Introduction
Ediacaran tubular fossils are the earliest skeletal metazoans, thus being important for understanding early metazoan evolution. Studies in the last few decades have made significant progresses in their phylogenetic affinity (Grant, Reference Grant1990; Hua et al., Reference Hua, Chen, Yuan, Zhang and Xiao2005; Vinn and Zatoń, Reference Vinn and Zatoń2012; Schiffbauer et al., Reference Schiffbauer, Selly, Jacquet, Merz, Nelson, Strange, Cai and Smith2020; Selly et al., Reference Selly, Schiffbauer, Jacquet, Smith and Nelson2020; Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b), paleoecology (Bengtson and Yue, Reference Bengtson and Yue1992; Cai et al., Reference Cai, Hua and Zhang2013; Penny et al., Reference Penny, Wood, Curtis, Bowyer, Tostevin and Hoffman2014; Cortijo et al., Reference Cortijo, Cai, Hua, Schiffbauer and Xiao2015a; Becker-Kerber et al., Reference Becker-Kerber, Pacheco, Rudnitzki, Galante, Rodrigues and de Moraes Leme2017; Wood et al., Reference Wood, Curtis, Penny, Zhuravlev, Curtis-Walcott, Iipinge and Bowyer2017a; Mehra and Maloof, Reference Mehra and Maloof2018; Shore and Wood, Reference Shore and Wood2021), biomineralization (Grant, Reference Grant1990; Chen et al., Reference Chen, Bengtson, Zhou, Hua and Yue2008; Wood et al., Reference Wood, Zhuravlev, Sukhov, Zhu and Zhao2017c; Pruss et al., Reference Pruss, Blättler, Macdonald and Higgins2018; Wood, Reference Wood2018; Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b), taphonomy (Grant, Reference Grant1990; Cai et al., Reference Cai, Hua, Xiao, Schiffbauer and Li2010; Pacheco et al., Reference Pacheco, Leme and Machado2011; Becker-Kerber et al., Reference Becker-Kerber, da Silva, Borges Amorim, Liza Alves Forancelli Pacheco and de Moraes Leme2019; Walde et al., Reference Walde, Weber, Erdtmann and Steiner2019; Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b), paleobiogeography (Warren et al., Reference Warren, Quaglio, Riccomini, Simões, Poiré, Stríkis, Anelli and Strikis2014, 2017), and biostratigraphy (Gaucher and Germs, Reference Gaucher, Germs, Gaucher, Sial, Halverson and Frimmel2009; Yang et al., Reference Yang, Steiner, Zhu, Li, Liu and Liu2016; Zhu et al., Reference Zhu, Zhuravlev, Wood, Zhao and Sukhov2017; Cai et al., Reference Cai, Xiao, Li and Hua2019).
Although these organisms are considered useful markers for identifying the latest Ediacaran (Xiao et al., Reference Xiao, Narbonne, Zhou, Laflamme, Grazhdankin, Moczydlowska-Vidal and Cui2016) with proposed biozonations (Gaucher and Germs, Reference Gaucher, Germs, Gaucher, Sial, Halverson and Frimmel2009; Adorno et al., Reference Adorno2017; Zhu et al., Reference Zhu, Zhuravlev, Wood, Zhao and Sukhov2017), there is no consensus on their utility for global correlation. One of the main obstacles is their taxonomic confusion. Thus, the goal of this study is to provide a systematic revision of key Ediacaran tubular taxa, as well as a comprehensive review. To do this, we return to the type materials for the Ediacaran tubular taxa, Cloudina, Sinotubulites, and Conotubus, discussing their systematic treatments following the regulation of International Code of Zoological Nomenclature (ICZN, 1999).
Materials and methods
More than 200 tubular specimens of Cloudina and Sinotubulites from Namibia, US, and Paraguay were ground and polished to produce uncovered thin-sections. Both longitudinal and transversal sections were obtained, observed with binocular microscopes, and documented with digital cameras. Published and collected specimens of Cloudina were measured with the software of ImageJ 1.48V. Statistical data were subsequently obtained with Origin Pro. 8.6.
Repositories and institutional abbreviations
Types, figures, and other specimens examined in this study are deposited in the following institutions: Institute of Geology, Chinese Academy of Geological Sciences (IGCAGS), Beijing, China; Geological Museum of China (GMC), Beijing, China; Iziko South African Museum (IZIKO), Cape Town, South Africa; Earth Sciences Museum—Brazilian Geological Survey, Rio de Janeiro, Brazil; Northwest University (NU), Xi'an, China.
Systematic paleontology
Family Cloudinidae Hahn and Pflug, Reference Hahn and Pflug1985
Genus Cloudina Germs, Reference Germs1972
Type species
Cloudina hartmannae Germs, Reference Germs1972, from the Schwarzkalk Limestone Member of Kuibis Formation (=Omkyk Member of the Zaris Formation, Kuibis Subgroup), Driedoornvlakte Farm near Schlip, Namibia, by original designation.
Other species
Cloudina carinata Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2010, and Cloudina xuanjiangpingensis Cai et al., Reference Cai, Cortijo, Schiffbauer and Hua2017.
Original diagnosis (Germs, Reference Germs1972)
The calcareous tubes are sinuous, composed of stacked cones so arranged as usually to give the appearance to the composite structure of having two walls. The outer wall bears annular ridges and depressions. The inner wall is smoother and in some specimens shows very fine annular ridges. The inner tubes are ordinarily not centrally located in transverse section. The apex has nearly always been found open, while the distal end has always been found open. Two species are recognized, C. hartmannae and C. riemkeae, as described below.
Emended diagnosis
Sinuous or straight tubular microfossils with regularly arranged collar-structures. Collars usually flare away at the upper part of the tubes with annulations, longitudinal crests, or weak ridges occasionally observed on their surfaces. The tubes have a smooth inner wall and a circular cross section. Closed proximal ends and dichotomous branchings are occasionally observed. The tube wall is composed of multi-layered laminae.
Cloudina hartmannae Germs, Reference Germs1972
Figures 1.1, 1.2, 3–5, 7.1, 8.1
- Reference Germs1972
Cloudina hartmannae Germs, p. 753, text-fig. 3C, pl. 1, figs. 1–7.
- Reference Germs1972
Cloudina riemkeae Germs, p. 755, text-fig. 3D, pl. 1, figs. 8–13.
- Reference Glaessner1976
Cloudina hartmanae; Glaessner, p. 266, text-fig. 3, pl. 1, fig. 2, pl. 2, fig. 5.
- Reference Mount, Gevirtzman and Signer1983
Coleolella sp.; Mount et al., p. 225, figs. 3C, D.
- ?Reference Signor, McMenamin, Gevirtzman and Mount1983
Coleolella sp.; Signor et al., p. 416, fig. 3b.
- ?Reference Signor, McMenamin, Gevirtzman and Mount1983
Multiple–walled tubular fossil; Signor et al., p. 416, fig. 3e.
- ?Reference Hahn and Pflug1985
Cloudina hartmannae; Hahn and Pflug, p. 422, pl. 2, fig. 5.
- [non] Reference Hahn and Pflug1985
Cloudina waldei Hahn and Pflug, p. 423, text-fig. 7, pl. 1, figs. 1, 2, pl. 2, figs. 3, 4.
- ?Reference Zaine and Fairchild1987
Cloudina lucianoi (Beurlen and Sommer, Reference Beurlen and Sommer1957); Zaine and Fairchild, p. 804, figs. 1–7.
- Reference Signor, Mount and Onken1987
Nevadatubulus dunfeei Signor, Mount, and Onken, p. 429, figs. 3, 4.1–4.14.
- Reference Conway Morris, Mattes and Chen1990
Cloudina cf. hartmannae; Conway Morris et al., p. 252, fig. 4.
- Reference Conway Morris, Mattes and Chen1990
Cloudinid-like tube; Conway Morris et al., p. 254, fig. 5.
- Reference Grant1990
Cloudina hartmannae; Grant, p. 261, figs. 4, 6A, 8D.
- Reference Grant1990
Cloudina riemkeae; Grant, p. 261, figs. 5, 7A, B, 8C.
- Reference Grant1990
Cloudina sp.; Grant, p. 261, figs. 7C, D, 10B.
- ?[non] Reference Grant1990
Cloudina sp.; Grant, p. 275, fig. 8B.
- Reference Bengtson and Yue1992
Cloudina hartmannae; Bengtson and Yue, p. 368, figs. 2, 3.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Cloudina sp.; Li et al., p. 94, text-figs. 3–6, 7:7, 8, pl. 8, figs. 4, 16, 17.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Cloudina sinensis Zhang, Li, and Dong in Li et al., p. 93, text-figs. 3–6, 7:1–6, 10, pl. 8, figs. 1–3, 5, 7–12, 14, 15, 18.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Cloudina lijiagouensis Zhang, Li, and Dong in Li et al., p. 94, text-fig. 3–6, 7:9, pl. 8, fig. 6.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus varius Zhang, Li, and Dong in Li et al., p. 92, text-fig. 3–7, 8:9, pl. 9, figs. 3, 4, 6, 9.
