperatures and limited liquid water will preclude biological complexity on land.
For slow, but synchronously rotating, planets (those with orbital periods exceeding than 20 days (Wolf, Reference Wolf2017; Wolf et al., Reference Wolf, Shields, Kopparapu, Haqq-Misra and Toon2017)), the habitable area is defined by the surface area subtended by angle θ around the Sub-Stellar Point (SSP). This is shown by equation 2:
 $$ A = 2{\rm \pi }r^2(1 - \cos {\rm \theta }).$$
$$ A = 2{\rm \pi }r^2(1 - \cos {\rm \theta }).$$Oceans can also host complex life, as it would be highly improbable that these would be frozen to any significant depth, given modest circulation from the warmer, illuminated hemisphere. However, as the life in the oceans in this hemisphere will be constrained by ice cover; and will flank frozen land. Therefore, we assume that even with biological complexity in the oceans, such complexity will have limited impact on the sub-aerial biosphere.
Next, we consider, what areas of the illuminated landscape will be available for speciation. If we consider which life might host intelligent life, then we can exclude the oceans as aquatic organisms, irrespective of their intrinsic intellect will never be able to develop technology. Moreover, the oceans hold less oxygen; therefore, except where the oceans are intrinsically warm, metabolic rates will be unlikely to support organisms with highly complex nervous systems.
Returning to the land, variations in temperature, light intensity and humidity constrain the habitable niche for subaerial organisms (Kay et al., Reference Kay, Madden, Van Schaik and Higdon1997; Gaston, Reference Gaston2000; Šímová et al., Reference Šímová, Violle, Kraft, Storch, Svenning, Boyle, Donoghue, Jørgensen, McGill, Morueta-Holme, Piel, Peet, Regetz, Schildhauer, Spencer, Thiers, Wiser and Enquist2015; Gatti, Reference Gatti2016a, Reference Gatti2016b). This area is referred to as the niche amplitude and is defined by equation 3. Niche amplitude defines the habitable hypervolume, in terms of the planet's surface conditions (Gatti, Reference Gatti2016a, Reference Gatti2016b; Blonder, Reference Blonder2017).
 $$V_n = \sqrt {(T^2 + H^2 + {\rm NP}{\rm P}^2)}. $$
$$V_n = \sqrt {(T^2 + H^2 + {\rm NP}{\rm P}^2)}. $$Here, T is the temperature, H is the humidity and NPP is the net primary productivity of photosynthetic organisms. That is the capacity of photosynthesis to fix carbon, or gross primary productivity, minus respiratory losses (RL). These variables adequately constrain the overall biological diversity on the Earth (Gatti, Reference Gatti2016a, Reference Gatti2016b). For a synchronously rotating planet, temperature and light vary in an identical manner around the SSP, in a manner dependent on the angular separation from the SSP. This is given by equation 4:
 $$I_{\rm a} = I\cos {\rm \theta }.$$
$$I_{\rm a} = I\cos {\rm \theta }.$$We may then substitute equation 4 into equation 3, giving rise to a simple relationship between angular separation, temperature and light intensity – which will be the primary determinants of net primary productivity in photosynthetic organisms. This gives us equation 5, where niche amplitude relates simply to the physical conditions pertaining to the angular separation from the SSP:
 $$V_n = \cos {\rm \theta} \sqrt {(k_1^2 + k_2^2 + k_3^2 )}, $$
$$V_n = \cos {\rm \theta} \sqrt {(k_1^2 + k_2^2 + k_3^2 )}, $$where, in this instance, k 1–k 3 are constants relating to: (1) the optical density of the atmosphere, the abundance of greenhouse gases and the transport of heat; (2) the availability and transport of moisture; and (3) the overall efficiency of photosynthetic processes. The angle, θ, is the angular separation from the SSP as before.
While we cannot directly determine how probable it is for intelligent life to arise, these simple mathematical relationships can be used to constrain the overall biological potential of a planet, using simple astrophysically determinable quantities.
We can then constrain habitable regions, with appropriate temperature, humidity and light intensity using the final equation 6. However, we note that some organisms – particularly intelligent species or generalists – may survive in a broader variety of ecosystems, assuming equation 5 constrains the availability of nutrition.
