


INTRODUCTION
The blue shark (Prionace glauca Linnaeus, 1758) is characterized as having a higher number of embryos and embryonic malformations than any other species of elasmobranchs. This study reports for the first time four malformations of blue shark embryos from the western coast of Baja California Sur (BCS), Mexico. The blue shark is an elasmobranch (family Carcharhinidae) with wide distribution in tropical and temperate areas of the Atlantic, Pacific and Indian Oceans. In the Pacific Ocean, the blue shark inhabits pelagic and neritic waters (4–37°N and 104–157°W) in temperatures from 14 to 27 °C (Holts et al., Reference Holts, Wilson and Lowe2001; Weng et al., Reference Weng, Castilho, Morrissette, Landeira-Fernández, Holts, Schallert, Goldman and Block2005). It is the most abundant pelagic shark and a main resource for several fisheries in the world (Carey & Scharold, Reference Carey and Scharold1990; Nakano & Stevens, Reference Nakano, Stevens, Camhi, Pikitch and Babcock2008), and feeds mainly on mesopelagic cephalopods and crustaceans (Hernández-Aguilar et al., Reference Hernández-Aguilar, Escobar-Sánchez, Galván-Magaña and Abitia-Cárdenas2016). Along the western coast of BCS, blue shark are in high demand for industrial and artisanal fisheries, representing 63% of the total catch in the region (Galván-Magaña, Reference Galván-Magaña, Urciaga, Beltrán and Lluch2009). This fishery faces management problems with overfishing because catches are predominately immature sharks, motivating the need for further research (Sosa-Nishizaki et al., Reference Sosa-Nishizaki, Márquez-Farías, Villavicencio-Garayzar, Camhi, Pikitch and Babcock2008). The catch size reported has been 77–274 cm TL (Cruz et al., Reference Cruz, Soriano, Santana, Ramirez and Valdez2011) while size at first maturity in females is 186–212 cm TL (Nakano & Seki, Reference Nakano and Seki2003). The blue shark is a viviparous species with a gestation period of 9–12 months and high fecundity (Pratt, Reference Pratt1993; Hazin et al., Reference Hazin, Pinheiro and Broadhurst2000). Females produce between 5 and 52 offspring every two years with one year of gestation and one year of repose (Compagno, Reference Compagno1984; Hamlett, Reference Hamlett2005). Previous studies in blue shark have suggested that the short space in female wombs and the large number of offspring in this species may be responsible for malformations (Galván-Magaña et al., Reference Galván-Magaña, Escobar-Sánchez and Carrera-Fernández2011). Additionally, malformations may be the result of genetic alterations during embryonic development or by pollution in the area of species distribution (Whitehead et al., Reference Whitehead, Dubansky, Bodinier, Garcia, Miles, Pilley, Raghunathan, Roach, Walker, Walter, Rice and Galvez2012). Early studies on malformations in sharks have reported deformities mainly in skeletal traits, such as pectoral fins not fused to the head, albinism and dicephaly and trunk rolled in the case of rays (Heupel et al., Reference Heupel, Simpfendorfer and Bennett1999; Mancini et al., Reference Mancini, Casas and Amorim2006; Ochoa-Diaz et al., Reference Ochoa-Diaz, Rodriguez-Romero, López-Martínez and Maldonado-García2016). This study reports for the first time four malformations of blue shark embryos from the western coast of BCS, Mexico.
MATERIALS AND METHODS
The samples were caught by the industrial fishing boat ‘Damasta’, which operates along the coast of the Pacific Ocean from Punta Abreojos to Cabo San Lucas, BCS, Mexico, encompassing the coordinates 26–27°N and 110–112°W in 2012 and 2013 (Figure 1).