- Reference Zhang and Hua2000
Cloudina sinensis; Zhang and Hua, p. 326, text-fig. 1–6, pl. 1, figs. 2, 17.
- Reference Zhang and Hua2000
Cloudina lijiagouensis; Zhang and Hua, p. 332, pl. 1, fig. 3.
- Reference Zhang and Hua2000
Cloudina sp.; Zhang and Hua, p. 326, text-figs. 2, 7, 8:9, pl. 1, figs. 1, 6, 9, 14, 18.
- Reference Hua, Zhang, Zhang and Wang2000b
Cloudina sp.; Hua et al., p. 513, pl. 2, fig. 3.
- Reference Hua, Zhang, Zhang and Wang2000b
Cloudina lijiagouensis; Hua et al., p. 513, pl. 2, fig. 4.
- Reference Brain2001
Cloudina; Brain, p. 478, figs. 1, 2.
- Reference Chen and Sun2001
Cloudina sinensis; Chen and Sun, p. 195, pl. 1, figs. 1–13, pl. 2, figs. 1–8, pl. 3, figs. 1–5.
- ?Reference Hofmann and Mountjoy2001
Cloudina; Hofmann and Mountjoy, p. 1093, figs. 2G, H, J.
- ?[non] Reference Hofmann and Mountjoy2001
Cloudina; Hofmann and Mountjoy, p. 1093, fig. 2I.
- Reference Hua, Pratt and Zhang2003a
Cloudina hartmannae; Hua et al., p. 456, figs. 2D–H.
- Reference Hua, Pratt and Zhang2003a
Cloudina hartmannae; Hua et al., p. 457, fig. 3.
- Reference Hua, Zhang and Chen2003b
Morphotype II; Hua et al., p. 197, pl. 2, fig. 1.
- Reference Gaucher, Boggiani, Sprechmann, Sial and Fairchild2003
Cloudina lucianoi; Gaucher et al., p. 261, figs. 10A, B, D–K.
- Reference Hua, Chen, Yuan, Zhang and Xiao2005
Cloudina hartmannae; Hua et al., p. 278, figs. 1A–P.
- Reference Hua, Chen and Yuan2007
Cloudina hartmannae; Hua et al., p. 269, figs. 5:4, 5:6, 6:3–5.
- Reference Chen, Bengtson, Zhou, Hua and Yue2008
Cloudina; Chen et al., p. 39, fig. 2D.
- Reference Cai, Hua, Xiao, Schiffbauer and Li2010
Cloudina hartmannae; Cai et al., p. 492, fig. 4M.
- Reference Cai, Schiffbauer, Hua and Xiao2011
Cloudina lucianoi; Warren et al., p. 384, figs. 2D, 3A–E, 4A–G.
- Reference Warren, Pacheco, Fairchild, Simões, Riccomini, Boggiani and Cáceres2012
Cloudina sp.; Warren et al., p. 692, fig. 2A.
- ?Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov2012
Cloudina ex gr. C. riemkeae; Zhuravlev et al., p. 209, figs. 3A, C–E
- ?Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov2012
Cloudina ex gr. C. hartmannae; Zhuravlev et al., p. 209, figs. 3B, F–H.
- Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov2012
Cloudina hartmannae; Zhuravlev et al., p. 215 figs. 6C, D.
- Reference Warren, Quaglio, Riccomini, Simões, Poiré, Stríkis, Anelli and Strikis2014
Cloudina; Warren et al., p. 393, figs. 2A–G.
- Reference Cortijo, Cai, Hua, Schiffbauer and Xiao2015a
Cloudina; Cortijo et al., p. 421, figs. 2, 3.
- Reference Cortijo, Martí Mus, Jensen and Palacios2015b
Cloudina hartmanae; Cortijo et al., p. 189, figs. 3A–D, F, I–L, 4, 5.
- ?Reference Cortijo, Martí Mus, Jensen and Palacios2015
Cloudina lucianoi; Walde et al., p. 219, figs. 4F, 5A–D, 6 (part).
- [non] Reference Walde, do Carmo, Guimarães, Vieira, Erdtmann, Sanchez, Adorno and Tobias2015
Cloudina waldei; Walde et al., p. 219, fig. 4E.
- Reference Yang, Steiner, Zhu, Li, Liu and Liu2016
Cloudina hartmannae; Yang et al., p. 210, figs. 4A–D, G, H, 5D.
- Reference Chen, Zhou, Zhang, Wei and Zhang2016
Cloudina hartmannae; Chen et al., p. 98, fig. 3C, D.
- Reference Xiao, Narbonne, Zhou, Laflamme, Grazhdankin, Moczydlowska-Vidal and Cui2016
Cloudina riemkeae; Xiao et al., p. 544, fig. 4G, H.
- Reference Cai, Cortijo, Schiffbauer and Hua2017
Cloudina hartmannae; Cai et al., p. 150, fig. 4.
- Reference Cai, Cortijo, Schiffbauer and Hua2017
Cloudina ningqiangensis Cai et al., p. 153, fig 6.
- Reference Becker-Kerber, Pacheco, Rudnitzki, Galante, Rodrigues and de Moraes Leme2017
Cloudina hartmannae; Becker-Kerber et al., p. 4, figs. 2, 3A, C, D, F–I.
- Reference Adorno2017
Cloudina hartmanae; Adorno et al., p. 30, fig. 10A, C.
- [non] Reference Adorno2017
Cloudina hartmanae; Adorno et al., p. 30, fig. 10B.
- Reference Adorno2017
Cloudina hartmannae; Adorno et al., p. 30, fig. 11A.
- Reference Adorno2017
Cloudina riemkeae; Adorno et al., p. 30, fig. 11C.
- ?Reference Adorno2017
Cloudina lucianoi; Adorno et al., p. 30, fig. 11B.
- ?Reference Adorno2017
Cloudina lucianoi; Adorno et al., p. 29, figs. 9, 10D, E, 12K.
- ?[non] Reference Adorno2017
Cloudina lucianoi; Adorno et al., p. 32, fig. 12A–J, L–S.
- Reference Min, Hong, Yaoping and Bo2019a
Cloudina; Min et al., p. 19, figs. 1A–D, 2.
- Reference Min, Hong, Yaoping and Bo2019a
Multiconotubus; Min et al., p. 19, fig. 1E–K.
- Reference Becker-Kerber, da Silva, Borges Amorim, Liza Alves Forancelli Pacheco and de Moraes Leme2019
Cloudina; Becker-Kerber et al., p. 3, 8, figs. 3, 8.
- ?[non] Reference Liang, Cai, Nolan and Xiao2020
Cloudina ningqiangensis; Liang et al., p. 4, fig. 3.
- Reference Liang, Cai, Nolan and Xiao2020
Cloudina hartmannae; Liang et al., p. 5, fig. 4.
- ?Reference Liang, Cai, Nolan and Xiao2020
Cloudina cf. carinata; Liang et al., p. 6, fig. 5c.
- Reference Shore, Wood, Curtis and Bowyer2020
Cloudina hartmannae; Shore et al., p. 878, figs. 1C, 3I, J.
- Reference Yang, Shang, Steiner and Liu2020a
Cloudina hartmannae; Yang et al., p. 450, figs. 2A–C.
- Reference Yang, Shang, Steiner and Liu2020b
Cloudina hartmannae; Yang et al., p. 4, figs. 2D, E, 3A, B, 5A.
Holotype
S.A.M.K. 1071 (Fig. 1.1), Iziko South African Museum, Cape Town, South Africa.

Figure 1. Originally published materials of Cloudina (1, 2) from the Omkyk Member (formerly named Schwarzkalk Limestone Member) of the Zaris Formation at Driedoornvlakte Farm near Schlip, Namibia (Germs, Reference Germs1972; photo courtesy of the Iziko Museum of South Africa from specimens housed at this institution) and Aulophycus (3–9) reported from the Tamengo Formation, Corumbá Group, Ladário Town, Mato Grosso do Sul State, Brazil (Beurlen and Sommer, Reference Beurlen and Sommer1957; photo courtesy of the Earth Sciences Museum—Brazilian Geological Survey, from specimens housed at this institution). (1) Holotype of Cloudina hartmannae, S.A.M.K. 1071. (2) Holotype of Cloudina riemkeae (=Cloudina hartmannae), S.A.M.K. 1072. (3) Holotype of Aulophycus lucianoi, D.G.M. 1149. (4) Close-up photograph of (3) showing cross-sections with single wall and one longitudinal section with a single wall. (5) Close-up photograph of (3) showing multi-walls and angular folds resembling Sinotubulites (red arrow); the photograph also shows cross sections with various diameters. (6) Close-up photograph of (3) showing cross-section specimens with a single wall. (7) Close-up photograph of (3) showing longitudinal sections with a single wall and specimens with various diameters. (8) Specimens from paratype D.G.M. 1156 shows specimens with various diameter and single wall, some of which resemble Namacalathus (e.g., white arrow); one specimen with multi-walls and angular folds resembles Sinotubulites (red arrow). (9) Specimens from paratype D.G.M. 1156 show cross sections with a single wall and various diameters. Scale bars: (1) 5 mm; (2) 1 mm; (3) 2 cm; (4–7) 300 μm; (8, 9) 500 μm.