Using a modified form of equation 2, we can apply a simple equation that delineates ‘latitudinal bands’ around the SSP, rather than around the rotation axis, as it would be on an asynchronously rotating planet. On an asynchronously rotating planet, the latitudinal band is just that – a region spanning the entire surface, bounded by two latitudinal coordinates that subtend the rotation pole. However, for a synchronously rotating planet, this ‘band’ is a region flanking the SSP, bounded by specific angular separations. Equation 2 then becomes equation 7, where θ1 and θ2 are the boundary latitudes:
 $$A = 2{\rm \pi }r^2(\sin {\rm \theta }_1 - \sin {\rm \theta }_2).$$
$$A = 2{\rm \pi }r^2(\sin {\rm \theta }_1 - \sin {\rm \theta }_2).$$Finally, we consider a realistic planet that operates plate tectonics. While plate tectonics is not the only means through which volatiles may be cycled in and out of the mantle, it is perhaps the most efficient method. Consequently, while we consider alternatives in the Discussion section, we will focus, here, on planets with a mobile lid. The principle motivation is not volatile cycling and overall habitability, rather the constraint on ‘superficial’ habitability, imposed by the physical motion of ecosystems across the planet's surface.
Using the Earth as an analogue, we find plate velocities are typically on the order of 0.5–15 cm year−1. If we consider subaerial land – continental crust – this falls to 0.5–4 cm−1 (Zahirovic et al., Reference Zahirovic, Müller, Seton and Flament2015), with the largest and oldest continents moving at the most sluggish velocities. Assuming that for a typical telluric planet, continental crust will move with similar velocities, we can now consider how quickly organisms will move into and out of regions with suitable conditions.
Results
Plate tectonics and resident time
A species will occupy a hypervolume that is defined by the environmental characteristics found in its niche (Rosenzweig, Reference Rosenzweig1968; Šímová et al., Reference Šímová, Violle, Kraft, Storch, Svenning, Boyle, Donoghue, Jørgensen, McGill, Morueta-Holme, Piel, Peet, Regetz, Schildhauer, Spencer, Thiers, Wiser and Enquist2015; Blonder, Reference Blonder2017). Therefore, there will be a defined geographical area in which this species will survive and prosper. On any planet that has plate tectonics, habitable areas will migrate across continental surface as the continents, as a whole, move with plate motion. Organisms that are suited to the highest temperatures will be restricted to the regions proximal to the SSP, while cold-adapted species will be restricted to regions in which lie nearest the terminator, or at altitude. This pattern will clearly migrate towards or away from the SSP in a manner dictated by orbital separation and greenhouse gas abundance. On planets that are intrinsically warmer, either because they lie on the inner edge of the habitable zone or have a greater abundance of greenhouse gases in their atmosphere, the habitable ’torrid’ region will lie closer to the terminator than it will for intrinsically cooler planets (Leconte et al., Reference Leconte, Forget, Charnay, Wordsworth, Selsis, Millour and Spiga2013; Wolf, Reference Wolf2017).
In a simplistic (and unrealistic) planet model with an even surface and no intruding geographical features, we can imagine that for particular species the habitable hypervolume will vary in concentric bands around the SSP delineated by equation 6 where conditions vary according to the overall parameters set in equation 5. In the simplest scenario, the terminator defines the habitable edge of this surface for all complex species and the entire illuminated hemisphere is equally habitable. However, as this is grossly untrue for life on Earth – and makes no sense in general, we need to consider the ‘latitudinal band model’, where the range of a species is constrained in a particular hypervolume with set conditions bounded by different angular separations from the SSP.
If we assume that continents move across tidally locked planets with velocities similar to those on Earth, we can calculate how long it will take a geographical region to move from one environmental region with a defined hypervolume to another. Terrestrial continents typically form the slowest moving plate components on Earth, with velocities on the order of 0.5–5 cm year−1 (Zahirovic et al., Reference Zahirovic, Müller, Seton and Flament2015). The oldest continental surfaces on Earth contain cratonic regions dating back to the Archaean. There is a generic relationship between the age, the cratonic volume and the plate velocity (Zahirovic et al., Reference Zahirovic, Müller, Seton and Flament2015). Those regions with the greatest volume of craton likely have deeply penetrating mantle roots and these slow the continental motion through the upper mantle. On a planet, with a radius equivalent to the Earth, this takes the continent from the SSP to the terminator in 200 million to 2 billion years, assuming an unlikely and unrealistic straight path.
Our understanding of terrestrial plate motion is limited in terms of longitudinal and latitudinal motion by magnetic stripes in the ocean floor (Wu and Kravchinsky, Reference Wu and Kravchinsky2014; Zahirovic et al., Reference Zahirovic, Müller, Seton and Flament2015). Prior to 200 million years ago, paleolatitude is determined for continental crust using information from the orientation of the magnetic field that is preserved in rocks of varying ages (Wu and Kravchinsky, Reference Wu and Kravchinsky2014). However, determination of paleolongitude is trickier as we assume the magnetic field varies little in this direction (Wu and Kravchinsky, Reference Wu and Kravchinsky2014). Consequently, until very recently, relative plate positions, prior to the break-up of Pangaea, have been determined indirectly using apparent polar wander paths (Wu and Kravchinsky, Reference Wu and Kravchinsky2014; Wei, Reference Wei2017).
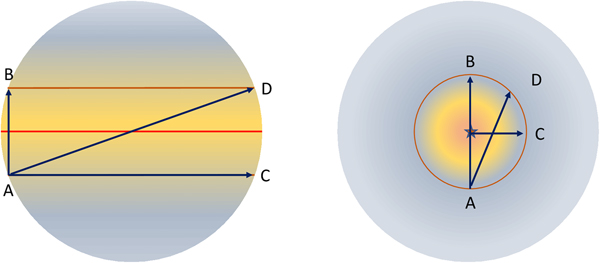
If we generalize about planetary size and assume that organisms will occupy defined concentric zones around the SSP, then we can estimate how long a species will have in its habitable niche before encountering conditions that are unfavourable for it. We will assume, for the sake of simplicity that each ecosystem occupies a band that is 10o wide (Fig. 1). The arc length that subtends this region is given by equation 7:
 $$s = {\rm \theta }r,$$
$$s = {\rm \theta }r,$$where s is the arc length, θ is the angle in radians and r is the planetary radius. The time taken to cross the terminator from the SSP is given by equation 8:
 $$t = \displaystyle{{{\rm \theta} r} \over v},$$
$$t = \displaystyle{{{\rm \theta} r} \over v},$$where t is the time in years, θ and r have the same meaning as equation 5, while v is the average plate velocity in mm year−1. θ will be π/2 radians for a full transit from SSP to terminator.
Fig. 1. The key difference between the habitability and evolutionary potential of a planet is defined by the area of its surface that is habitable for any particular species. An asynchronously rotating planet (left) has latitudinal bands with a far larger surface area than the corresponding ecological domains on a planet that is synchronously rotating with its star (right).
The time taken to cross an ecological region, such as a tropical or sub-tropical domain will be defined by its breadth, but if we assume each ecological domain is 10° wide relative to the SSP, then the time to cross each will approximate one-ninth the time required to move from the terminator to the SSP, or vice-versa (Fig. 1). This is defined by equation 8. This sets a time of approximately 20–220 million years that a species will spend in a region that has the conditions needed to support it. If one compares this to the Earth, an entire latitudinal band is conceivably habitable for any species, assuming no geographical variation in rainfall, temperature, humidity or whatever other niche quality is needed for its survival. The time it takes a continent to move from the equator to the edge of the tropics then depends on the path it takes across the band. This will be proportional to the angle, relative to the polar axis. This is illustrated in Fig. 2. This allows species to wander with their continent effectively indefinitely if they remain in a longitudinal band. Since there seems an equal probability that a plate will move in a latitudinal and longitudinal direction, the shape of the continuously habitable region around the SSP is then significant.