Diagnosis
Organisms forming tubular exoskeletons with collar structures. Elongate collars are close to the tubular body near the base to lower part and flare away at the upper part of the tubes. The tube wall is composed of multi-layered laminae, smooth inner wall, and a circular cross section. Closed proximal end and dichotomous branching are occasionally observed.
Occurrence
Omkyk (Germs, Reference Germs1972; Grant, Reference Grant1990; Brain, Reference Brain2001), Nasep (Germs, Reference Germs1995), Mara? (Germs, Reference Germs1983), Mooifontein (Germs, Reference Germs1983; Grant, Reference Grant1990; Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b; Figs. 3, 4), Kliphoek? (Grant, Reference Grant1990), Huns (Grant, Reference Grant1990; Germs, Reference Germs1995), and Spitskop (Germs, Reference Germs1995) members of the Nama Group, South Namibia; Tamengo Formation, Corumbá Group, Mato Grosso do Sul State, Brazil (Beurlen and Sommer, Reference Beurlen and Sommer1957; Adorno et al., Reference Adorno2017; Becker-Kerber et al., Reference Becker-Kerber, Pacheco, Rudnitzki, Galante, Rodrigues and de Moraes Leme2017); Sete Lagoas Formation, Bambuí Group, Januária area, central Brazil (Warren et al., Reference Warren, Quaglio, Riccomini, Simões, Poiré, Stríkis, Anelli and Strikis2014); Tagatiya Guazu Formation, Itapucumi Group, northeastern Paraguay (Warren et al., Reference Warren, Fairchild, Gaucher, Boggiani, Poiré, Anelli and Inchausti2011, Reference Warren, Quaglio, Simões, Gaucher, Riccomini, Poiré, Freitas, Boggiani and Sial2017; this study); Shibantan (Chen and Wang, Reference Chen and Wang1977; Fig. 7) and Baimatuo (Liang et al., Reference Liang, Cai, Nolan and Xiao2020) members of Dengying Formation, Yichang City of Hubei Province, China; Dengying Formation, Shennongjia area of Hubei Province, South China, (Steiner et al., Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Yang et al., Reference Yang, Shang, Steiner and Liu2020a); Dengying Formation (Conway Morris et al., Reference Conway Morris, Mattes and Chen1990; Hua et al., Reference Hua, Chen, Yuan, Zhang and Xiao2005; Cai et al., Reference Cai, Cortijo, Schiffbauer and Hua2017), Ningqiang County of Shaanxi Province, South China; Kuanchuanpu Formation, Ningqiang County of Shaanxi Province, South China (Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b); Deep Spring Formation (Mount et al., Reference Mount, Gevirtzman and Signer1983; reported as Coleolella sp.; Signor et al., Reference Signor, Mount and Onken1987; reported as Nevadatubulus; this study), western USA; Villarta Limestone of Ibor Group, Villarta de los Montes Village, Bada-joz Province, Spain (Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2015b); Byng Formation of Miette Group, Salient Mountain area, British Columbia, Canada (Hofmann and Mountjoy, Reference Hofmann and Mountjoy2001); Ara Formation of Huqf Group, Birba Area, Oman (Conway Morris et al., Reference Conway Morris, Mattes and Chen1990); Ust’-Yudoma Formation, Uchur-Maya region, Southeast Siberia (Zhuravlev et al., Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov2012; Zhu et al., Reference Zhu, Zhuravlev, Wood, Zhao and Sukhov2017); Tarzhul’ Formation in Kuznetsk Alatau (Terleev et al., Reference Terleev, Postnikov, Tokarev, Sosnovskaya and Bagmet2011) and Raiga Formation in Tomsk Region (Kontorovich et al., Reference Kontorovich, Varlamov, Grazhdankin, Karlova and Klets2008), West Siberia, Russia.

Figure 3. Cloudina hartmannae from the Mooifontein Member of Nama Group in Aar Farm, Namibia (1, 2, 5–7) and Tagatiya Guazu Formation, Itapucumi Group, northeastern Paraguay (3, 4, 8); note the evident morphological variations between distinct specimens, some of which (5) resemble Multiconotubus chinensis Cai et al., Reference Cai, Cortijo, Schiffbauer and Hua2017. (1) A0803. (2) A1201. (3) PGC010202. (4) PGC010501. (5, 7) NFA210103–2. (6) A01–02. (8) PGC010602. Scale bars: (5) 200 μm; others, 500 μm. Specimens are deposited at the Institute of Geology, Chinese Academy of Geological Sciences (CAGS).

Figure 4. Transversal views of Cloudina hartmannae Germs, Reference Germs1972 from Mooifontein Member of Nama Group, Aar Farm, Namibia (1), northeastern Paraguay (2) and Deep Spring Formation of Mount Dunfee in Nevada, USA (3). (1) NFA210202–2. (2) PGC010603. (3) ELG370501. Scale bars: (1, 2) are 200 μm; (3), 500 μm. Specimens are deposited at the Institute of Geology, CAGS.
Description
Straight or sinuous tubular constructions exist with both closely or widely expanding collared structures. Fossil length varies from hundreds of micrometers to several centimeters, with a diameter range of hundreds of micrometers to ~5 mm. Cross sections are usually circular, but sometimes oval due to compression. Wall thickness is between 10–30 μm. Multi-layered laminae of the tube wall range from 1–5 μm in thickness and preserve organic residues.
Measurements
The width and spacing of collar structures of previously described species of Cloudina were measured to provide a statistic assessment on the size variation of the genus and its different species (Fig. 2). The data are based on the type material, additional published material, as well as our collected specimens (Supplemental data). Only specimens that were preserved in a state of preservation that secured an exact size measurement were measured, which resulted in 190 data sets that were obtained based on literature and our own material. It is emphasized that the size measurements can partly be affected by the fossil preservation (e.g., recrystallization and additional cement growth on tubes) and may have been biased by selective sampling and preparation methods (e.g., specimens extracted by acid or presented by thin-section). Despite this, general size trends indicated by the data are considered to be useful as a support of the systematic work. The measurements show that collar width and spacing have a wide range of variation (60 to ~4600 μm and 36 to ~3750 μm, respectively; Fig. 2.1) while they are relatively well correlated in an approximately linear trend. Cloudina carinata is generally larger than most individuals of Cloudina hartmannae and other species. However, some of the specimens of Cloudina hartmannae, including its holotype, are larger than most of the other collected specimens. The majority of specimens of Cloudina hartmannae are of smaller size, indistinguishable from the type specimens of C. riemkeae, which originally was distinguished by an alleged size difference (Germs, Reference Germs1972). The size statistics of the collar width shows that the diameter of Cloudina hartmannae has a size peak ~500 μm (Fig. 2.2). The size peak at 3000 μm is contributed by Cloudina carinata (Fig. 2.2; Supplemental data).

Figure 2. (1) Scatter plot of collar width and spacing in Cloudina. It is noted that the collar widths and spacing are relatively proportional for Cloudina; species designations are according to the original publications instead of the taxonomic revision herein. Some specimens originally described with genus designation were identified to species level herein. (2) Frequency distribution histograms of the diameters of Cloudina hartmannae and Cloudina carinata; note the peak at 3000 μm reflects specimens of Cloudina carinata (in red). The data of Cloudina hartmannae (see original data and relevant references in the supplementary file) are based on the new taxonomic assignment in this study. Abbreviations: C. = Cloudina; N. = Nevadatubulus; H. = Holotype.
Remarks
Germs (Reference Germs1972) named two species, Cloudina hartmannae and Cloudina riemkeae, from Namibia, attributing a Precambrian age to these organisms. The holotype of C. hartmannae (Fig. 1.1) shows weakly preserved collars and a pervasive recrystallization of calcite crystallites. The type specimen of Cloudina riemkeae (Fig. 1.2) is also preserved in a sample with strongly recrystallized spar calcite. The size statistics show that Cloudina has a broad variation (60 to ~4600 μm) of collar width (i.e., the outer diameter). Although sampling and preservational bias may have led to relatively fewer large specimens presented in thin-sections and publications, the size of holotype of C. riemkeae is well within the size cluster of measured C. hartmannae (Fig. 2.1). Although morphologic details are obscured as a result of recrystallization, the preserved characters demonstrate that C. riemkeae shares the same tubular collared-structures with C. hartmannae (Fig. 1.1) and contradicts the original observation of strong size differentiation (Germs, Reference Germs1972). We therefore do not see an objective basis for a distinction of C. riemkeae, and hence support Yang et al. (Reference Yang, Steiner, Zhu, Li, Liu and Liu2016) in synonymizing it with C. hartmannae.