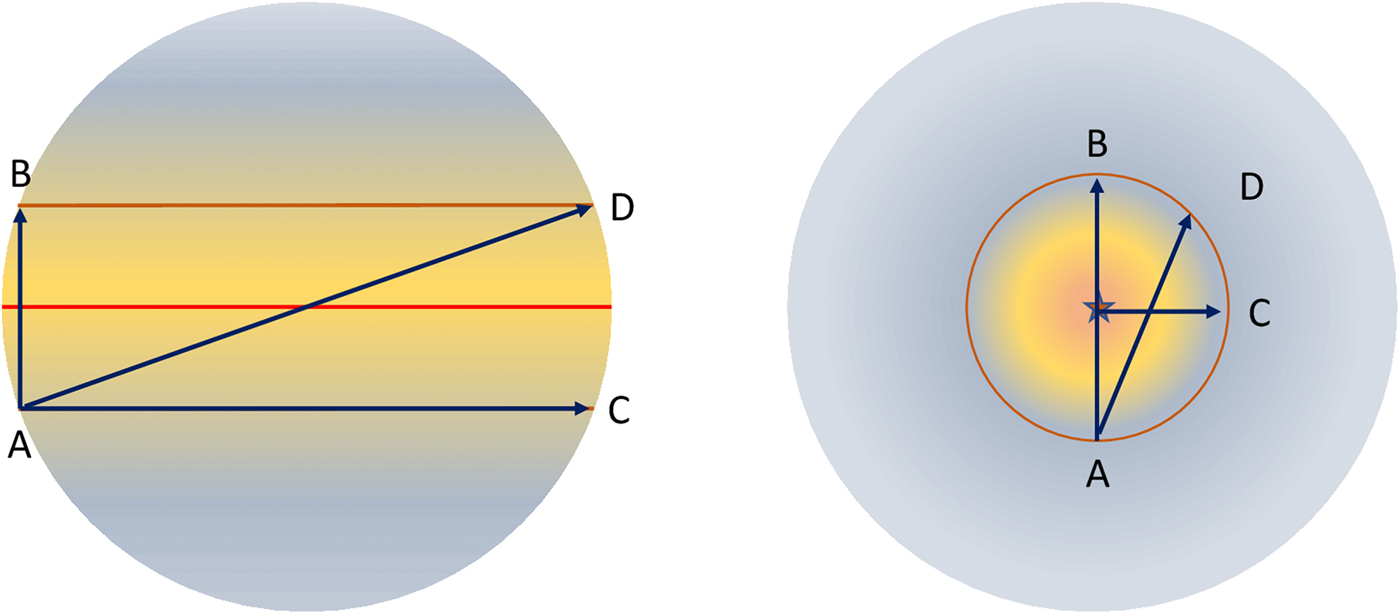
Fig. 2. The evolutionary timescale an organism (or rather a population of an organism) has is determined by the relative plate velocity. While the latitudinal timescale (the time to migrate in a north–south direction, A to B) is similar on both planets, this is untrue of motion in other planes. Movement from A to C or A to D takes considerably 1onger on the asynchronously rotating planet than on the synchronously rotating world. There is considerable impact on the niche hypervolume, as defined by light intensity, humidity and temperature. (We ignore the effects of ocean currents, which modify this even more – and of course assume that this planet is a perfect sphere.)
One might then assume that if the angular width of the band is 10° wide (the angular separation from the SSP), this would be equivalent to a latitudinal band of comparable latitude on Earth. However, it is not for two reasons. The first is prosaic. If one considers the area on a synchronous rotator that has the highest temperatures, this is a cap defined by equation 2, rather than a band defined by equation 6. Therefore, this is far smaller in extent. Thus, unless a species in this area is tolerant of a wider range of conditions than is the case for most species on Earth, the continuously habitable hypervolume will be far smaller (Gillman et al., Reference Gillman, Wright, Cusens, McBride, Malhi and Whittaker2015).
Similarly, but less obviously, each climatic region on Earth is duplicated around the equatorial band. A synchronously rotating planet has one equivalent band, whose width depends on the abundance of greenhouse gases and incident stellar flux. Therefore, by this criterion, the area is also smaller. Moreover, if the most complex species occupy the region closest to the terminator, they will be at a far higher risk of extinction by virtue of the fact plate tectonic motion may carry them across the terminator into cold darkness.