Aulophycus lucianoi Beurlen and Sommer, Reference Beurlen and Sommer1957, was initially reported as a middle–late Cambrian alga from the Tamengo Formation of Corumbá Group, Brazil, although the specimen donor, Dr. Luciano Jacques de Morais, considered it to be of animal origin (Beurlen and Sommer, Reference Beurlen and Sommer1957). The original description (translated from the original description in Portuguese) of the type materials is: “…These are tubiform stems, straight or somewhat curved, not segmented, from less than 1 mm to more than 5 mm in diameter, with wall of variable thickness, thin or not; in the specimens preserved in relief, the wall appears wrinkled, in the polished slabs these wrinkles are represented by thin, sinuous lines. In the same polished surfaces, we find transverse and oblique cuts, frequently, showing several surrounding layers, sometimes concentric or not, with cross-sections presenting circular, round, polygonal and oval contours. Absence of preferential orientation within the strata bearing the clusters of specimens analysed, where alternate beds of higher incidence with other with sparse stems (st. I; V a, b). Stand out the terminations of the stems for their rounded ends (est. II a; III b; V b). The carefully studied sample surfaces do not authorize to speak of ramifications of stalks, nor isolated nodules. The analysed figures don't show that it cannot be interpreted as section made through a circular or oval tube, with closed ends or not, rounded or slightly tapered, the whole stem rarely reaching its 10 mm in length…” (Beurlen and Sommer, Reference Beurlen and Sommer1957, p. 32).
Zaine and Fairchild (Reference Zaine and Fairchild1987) subsequently re-combined A. lucianoi to Cloudina lucianoi. Recent studies further suggested the synonymy of Cloudina lucianoi with Cloudina hartmannae (Adorno et al., Reference Adorno2017, but see Becker-Kerber and Pacheco, Reference Becker-Kerber and Pacheco2018), and modified the systematic treatment, considering Cloudina hartmannae to be a junior synonym of Cloudina lucianoi or vice versa (Yang et al., Reference Yang, Steiner, Zhu, Li, Liu and Liu2016). An examination of type specimens of Aulophycus lucianoi (Fig. 1) is carried out herein. Considering Aulophycus lucianoi was later reassigned from algae to metazoans (e.g., Hahn and Pflug, Reference Hahn and Pflug1985; Zaine and Fairchild, Reference Zaine and Fairchild1987), the holotype sample (D.G.M. 1149) containing multiple tubular remains is therefore treated as an assemblage of syntypes (ICZN, 1999, Article 72.1) and examined here (Fig. 1.3–1.9; image credit to Dr. Rafael Costa da Silva from Earth Sciences Museum—Brazilian Geological Survey). The specimens show circular cross sections with single or angularly folded multiple walls, but none of them can be undoubtedly assigned to Cloudina because they lack collar structures. In consideration with the other type specimens and their original descriptions (see above, Beurlen and Sommer, Reference Beurlen and Sommer1957), most of them (Fig. 1.5, 1.8) may be assigned to Sinotubulites or other unknown taxa based of their corrugated multi-layered walls in cross-section. Nonetheless, it is possible that a few of the specimens (on other type samples, but not the holotype sample) might be compared to Cloudina (e.g., specimens on paratype sample 1157). Hahn and Pflug (Reference Hahn and Pflug1985) tentatively suggested that A. lucianoi was synonymous with Cloudina waldei Hahn and Pflug, Reference Hahn and Pflug1985, while the latter is herein reassigned to Sinotubulites baimatuoensis (see discussion below). We thus suggest that Cloudina hartmannae should be retained as a species and not synonymized with Aulophycus lucianoi.
It is noted that the correction of C. hartmannae to C. hartmanae (e.g., Glaessner, Reference Glaessner1976; Adorno et al., Reference Adorno2017) is considered invalid. Germs (Reference Germs1972) initially present a corrected spelling of the source name (Professor Olga Hartman) with a consistent presentation of the taxonomic name C. hartmannae through the text. Thus we consider this species name is after an inappropriate latinization and should be considered valid according to the ICZN (1999, Article 32.5.1).
Cloudina lijiagouensis Zhang et al. in Li et al., Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992, is defined by a single specimen with a spherical shell adhered to the collared tubular body (Li et al., Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992, pl. 8, fig. 6). It is considered here that the spherical shell represents a branch fragment of the main tubular body, as reported commonly in other studies (Hua et al., Reference Hua, Pratt and Zhang2003a; Min et al., Reference Min, Hong, Yaoping and Bo2019a). Cloudina sinensis Zhang et al. in Li et al., Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992, originally was differentiated from Cloudina hartmannae based on its smaller diameter, a lack of dichotomous branching, and ornamentations on the surface. Size statistics revealed the broad range of Cloudina hartmannae (Fig. 2). Dichotomous branching is more common in the three-dimensional specimens from South China compared to those from Namibia (Germs, Reference Germs1972; Shore et al., Reference Shore, Wood, Curtis and Bowyer2020) and South America (Warren et al., Reference Warren, Quaglio, Riccomini, Simões, Poiré, Stríkis, Anelli and Strikis2014), which are usually studied in thin-sections. We agree with the opinion by Cai et al. (Reference Cai, Cortijo, Schiffbauer and Hua2017), synonymizing C. sinensis and C. lijiagouensis with Cloudina hartmannae. Nevadatubulus dunfeei was considered to be distinct from Cloudina hartmannae in size and its coarse layering (Signor et al., Reference Signor, Mount and Onken1987). While its size variation is within the size range of C. hartmannae (Fig. 2.1), the coarse layering is considered to have resulted from taphonomic effect and acid maceration. Due to similarity in the described collar structures, the species is suggested to be synonymized with C. hartmannae (Yang et al., Reference Yang, Steiner, Zhu, Li, Liu and Liu2016; Cai et al., Reference Cai, Cortijo, Schiffbauer and Hua2017). On the other hand, Cloudina carinata, from the Villarta Limestone of the Ibor Group of Spain (Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2010) and Tamengo Formation of Brazil (Adorno et al., Reference Adorno, Walde, Erdtmann, Denezine, Cortijo, Do Carmo, Giorgioni, Ramos and Fazio2019), is distinguished from C. hartmannae by its longitudinal crests. The putatively recrystallized Cloudina carinata in Siberia (Terleev et al., Reference Terleev, Postnikov, Tokarev, Sosnovskaya and Bagmet2011) cannot be unequivocally confirmed by observations through thin-sections. Cloudina cf. carinata (Liang et al., Reference Liang, Cai, Nolan and Xiao2020) needs further examination because it displays irregular longitudinal ridges, which may have resulted from diagenetic alteration during silicification.
Coleolella sp. and other problematic tubular fossils (see synonym list above) from the Deep Spring Formation and the upper Reed Dolomite (White-Inyo Mountains of California and Esmeralda County, Nevada) is reassigned to Cloudina hartmannae (Fig. 5). The Wyattia Taylor, Reference Taylor1966, reported from the Reed Dolomite in the White-Inyo Ranges, USA, also could be synonymous with Cloudina (Grant, Reference Grant1990; Zhuravlev et al., Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov2012). However, the pervasive recrystallization and obscuration of features, such as the collared structure, leave this synonymization tenuous. Reports of Cloudina? sp. from Antarctica (Yochelson and Stump, Reference Yochelson and Stump1977) and Cloudina? borrelloi Yochelson and Herrera, Reference Yochelson and Herrera1974, lack key characteristics of Cloudina, such as collars and multiple layers, rendering this identification dubious. Conotubus varius Zhang et al., 1992 (Li et al., Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992) is suggested to be Cloudina hartmannae by its typical collared structures instead of transversal ridges (Fig. 8.4; see the diagnosis of Sinotubulites below).

Figure 5. Cloudina hartmannae Germs, Reference Germs1972, from the Deep Spring Formation of Mount Dunfee in Nevada, USA. (1) ELG370401. (2) ELG370402. Scale bars are 500 μm. Specimens are deposited at the Institute of Geology, CAGS.