In essence, species that evolve on synchronously rotating planets will spend proportionately less time in a region that is habitable for them, compared with corresponding species evolving on asynchronously rotating planets. Consequently, sub-aerial species are far more likely to become extinct due to climatological constraints on tidally locked planets than on non-synchronously rotating worlds. Moreover, remember that the terminator forms an absolute limit for photosynthetic life and anything that depends on it. On the Earth, photosynthesis continues unabated to the poles, with only the frozen core of Antarctica being inhospitable to it. The presence of photosynthetic life in Antarctica's dry valleys and in the peninsula illustrates the robust nature of the process, but emphasizes the difference with synchronously rotating planets (Fell et al., Reference Fell, Scorzetti, Connell and Craig2006).
The impact of atmospheric circulation models
The impact of synchronous rotation on habitability is accentuated by global atmospheric circulation patterns. On synchronous but slow-rotating planets (those with orbital periods exceeding 20 days), airflow is broadly radial towards the SSP (Haqq-Misra et al., Reference Haqq-Misra, Wolf, Joshi, Zhang and Kopparapu2017; Turbet et al., Reference Turbet, Bolmont, Leconte, Forget, Selsis, Tobie, Caldas, Naar and Gillon2017; Wolf, Reference Wolf2017; Wolf et al., Reference Wolf, Shields, Kopparapu, Haqq-Misra and Toon2017). This brings cold air over the terminator at the surface towards the SSP. Such air, as well as being cold is likely to hold relatively little moisture and have a low relative humidity. Moreover, circulation models posit the presence of a strong, overlying temperature inversion aloft, that will inhibit convection, distant to the SSP. Therefore, continental surfaces located distally to the SSP (and proximal to the terminator) are likely to have a low potential biodiversity as defined by equation 5. Here, equation 5 substitutes for other measures of species diversity, such as the Shannon Index (H′) or Simpson Index (D). This is permissible as terrestrial biological diversity follows the niche amplitude as defined by equations 3 and 5 (Gillman et al., Reference Gillman, Wright, Cusens, McBride, Malhi and Whittaker2015). Here, the limiting factor is humidity, with terminator–proximal regions dry, and consequently having a low niche amplitude (Gatti, Reference Gatti2016a).
Synchronously rotating planets with shorter orbital periods have more focused atmospheric flow along zonal bands. While this may expand the regions in which niche amplitude is higher (Stevenson and Large, Reference Stevenson and Large2017; Stevenson, Reference Stevenson2018a, Reference Stevenson2018b), the continental surface over which complex life may evolve remains considerably restricted compared with the Earth. However, speciation in the oceans should, in principle, resemble that of the Earth. This is because the number of available niches is likely to be similar between synchronously and asynchronously rotating planets. The presence of a perpetually dark hemisphere is offset by the presence of its opposite. While the total number of geographical niches may be half that of the Earth, there will still be significant numbers of niches in the dark hemisphere, with depth being the primary consideration. Clearly, the number of epipelagic (surface) and mesopelagic (twilight) zones will be restricted to the illuminated hemisphere. However, these constitute a relatively minor volume of the oceans and hence have a lesser impact on the number of available niches.
Additional limiting factors on niche availability
Moreover, the manner in which oceans circulate may further limit (but possibly enhance) habitable regions by altering precipitation and temperature profiles (Cullum et al., Reference Cullum, Stevens and Joshi2014; de Lavergne et al., Reference de Lavergne, Madec, Roquet, Holmes and McDougall2017; Stevenson, Reference Stevenson2018a).
Temporal niches, whereby organisms are active only at certain times of the day, will obviously be absent on synchronously rotating planets: there is no temporal variation. The only exception will be where the central star is eclipsed by any sister planets giving rise to short periods of darkness or substantive periodic dimming. Thus, the number of biological niches is lower still on synchronously rotating worlds, compared with asynchronously rotating planets, like the Earth.
An important consideration for the evolution of sub-aerial animal life will be the abundance and variety of coastal niches, into which aquatic life can evolve. If these are limited, for example, by limited land cover in the illuminated hemisphere, evolution of land-based species may be restricted. Thus, plate configurations and velocities will be an important determinant of speciation of life on land.
Plate tectonics, thick lids and the evolution of complexity
This simple analysis suggests that synchronously rotating planets will be relatively poor abodes for the evolution of complex, multicellular life and hence intelligent species. Remember that this conclusion is dependent on such intelligent species evolving on land and on planets that operate plate tectonics. The evolution of biological complexity in the oceans is not affected.
The get-out-clause is provided by the mass of the planet. Modelling suggests that plate tectonics will operate for periods defined by planetary mass and the mode in which the mantle convects (O'Neill et al., Reference O'Neill, Jellinek and Lenardic2005, Reference O'Neill, Lenardic, Weller, Moresi, Quenette and Zhang2016; Kite et al., Reference Kite, Manga and Gaidos2008; Korenaga, Reference Korenaga2012; Tackley et al., Reference Tackley, Ammann, Brodholt, Dobson and Valencia2012; Bercovici et al., Reference Bercovici, Tackley and Ricard2015). Planets with very low mass or super-terrans with masses in excess of three Earth masses are likely to cease plate tectonics at early periods, either because they become too cool at early epochs, or because they form a thick, stable granitic lid which resists subduction. Some planets may skip plate tectonics altogether if they produce a thick, stagnant, lid at early times. While these planets may have other problems relating to the function of the carbonate–silicate cycle (Edson et al., Reference Edson, Lee, Bannon, Kasting and Pollard2011, Reference Edson, Kasting, Pollard, Lee and Bannon2012), such worlds will have a stable surface on which complexity can evolve, unaffected by plate motion.
Finally, the duration in which terrestrial planets may retain surface oceans also varies with planetary mass and the mode of mantle convection (Korenaga, Reference Korenaga2011; Schaefer and Sasselov, Reference Schaefer and Sasselov2015). Thus, plate tectonics is a double-edged sword – an important (but perhaps not essential) prerequisite for the maintenance of a carbonate–silicate cycle and oceans, but detrimental to the evolution of biological complexity on synchronously rotating planets.
The ‘intellectual’ necessity of continental crust
While the Earth developed biological complexity initially in its oceans, it was the land on which mammals and homeothermy evolved (Kurbel, Reference Kurbel2014). Homeothermy is almost certainly going to evolve on land, where temperatures vary diurnally and seasonally. Oceans are on average cooler, have lower levels of variation in conditions and, critically, have lower concentrations of oxygen than continental surfaces. Limited oxygen reduces the ability of organisms to operate the kind of active metabolisms required for homeothermy. In turn, ‘warm-bloodedness’ allows organisms to continually operate the kinds of active metabolisms needed for intelligence. While some poikilothermic species may retain active metabolism if the environment is continually warm (as it may be in the oceans or in close proximity to the SSP), plate motion will require mass migration of species to keep pace with continental movement. As continental area is not limitless, this may not be possible unless a shallow, thick, stagnant lid is present.
In this regard, one might suspect that the persistence of temperatures over millions of years at any one location, relative to the SSP, will preclude the evolution of homeothermy on such worlds. While plate tectonics may limit the time that is available for the evolution of biological complexity at any one location, an absence of it may remove the environmental pressure required for the evolution of homeothermy and consequently intelligence. This may add a further twist that explains why intelligent aliens never drop by. Without an evolutionary selective pressure, homeotherms never arise on such worlds and the kind of high-metabolism organism never emerges that looks up at the sky in awe. A varying climate becomes a necessity, thus eliminating 75% of the possible stars in the universe as hosts for indigenous, intelligent life.
More fundamentally, if any complex species is non-technological, sooner or later continental motion will take them either to less habitable regions of the surface – or across the terminator where they will cease to exist (Stevenson, Reference Stevenson2017, Reference Stevenson2018b; Stevenson and Large, Reference Stevenson and Large2017). Operation of plate tectonics on tidally locked planets then leads to the geographical restriction of species through its dispersal and likely extinction.