It is emphasized that Cloudina hartmannae has a wide morphological variation in collar size (Figs. 2–4), density, and angles. Multiconotubus Cai et al. (Reference Cai, Cortijo, Schiffbauer and Hua2017) was defined as tubular fossils with multi-layered and nested cone-shaped layers. Compared with most of the known specimens of Cloudina hartmannae, this indicates that it consists of tubes with collars of high density and small divergent angles. This species requires further study to determine if there are genus-level differences or merely inter-/intraspecific variations, as observed among various specimens (e.g., Fig. 3). The Cloudina-like fossil Feiyanella manica Han et al., Reference Han, Cai, Schiffbauer, Hua, Wang, Yang, Uesugi, Komiya and Sun2017, is a tubular fossil with multi-layered shells and wrinkled/folded outer walls, as well as two orders of dichotomous branchings, forming three generations of tubes (Han et al., Reference Han, Cai, Schiffbauer, Hua, Wang, Yang, Uesugi, Komiya and Sun2017). The fossils superficially resemble branching algae, such as Epiphyton from the Dengying Formation of the same area (Min et al., Reference Min, Hua, Liu, Sun, Cui and Jiang2019b). The published Cloudina specimens from Mexico (Sour-Tovar et al., Reference Sour-Tovar, Hagadorn and Huitrón-Rubio2007; Hodgin et al., Reference Hodgin, Nelson, Wall, Barrón-Díaz, Webb, Schmitz, Fike, Hagadorn and Smith2020) show typical corrugated surfaces and irregularly distributed longitudinal and transversal ridges, but the typical collar-structure was missing (McMenamin, Reference McMenamin1985; Sour-Tovar et al., Reference Sour-Tovar, Hagadorn and Huitrón-Rubio2007, text-figs. 2D, E). As a result, most of these individuals can be assigned to Sinotubulites. Cloudina waldei Hahn and Pflug, Reference Hahn and Pflug1985, from the Tamengo Formation, Corumbá Group in southwest Brazil likely developed a “strongly wrinkled outer wall” (Hahn and Pflug, Reference Hahn and Pflug1985, p. 423) instead of collars because the outer wall is described as (translated from the original description in German): “Transverse ridges are interrupted by narrow, irregularly running transverse furrows; transverse ridges can also occur (pl. 1, fig. 1c). In some specimens there are also external longitudinal ridges which, in conjunction with the transverse ridges, can produce a cassette-like field of the outer surface…” (Hahn and Pflug, Reference Hahn and Pflug1985, p. 425). These descriptions, as well as the illustrated specimens (Hahn and Pflug, Reference Hahn and Pflug1985, pls. 1, 2.3, 2.4), support our assignment of the species to Sinotubulites baimatuoensis.
Branching individuals of Cloudina have been reported rarely from South China (Hua et al., Reference Hua, Chen, Yuan, Zhang and Xiao2005; Min et al., Reference Min, Hong, Yaoping and Bo2019a), Spain (Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2010; Cortijo et al., Reference Cortijo, Cai, Hua, Schiffbauer and Xiao2015a), Brazil (Warren et al., Reference Warren, Quaglio, Riccomini, Simões, Poiré, Stríkis, Anelli and Strikis2014), and Namibia (Germs, Reference Germs1972; Shore et al., Reference Shore, Wood, Curtis and Bowyer2020). Considering the strong recrystallization and that post-mortem stacking of specimens could result in apparent branching, the branching features observed in Cloudina specimens from Namibia (Germs, Reference Germs1972; Shore et al., Reference Shore, Wood, Curtis and Bowyer2020) and Brazil (Warren et al., Reference Warren, Quaglio, Riccomini, Simões, Poiré, Stríkis, Anelli and Strikis2014) are awaiting further confirmation. Disproportionate reports for the rarity of branching specimens might indicate the possibility that these specimens indicate particular ecological conditions that forced local development of budding and branching during reproduction in distinct habits, or even that they might be assigned to a different species. Another possible explanation is based on the taphonomic bias (i.e., phosphatic specimens can be extracted in great numbers by acid digestion compared to calcitic specimens), which are mainly observed with thin sections. Furthermore, multiple taphomodes lead to morphological variations of Cloudina. Specifically, three-dimensionally preserved phosphatic specimens exhibit characteristics of closed ends and fine decorations on the shell surfaces, whereas such features cannot be easily observed in thin-sections of calcitic specimens (Figs. 3, 4). Nonetheless, two-dimensional (thin-section) data of the ultrastructure, compositional comparison, as well as statistics of the collar width and spacing (Fig. 2) are equally beneficial for taxonomic understanding. There also exists the possibility that the closed end is simply a diagenetic feature created by a cover of phosphate cement.
Phylum, Class, Order, Family uncertain
Genus Sinotubulites Chen, Chen, and Qian., Reference Chen, Chen and Qian1981
Type species
Sinotubulites baimatuoensis Chen, Chen, and Qian, Reference Chen, Chen and Qian1981, from the Middle (=Shibantan) Member of the Dengying Formation, Shibantan of Yichang City, China, by original designation.
Other species
Sinotubulites triangularis Cai et al., Reference Cai, Xiao, Hua and Yuan2015; Sinotubulites pentacarinalis Cai et al., Reference Cai, Xiao, Hua and Yuan2015; and Sinotubulites hexagonus Cai et al., Reference Cai, Xiao, Hua and Yuan2015.
Emended diagnosis
Straight or curved tubular fossils with variable diameter, decorated with corrugations, transverse annulations, and discrete longitudinal ridges. The annulae of the external decoration partially show Y-shaped arrangements. The shell wall is relatively thick and multi-layered. The cross-section is circular or irregularly polygonal due to the presence of longitudinal ridges. Inner wall is smooth, and the margin of the inner wall is always circular in cross-section.
Sinotubulites baimatuoensis Chen, Chen, and Qian, Reference Chen, Chen and Qian1981
Figures 6, 7.3, 8.2–8.4, 9
- Reference Chen and Wang1977
Cloudina? sp.; Chen and Wang, p. 220, figs. 1a, b (part).
- Reference Chen, Chen and Qian1981
Sinotubulites baimatuoensis Chen et al., p. 119, pl. 1, figs. 1, 2, pl. 2, figs. 1–6 (part).
- Reference Chen, Chen and Qian1981
Skolithos miaoheensis Chen, Chen, and Qian, p. 117, pl. 1, figs. 4, 5 (part).
- Reference Mount, Gevirtzman and Signer1983
Salanytheca sp.; Mount et al., p. 225, fig. 3A.
- ?Reference Mount, Gevirtzman and Signer1983
Salanytheca sp.; Mount et al., p. 225, fig. 3B.
- Reference Signor, McMenamin, Gevirtzman and Mount1983
Salanytheca sp.; Signor et al., p. 416, fig. 3c.
- Reference Signor, McMenamin, Gevirtzman and Mount1983
Smooth, single-walled shell; Signor et al., p. 416, fig. 3f.
- Reference Signor, McMenamin, Gevirtzman and Mount1983
Irregularly annulated tube; Signor et al., p. 416, fig. 3g.
- Reference Signor, McMenamin, Gevirtzman and Mount1983
Regularly annulated tube; Signor et al., p. 416, fig. 3h.
- Reference Hahn and Pflug1985
Cloudina waldei Hahn and Pflug, p. 423, fig. 7, pl. 1, figs. 1, 2, pl. 2, figs. 3, 4.
- Reference McMenamin1985
Sinotubulites cienegensis McMenamin, p. 1417, figs. 3.2–3.6, 4.1, 4.2, 4.4–4.7, 5.2, 5.5, 5.6.
- ?Reference McMenamin1985
Invaginated tubular or narrow conical shells with thin wall; McMenamin, p. 1422, fig. 5.3.
- Reference McMenamin1985
Sinotubulites baimatuoensis; McMenamin, p. 1416, fig. 6.
- Reference Signor, Mount and Onken1987
Sinotubulites cienegensis; Signor et al., p. 431, fig. 5.1.
- ?Reference Signor, Mount and Onken1987
Salanytheca sp.; Signor et al., fig. 5.2.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Qinella shaanxiensis Zhang, Li, and Dong in Li et al., p. 94, pl. 7, figs. 1–5, 8–10, pl. 9, figs. 5, 7, 10.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Qinella levis Zhang, Li, and Dong in Li et al., p. 96, pl. 7, fig. 6.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Qinella lijiagouensis Zhang, Li, and Dong in Li et al., p. 96, text-fig. 6-3, pl. 7, fig. 7.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Qinella cf. lijiagouensis; Li et al., p. 97, pl. 14, figs. 6a–c.
- ?Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Qinella sp.; Li et al., p. 98, pl. 14, fig. 4.
- ?Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Sinotubulites baimatuoensis; Li et al., p. 98, pl. 16, fig. 1.
- Reference Ding, Xing, Wang, Yin and Gao1993
Sinotubulites miaoheensis Ding et al., p. 120, pl. 2, figs.1–6.
- Reference Hua, Zhang, Zhang and Wang2000a
Qinella levis; Hua et al., p. 386, pl. 1, figs. 1, 3, 4, 9–12.
- Reference Hua, Zhang, Zhang and Wang2000a
Sinotubulites cienegensis; Hua et al., p. 386, pl. 1, figs. 5–8, pl. 2, figs. 1–5, 6a, b, 14.
- Reference Hua, Zhang, Zhang and Wang2000b
Sinotubulites cienegensis; Hua et al., p. 513, pl. 1, fig. 1b, pl. 2, fig. 6.
- Reference Chen and Sun2001
Sinotubulites shaanxiensis; Chen and Sun, p. 196, pl. 3, fig. 7, pl. 4, figs. 1, 2.
- Reference Hua, Pratt and Zhang2003a
Sinotubulites cienegensis; Hua et al., p. 456, figs. 2A–C.
- Reference Hua, Zhang and Chen2003b
Morphotype I; Hua et al., p. 197, pl. 1, fig. 2.