Discussion
In this paper, we show that there is a simple and rather prosaic reason for ‘the great silence’. The majority of habitable planets – as determined by surface temperatures and the presence of liquid water – are likely to orbit red dwarf stars (Heath et al., Reference Heath, Doyle, Joshi and Haberle1999; Irwin et al., Reference Irwin, Charbonneau, Nutzman and Falco2008; Brugger et al., Reference Brugger, Mousis, Deleuil and Lunine2016). If we assume that atmospheric erosion is insufficient to sterilize all of the conceivably habitable planets (e.g. Wheatley et al., Reference Wheatley, Louden, Bourrier, Ehrenreich and Gillon2016; Turbet et al., Reference Turbet, Bolmont, Leconte, Forget, Selsis, Tobie, Caldas, Naar and Gillon2017), we are still left with a more prosaic reason for the ‘great silence’: plate tectonics and limited biological hypervolumes.
Plate tectonics may be essential for the maintenance of an active carbonate–silicate cycle that, in turn, is needed for the long-term habitability of all terrestrial planets (Edson et al., Reference Edson, Lee, Bannon, Kasting and Pollard2011, Reference Edson, Kasting, Pollard, Lee and Bannon2012). However, on tidally locked planets, the effective surface over which species may evolve is constrained by the presence of the terminator. If one considers individual species, the time each has in a particular climatic region is tightly constrained by synchronous rotation, with proportionately less time available in which to proliferate and hence evolve new species through natural selection.
Moreover, the absence of a diurnal cycle limits the information complexity of the environment and the number of temporal niches (Stevenson and Large, Reference Stevenson and Large2017).
While, we cannot be certain that the rather prosaic reason presented in this paper may give the greatest contribution to explaining the great silence. Synchronous rotation, when coupled with plate tectonic motion, may be sufficient to restrict the development of intelligent species to a minority of asynchronously rotating planets (with respect to their host star).
The exception will be those planets that are more massive or older than the Earth and have stagnant-lid surfaces (or have episodic tectonics). In some instances, one can envisage a limited carbonate–silicate cycle if they remain volcanically active (Fig. 3). Such worlds may be perfectly habitable and suitable for the evolution of biological complexity even where the planet is a synchronous rotator. However, even here, the lack of temporal variation (a lack of a diurnal cycle) will grossly restrict the number of available niches, with one niche per unit area, over geological periods. Asynchronous rotators, by contrast, will have temporal niches – ones that vary over the course of the day. This allows different species to occupy different temporal spaces that are geographically delineated.

Fig. 3. Vertical tectonics with a stagnant lid. Mantle plumes, at various stages in their evolution cause bulges, volcanism, while thick piles of volcanic material or mantle downwellings cause collapse with burial of carbonate rocks (green). A limited carbonate–silicate cycle is, therefore, possible irrespective of the presence of plate tectonics. This figure is reproduced, in modified form, from Stevenson (Reference Stevenson2018b).
One concludes that, while synchronously rotating planets make ideal habitats for the evolution of life in general, they are poor hosts for the evolution of complexity on land, where plate tectonics is active. The ‘great silence’ may be down to the simple truth that the majority of habitable worlds orbit red dwarfs and this limits the effective habitable surface and the number of available niches for the evolution of biological complexity. Finally, and perhaps fatally, the invariant climate in at each geographical location, relative to the SSP, removes selective pressure for the evolution of homeothermy. An active, inquisitive mind requires an active, temporally invariant metabolism just as much as it needs an evolving but habitable landscape. We suggest that a true measure of the biological complexity of planets will require a thorough mathematical analysis of the number of available niches over varying time intervals. While this is beyond the scope of this publication, such an analysis will undoubtedly refine and delineate those worlds that will host complexity and which proportion of these worlds will be able to host the evolution of intelligent life. We intend to tackle this in a future publication.
One hopes that these conclusions underscore the true complexity of biological evolution when it comes to determining which planets are most likely to host the development of biological complexity. In that sense, one hopes to see ‘astrobiology’ evolve into ‘astrogeography’ and ‘astroecology’ in the coming years. This paper represents a first step in this evolving journey.
Acknowledgements
I would also like to acknowledge the help and assistance of Alfie Gemmell and Christopher Loftus in reviewing some of the content of this manuscript. The also would like to thank the anonymous reviewer who suggested a number of important revisions that have made this article more lucid and pertinent.