- Reference Hua, Chen and Yuan2007
Sinotubulites; Hua et al., p. 269, figs. 5.7, 5.8.
- Reference Hua, Chen and Yuan2007
silica-replaced cloudiniids; Sour-Tovar et al., p. 171, text-figs. 2D, E.
- Reference Chen, Bengtson, Zhou, Hua and Yue2008
Sinotubulites; Chen et al., p. 39, figs. 2A–C, E, 3A–G, 4A–G.
- Reference Cai, Hua, Xiao, Schiffbauer and Li2010
Sinotubulites baimatuoensis; Cai et al., p. 492, fig. 4N.
- ?Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov2012
Sinotubulites sp.; Zhuravlev et al., p. 212, fig. 5.
- Reference Sun, Hua and Cai2012
Sinotubulites; Sun et al., p. 110, figs. 3, 4.
- Reference Cai, Xiao, Hua and Yuan2015
Sinotubulites baimatuoensis; Cai et al., p. 14, figs. 3, 4.
- Reference Cortijo, Martí Mus, Jensen and Palacios2015b
Sinotubulites baimatuoensis; Cortijo et al., p. 191, fig. 6.
- Reference Xiao, Narbonne, Zhou, Laflamme, Grazhdankin, Moczydlowska-Vidal and Cui2016
Sinotubulites; Xiao et al., p. 544, fig. 4I.
- Reference Walde, do Carmo, Guimarães, Vieira, Erdtmann, Sanchez, Adorno and Tobias2019
Sinotubulites baimatuoensis; Walde et al., p. 345, figs. 9A–E.
- Reference Hodgin, Nelson, Wall, Barrón-Díaz, Webb, Schmitz, Fike, Hagadorn and Smith2020
Cloudina; Hodgin et al., p. 117, figs. 2D, E.
- Reference Liang, Cai, Nolan and Xiao2020
Sinotubulites; Liang et al., p. 6, fig. 6.
- Reference Yang, Shang, Steiner and Liu2020a
Sinotubulites baimatuoensis; Yang et al., p. 450, figs. 2D, E.

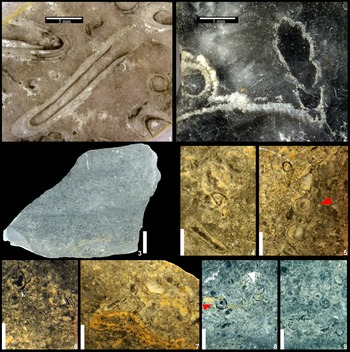
Figure 6. Rescanned original photograph of previous syntypes of Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981, displaying the originally reported specimen from the Middle Member (Shibantan Member), Dengying Formation in the Three Gorges region of Hubei Province, China, YG-96-1. Detailed fossil information is presented in Figures 7 and 8. The numbers along X and Y axes provide precise locations as well as a centimeter scale for individual specimens.

Figure 7. Specimens acquired from the previous type material (YG-96-1, Fig. 6) of Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981. The specimens contain Cloudina hartmannae: (1) smooth tubular fossils (1, 2) as well as Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981, (3). Scale bars are 1 cm. Specimens are deposited at the Geological Museum of China, Beijing, China.
Lectotype hereby designated
YG-96-5 (Fig. 8.4), deposited at Geological Museum of China, Beijing, China.

Figure 8. Detailed views of tubular fossils from the specimens shown in Figure 7. (1) Proximal view of Cloudina hartmannae and smooth tubular fossils (Fig. 7.1, red box, 90° clockwise rotation) with a coordinate of (6.6, 3.5; Fig. 6). (2) Proximal view of one paralectotype of Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981 (white arrow, red box in Fig. 7.3, rotated 90° counterclockwise) showing undeformed annulations. (3) Proximal view of one paralectotype of Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981 (green box in Fig. 7.3, 90° rotation counterclockwise) showing a longitudinal ridge; coordinate of (11.2, 4.6; Fig. 6). (4) Lectotype of Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981, YG-96-5. (5) Proximal view of (4) showing corrugated annulations on the surface. Scale bars: (1–3) 2 mm; (4, 5) 100 μm. Specimens are deposited at the Geological Museum of China, Beijing, China.
Paralectotype hereby designated
The specimens in Figure 8.2 and 8.3, from a hand slab deposited at Geological Museum of China, Beijing, China (original number of whole sample is YG-96-1).
Emended diagnosis
Small straight or slightly curved tubular fossils, millimetric to centimetric size, with surface ornamentation of transversal rings. Irregular rugae, discontinued transversal and longitudinal ridges can be preserved and arranged in Y-shaped patterns. The wall is relatively thick and multi-layered. The inner wall is originally smooth, and the cross-section is circular to oval or polygonal at the outer wall due to discrete longitudinal ridges.
Occurrence
Shibantan Member of Dengying Formation, Yichang City, China (Chen et al., Reference Chen, Chen and Qian1981; Figs. 6–8); Dengying Formation of Shennongjia area of Hubei (Yang et al., Reference Yang, Shang, Steiner and Liu2020a) and South Shaanxi provinces (Chen and Sun, Reference Chen and Sun2001; Chen et al., Reference Chen, Bengtson, Zhou, Hua and Yue2008; Cai et al., Reference Cai, Xiao, Hua and Yuan2015, Reference Cai, Xiao, Li and Hua2019), South China; Deep Spring Formation, Mount Dunfee, Nevada, USA (Mount et al., Reference Mount, Gevirtzman and Signer1983, reported as Salanytheca sp.; Signor et al., Reference Signor, McMenamin, Gevirtzman and Mount1983, reported as Salanytheca sp.); La Ciénega Formation, Sonora, Mexico (McMenamin, Reference McMenamin1985; Signor et al., Reference Signor, Mount and Onken1987; Sour-Tovar et al., Reference Sour-Tovar, Hagadorn and Huitrón-Rubio2007, reported as silica-replaced cloudiniids); Villarta Limestone of Ibor Group, Villarta de los Montes village, Bada-joz Province, Spain (Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2015b); Tamengo Formation, Corumbá Group, Mato Grosso do Sul State, Brazil (Hahn and Pflug, Reference Hahn and Pflug1985, originally reported as Cloudina waldei); Mooifontein Member, Nama Group, Aar Farm, South Namibia (Fig. 9).

Figure 9. Sinotubulites baimatuoensis Chen et al., Reference Chen, Chen and Qian1981, from the Mooifontein Member, Nama Group, Aar Farm, Namibia. (1) Longitudinal section showing corrugated shell surface, A1404; (2) magnified view (1, red box) revealing multi–layered shells; (3) magnified view (2, red box) showing the multilayers on the wall. Scale bars: (1) 500 μm; (2) 100 μm; (3) 50 μm. Specimens are deposited at the Institute of Geology, CAGS.
Description
Straight or sinuous, nearly cylindrical tubular fossils with a length of 2–18 mm. Outer cross sections are round or irregularly polygonal. Inner wall is smooth, circular to oval in cross section. Tube surfaces are ornamented with regular or irregular rugae, or annulations, which are sometimes arranged in a Y-shape. Some of the specimens have longitudinal ridges, which are often developed along the entire length of tubes, but occasionally are interrupted. The tubes have a thick, irregularly corrugated multi-layered wall.
Remarks
Abundant tubular fossils were reported from one silicified dolostone bed within the Shibantan Member of Dengying Formation of the Three Gorges Region and identified as Cloudina? sp. (Chen and Wang, Reference Chen and Wang1977). The subsequent study (Chen et al., Reference Chen, Chen and Qian1981) recognized morphological differences between these fossils and Cloudina, leading to the establishment of Sinotubulites baimatuoensis. Chen et al. (Reference Chen, Chen and Qian1981) described the hand-specimen “YG-96-1” (Fig. 6) containing multiple individuals as the genotype of Sinotubulites. This designation is invalid according to the regulation on genotype (ICZN, 1999). Therefore, all the described specimens of S. baimatuoensis presented in the study of Chen et al. (Reference Chen, Chen and Qian1981) are regarded as syntypes. The repository of the type materials was unknown for a long time, but recently, syntype materials of Sinotubulites baimatuoensis were recovered at the Geological Museum of China, allowing us to carry out a detailed re-examination (Figs. 7, 8). The rediscovered samples include the specimen “YG-96-5” (with part of it lost) and one small specimen preserved without numbering, as well as a rock slab “YG-96-1,” which is heavily silicified, showing limited details of the fossils. Given the varied preservation of all these specimens, we designate the specimen “YG-96-5” as the lectotype and the previous neotype (Cai et al., Reference Cai, Xiao, Hua and Yuan2015) is considered invalid (see ICZN, 1999, Article 75.8). Other existing type specimens are converted into paralectotypes. After its original documentation (Fig. 6, refigured from original photographs in Chen et al., Reference Chen, Chen and Qian1981), the specimen “YG-96-1” rock slab was later sectioned into four pieces by the original authors, with one piece, unfortunately, now missing. Each of the remaining three parts contains several tubular fossils (Fig. 7). After a careful examination of the syntype sample, we were able to recognize three distinct morphotypes of tubes (sensu Chen et al., Reference Chen, Chen and Qian1981) in the sample. One morphotype with typical collar structures can now be assigned to Cloudina hartmannae (Fig. 8.1). Another morphotype, with corrugations, ridges, and rings, is designated as Sinotubulites baimatuoensis (Fig. 8.2, 8.3). The last type, with a smooth tube surface, is considered as a steinkern or weathered specimen of unknown tubular fossils.
Cai et al. (Reference Cai, Xiao, Hua and Yuan2015) proposed new species of Sinotubulites based on different aperture shapes, including: S. triangularis Cai et al., Reference Cai, Xiao, Hua and Yuan2015; S. pentacarinalis Cai et al., Reference Cai, Xiao, Hua and Yuan2015; and S. hexagonus Cai et al., Reference Cai, Xiao, Hua and Yuan2015. Sinotubulites tubes with only a single ridge developed and specimens with transitional characteristics have been mentioned (Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2015b), raising doubts about how to distinguish between different Sinotubulites species. The formation of ridges and the morphology of the irregular aperture may have resulted from taphonomic effects. The lamellate walls of the tubes are irregularly corrugated, suggesting that they were once flexible and easily deformed, and might be considered as originally organic in composition. In addition, it has been suggested that Corumbella from South America resembles Sinotubulites and can be classified as a sinotubulitid due to morphological and ultrastructural similarities (Wood et al., Reference Wood, Ivantsov and Zhuravlev2017b; Walde et al., Reference Walde, Weber, Erdtmann and Steiner2019). Detailed examination of the two taxa are necessary to test this hypothesis.
Previous studies provided inconclusive proposals for the biological affinities of Sinotubulites, with both Chen et al. (Reference Chen, Chen and Qian1981) and Chen and Sun (Reference Chen and Sun2001) tentatively suggesting a potential annelid affinity. On the other hand, there are differences between Sinotubulites and dwelling tubes of modern annelids (Chen et al., Reference Chen, Bengtson, Zhou, Hua and Yue2008). Sinotubulites is often found in association with Cloudina, and both are suspected to be originally of organic composition with multiple lamellae (Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b). More studies on the lamellar structure and composition are required to understand phylogenetic affinity of Sinotubulites.
Phylum, Class, Order, Family uncertain
Genus Conotubus Zhang and Lin in Lin et al., Reference Lin, Zhang, Tao, Wang and Zhang1986
Type species
Conotubus hemiannulatus Zhang and Lin in Lin et al., Reference Lin, Zhang, Tao, Wang and Zhang1986 from the Lower Member (fifth bed = Gaojiashan Member) of Gaojiashan Formation, Hujiaba Town, Ningqiang County, Shaanxi, China, by original designation.
Other species
The type species only.
Diagnosis
As for the type species by monotypy.
Conotubus hemiannulatus Zhang and Lin in Lin et al., Reference Lin, Zhang, Tao, Wang and Zhang1986
Figure 10
- Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov1986
Conotubus hemiannulatus Zhang and Lin in Lin et al., p. 12, pl. 1, figs. 6, 10.
- Reference Zhang1986
Conotubus hemiannulatus; Zhang, p. 75, pl. 1, figs. 10, 13.
- Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov1986
Conotubus gaojiashanensis Zhang and Lin in Lin et al., p. 12, pl. 1, figs 3–5, 7.
- Reference Zhang1986
Conotubus gaojiashanensis; Zhang, p. 76, pl. 1, figs. 1, 3, 9.
- Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov1986
Conotubus impolitus Zhang and Lin in Lin et al., p. 12, pl. 1, figs. 1, 11.
- Reference Zhang1986
Conotubus impolitus; Zhang, p. 76, pl. 1, fig. 11.
- Reference Zhuravlev, Linan, Vintaned, Debrenne and Fedorov1986
Conotubus mimicus Zhang and Lin in Lin et al., p. 12, pl. 1, figs. 2, 9a, pl. 2, fig. 9.
- Reference Zhang1986
Conotubus mimicus; Zhang, p. 76, pl. 1, fig. 6.
- Reference Zhang1986
Conotubus hamatus Zhang, p. 76, pl. 1, figs. 2, 4.
- Reference Zhang1986
Conotubus cephalotus Zhang, p. 77, pl. 1, fig. 7.
- Reference Zhang1986
Conotubus adpressus Zhang, p. 77, pl. 1, figs. 8, 14, pl. 2, fig. 7.
- Reference Zhang1986
Conotubus sp.; Zhang, p. 77, pl. 1, fig. 12.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus hemiannulatus; Li et al., p. 91, pl. 11, figs. 9, 16, 18.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus gaojiashanensis; Li et al., p. 91, pl. 11, fig. 13, pl. 12, figs. 1, 4.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus impolitus; Li et al., p. 92, pl. 11, fig. 14.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus mimicus; Li et al., p. 92, pl. 11, fig. 2, pl. 12, fig. 2.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus hamatus; Li et al., p. 92, pl. 11, fig. 15.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus cephalotus; Li et al., p. 93, pl. 12, fig. 10.
- Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus adpressus; Li et al., p. 93, pl. 12, figs. 3, 5, 6.
- [non] Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992
Conotubus varius Zhang et al. in Li et al., p. 92, text-figs. 3–7, 8:9, pl. 9, figs. 3, 4, 6, 9.
- Reference Zhang and Hua2000
Conotubus hemiannulatus; Zhang and Hua, p. 329, pl. 1, fig. 8.
- Reference Zhang and Hua2000
Conotubus sp.; Zhang and Hua, p. 332, pl. 1, fig. 16.
- Reference Hua, Zhang, Zhang and Wang2000b
Conotubus sp.; Hua et al., p. 513, pl. 1, fig. 7a.
- Reference Cai and Hua2007
Conotubus hemiannulatus; Cai and Hua, p. 647, fig. 1.
- Reference Cai and Hua2007
Conotubus sp.; Cai and Hua, p. 648, fig. 3c.
- Reference Hua, Chen and Yuan2007
Conotubus; Hua et al., p. 269, figs. 5:1–5:3, 6:1, 6:2, 7:2, 7:3, 7:5, 8:2a.
- Reference Cai and Hua2008
Conotubus sp.; Cai and Hua, p. 217, figs. 1G, H.
- Reference Cai, Hua, Xiao, Schiffbauer and Li2010
Conotubus; Cai et al., p. 492, figs. 4B–F, 8A, B, 9A, 9C–J, 10A, B, 10G–I.
- Reference Cai, Schiffbauer, Hua and Xiao2011
Conotubus hemiannulatus; Cai et al., p. 49, figs. 5A, D, E, 6, 7.
- [non] Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016
Conotubus; Smith et al., p. 913, figs. 3A, B.
- Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b
Conotubus hemiannulatus; Yang et al., p. 49, fig. 5C1.
Lectotype hereby designated
V1-21 (Fig. 10; numbered as XG15 in Li et al., Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992, pl. 11, fig. 9), deposited at the Department of Geology, Northwest University, Xi'an, China.

Figure 10. Lectotype of Conotubus hemiannulatus Zhang and Lin in Lin et al., Reference Lin, Zhang, Tao, Wang and Zhang1986, V1–21. (1) General view of the lectotype showing strong and fine annulations; (2) lateral view of (1) revealing that the specimen has been compressed; (3) opposite view of (1) showing an uneven facet with no ridges or annulations preserved; (4) magnified view of (1, white arrow) showing the strong and fine annulations; (5) magnified view of the sectioned part of (2, white arrow) indicating a pyritized composition and unclear multilayer structure. Specimens are deposited at the Department of Geology, Northwest University, China. Scale bars: (1–3) 1 cm; (4) 2 mm; (5) 1 mm.
Emended diagnosis
Straight or curved tubular fossils decorated with corrugations and concentric annulations on the surface. The tube wall is composed of multiple layers with smooth inner wall. The cross section is oblate to round.
Occurrence
Gaojiashan Member of the Dengying Formation, Ningqiang County, South Shaanxi, China (Lin et al., Reference Lin, Zhang, Tao, Wang and Zhang1986; Li et al., Reference Li, Ding, Zhang, Dong, Chen, Ding, Zhang, Li and Dong1992; Hua et al., Reference Hua, Chen and Yuan2007; Cai et al., Reference Cai, Schiffbauer, Hua and Xiao2011).
Description
Oblate tapered tubes with a length of 40–80 mm and width of 4–10 mm. The fossils are slightly curved, with cross sections oblate or oval. The tubes show ridges and annulations on the surface, which are raised to the tapered end. The tube wall is composed of two or more laminae.
Remarks
Conotubus hemiannulatus Zhang and Lin in Lin et al., Reference Lin, Zhang, Tao, Wang and Zhang1986 was originally defined without a designation of holotype or paratype. Following ICZN (1999, Articles, 72.1.1, 73.2), the only two specimens of Conotubus hemiannulatus illustrated in the original publication became syntypes. The location of the two syntypes has been unknown for a long time. Here, we report one of these two syntypes from the collections of Northwest University, Xian, China, designating it as a lectotype. Based on characters such as prominent ridges and annulations of the lectotype, the specimens reported by Cai et al. (2011, figs. 3C, D) may not belong to Conotubus, but show a similarity to tubular fossils with collar structures (e.g., Saarina from Nevada; Selly et al., Reference Selly, Schiffbauer, Jacquet, Smith and Nelson2020; or possibly Cloudina). The cf. Cloudina sp. reported from the Wood Canyon Formation in the Death Valley region (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000) is probably comparable to the subsequently reported Conotubus from the same deposits (Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017), as well as the Deep Spring Formation of Mt. Dunfee, Nevada (Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016). Both of them resemble Costatubus Selly et al., Reference Selly, Schiffbauer, Jacquet, Smith and Nelson2020, Saarina Sokolov, Reference Sokolov and Sokolov1965, and Zuunia chimidtsereni Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b, in overall morphology as well as collar-structures.
The lectotype of Conotubus indicates a limonitic composition, likely resulting from diagenetic replacement of an original carbonaceous composition of tube walls. The premise for this assumption is that carbonaceous Conotubus has been reported from the Dengying Formation in South Shaanxi, China, and interpreted as precursor (ancestor) of Cloudina (Hua et al., Reference Hua, Chen and Yuan2007). Considering that Cloudina was interpreted as primarily organic in composition (Yang et al., Reference Yang, Steiner, Schiffbauer, Selly, Wu, Zhang and Liu2020b), another interpretation about these carbonaceous specimens is that they may represent a taphomorph of Cloudina preserved without secondary mineralization. This potential interpretation generally calls for a further examination of the carbonaceous Conotubus from South Shaanxi.
Although Lin et al. (Reference Lin, Zhang, Tao, Wang and Zhang1986) suspected that Conotubus is related to annelids by comparing it with Cretaceous fossils Diploconcha Conrad, Reference Conrad and Kerr1875, and Longitubus Howell, Reference Howell1943, other studies considered it closely related to Cloudina (Hua et al., Reference Hua, Chen and Yuan2007; Cai et al., Reference Cai, Schiffbauer, Hua and Xiao2011), with an undetermined phylogenetic affinity. Thus, its phylogenetic affinity remains uncertain.
Discussion
Based on the detailed taxonomic description, we propose that the genus Cloudina consists of Cloudina hartmannae Germs, Reference Germs1972, Cloudina carinata, Cortijo et al., Reference Cortijo, Martí Mus, Jensen and Palacios2010, and Cloudina xuanjiangpingensis Cai et al., Reference Cai, Cortijo, Schiffbauer and Hua2017. We further suggest that the tubular remains in the type specimens of Aulophycus lucianoi may be compared to Sinotubulites or other taxa. Lectotypes of Sinotubulites baimatuoensis and Conotubus hemiannulatus are designated. Cloudina waldei from the Tamengo Formation, Corumbá Group in Southwest Brazil is tentatively assigned to Sinotubulites baimatuoensis. Moreover, S. baimatuoensis is reported from the Mooifontein Member of Nama Group in Aar Farm, Namibia.
Considering their narrow temporal range, broad spatial distribution, multiple taphomodes, and various sedimentary facies, Ediacaran tubular fossils have been proposed as index fossils for the late Ediacaran (Grant, Reference Grant1990; Gaucher and Germs, Reference Gaucher, Germs, Gaucher, Sial, Halverson and Frimmel2009; Xiao et al., Reference Xiao, Narbonne, Zhou, Laflamme, Grazhdankin, Moczydlowska-Vidal and Cui2016; Adorno et al., Reference Adorno2017), although difficulties exist in biostratigraphic correlations because of their unresolved taxonomic status. Following the revision of the three taxa (Cloudina, Sinotubulites, and Conotubus), some of these tubular fossils are more acceptable for stratigraphic correlation and paleobiogeographic reconstruction (Fig. 11). The first appearance datum (FAD) of Cloudina hartmannae is considered approximately time-equivalent and has been applied as an index fossil for the late Ediacaran. Gaucher and Germs (Reference Gaucher, Germs, Gaucher, Sial, Halverson and Frimmel2009) proposed a specific biozone based on the cloudinid appearances—the Cloudina Taxon Range Zone, ranging from >550–542 Ma for the Nama Group in Namibia, Ara Group in Oman, Corumbá Group in Brazil, and the Arroyo del Soldado Group in Uruguay. The coexistence of Cloudina with typical early Cambrian small shelly fossils, such as Anabarites and Protohertzina (Yang et al., Reference Yang, Steiner, Zhu, Li, Liu and Liu2016; Zhu et al., Reference Zhu, Zhuravlev, Wood, Zhao and Sukhov2017; Cai et al., Reference Cai, Xiao, Li and Hua2019), demonstrates that the taxon range zone extends to the early Cambrian. Later on, the Cloudina-Namacalathus-Sinotubulites Assemblage Zone (Zhu et al., Reference Zhu, Zhuravlev, Wood, Zhao and Sukhov2017) was suggested, preceding the early Cambrian assemblage zone SSF I (Anabarites trisulcatus-Protohertzina anabarica Assemblage Zone). Also due to the range of Cloudina into early Cambrian strata, this assemblage zone requires a modification. Consequently, we redefine the terminal Ediacaran biozone as the Cloudina hartmannae Interval Zone based on species-level taxa. The zone is defined as the interval between the first appearance datum of Cloudina hartmannae and the first appearance datum of Protohertzina anabarica (i.e. the index fossil of the early Cambrian Anabarites trisulcatus-Protohertzina anabarica Assemblage Zone). The zone is also characterized by the co-occurrence of the species Cloudina hartmannae, Sinotubulites baimatuoensis, and Namacalathus hermanastes Grotzinger, Watters, and Knoll, Reference Grotzinger, Watters and Knoll2000 (Warren et al., Reference Warren, Quaglio, Simões, Gaucher, Riccomini, Poiré, Freitas, Boggiani and Sial2017). The age range of this biozone begins at ca. 550 Ma (Grotzinger et al., Reference Grotzinger, Bowring, Saylor and Kaufman1995) and ends at ca. 541 Ma, or 539 Ma with new data from Namibia (Linnemann et al., Reference Linnemann, Ovtcharova, Schaltegger, Gärtner and Hautmann2019). This zone is defined as an interval zone to avoid uncertainties with the range of Ediacaran tubular fossils into the Cambrian strata because there have been reports of the tubular fossils ranging into the basal Cambrian (Yang et al., Reference Yang, Steiner, Zhu, Li, Liu and Liu2016; Zhu et al., Reference Zhu, Zhuravlev, Wood, Zhao and Sukhov2017; Cai et al., Reference Cai, Xiao, Li and Hua2019). The definition of an interval zone must be seen as a compromise due to the rarity of other characteristic and widely distributed fauna in the Ediacaran, which might serve for a definition as an assemblage zone.

Figure 11. Paleogeographic reconstruction of Gondwana at ca. 541 Ma highlighting the distribution of Cloudina hartmannae Germs, Reference Germs1972, Sinotubulites baimatuoensis Chen et al., 1987, and Namacalathus hermanastes Grotzinger et al., Reference Grotzinger, Watters and Knoll2000. Base map is modified from Torsvik and Cocks (Reference Torsvik, Cocks, Harper and Servais2013). Occurrences of C. hartmannae and S. baimatuoensis as given in the systematic description paragraphs; occurrences of N. hermanastes according to Penny et al. (Reference Penny, Wood, Zhuravlev, Curtis, Bowyer and Tostevin2017) and Warren et al. (Reference Warren, Quaglio, Simões, Gaucher, Riccomini, Poiré, Freitas, Boggiani and Sial2017). Dashed line marks potential original outlines of Armorica microcontinent, of which only dispersed blocks remain.
Acknowledgments
We are grateful for the support of M.-E. Chen (Beijing), H. Hua (Xi'an), J.-J. Ren (Xi'an), X. Wang (Beijing), M. Denezine (Brasília), and R.R. Adorno (Brasília) for relocating the type materials. S. Mtshali (Cape Town) and R.C. da Silva (Earth Sciences Museum, Brazilian Geological Survey, Brazil) are thanked for providing photographs of Cloudina samples, holotypes, and syntypes. The study is funded by the NSFC (41972026; 41872024), the Strategic Priority Research Program (B) of Chinese Academy of Sciences, (XDB26000000) and China Geological Survey (DD20190008). M. Steiner is supported by DFG (Grant no. STE814/5-1). L.V. Warren is a fellow of the CNPq. The samples from Paraguay were collected during the project FAPESP (grant 2018/26230-6). E.F. Smith acknowledges support from the Smithsonian Institution Peter Buck Fellowship and NSF-1827669. We thank the Nevada Bureau of Land Management for a collection permit to E. Smith for the Nevada specimens.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.xgxd254hb.