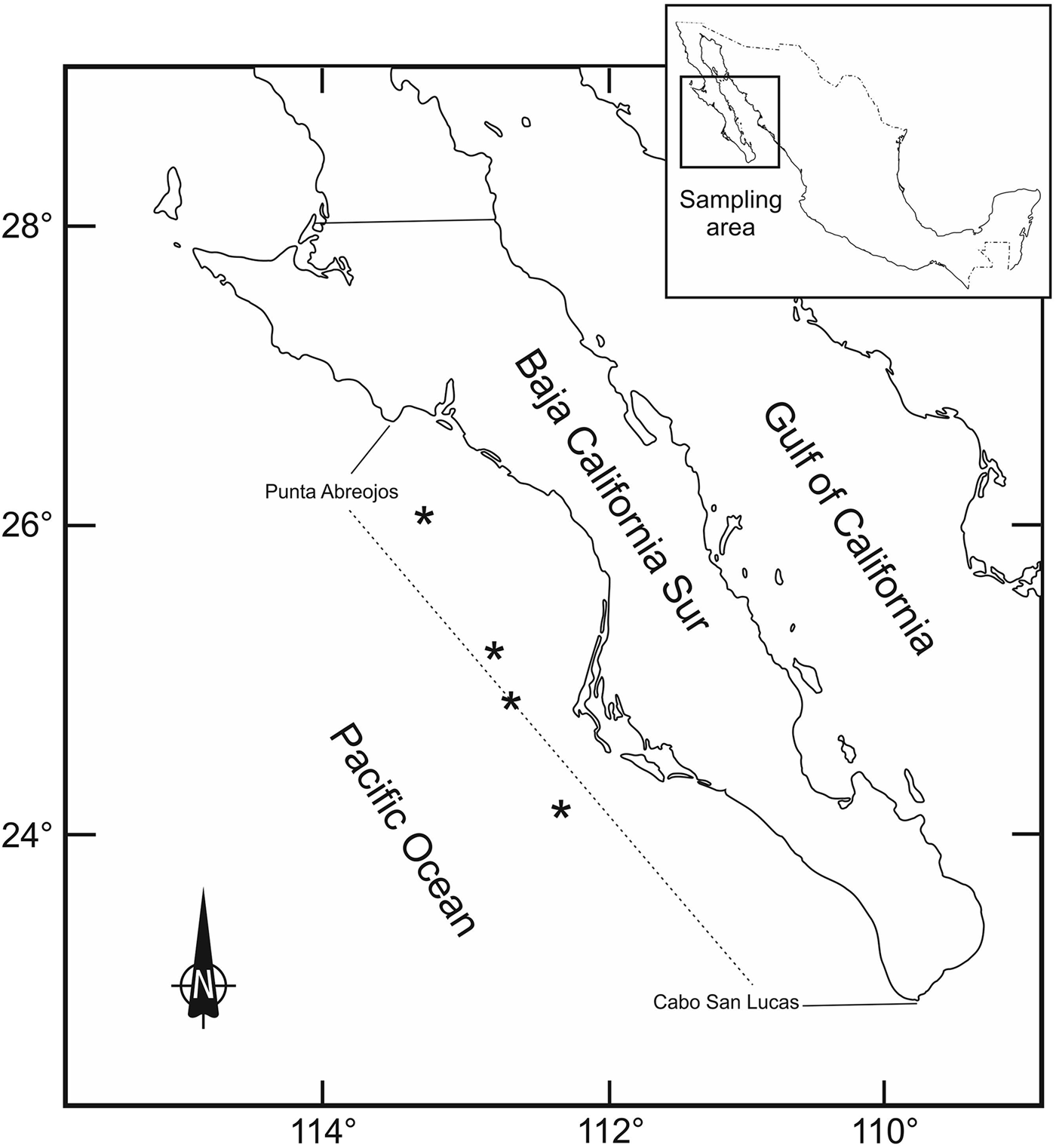
Fig. 1. Sampling area of the blue shark (Prionace glauca) embryos from the western coast of Baja California Sur, Mexico.
The sharks were caught using long line gear with hooks (no. 9) at a distance of 40–50 km off the coast and at depths of 90–120 m. Surface temperature ranged from 22.7 to 23.3 °C. Embryos obtained from the adult sharks were stored at −4 °C until their examination in the laboratory. Samples were transferred to the Fish Ecology Laboratory at Centro de Investigaciones Biologicas del Noroeste (CIBNOR) where morphometric data were taken (Table 1); malformations were observed, photographed and also dissected to detect any internal alteration. Unfortunately, sizes of mothers were not measured at that time due to the fast work of the fishermen. Malformed embryos were from four different female sharks. Although normal embryos were not sampled, size at birth is normally from 35 to 40 cm on average (Pratt, Reference Pratt1979; Stevens, Reference Stevens1984; Carrera-Fernandez et al., Reference Carrera-Fernandez, Galván-Magaña and Ceballos-Vázquez2010).
Table 1. Morphometric measurements (cm) of blue shark (Prionace glauca) embryos from the western coast of Baja California Sur, México.

RESULTS
Four new malformations and one dicephaly of blue shark embryos were observed from female sharks caught along the western coast of Baja California Sur, Mexico. The first malformations were observed in two female embryos caught in 2012, which had one nostril and short and crushed snouts. The embryos were 22 and 31 cm total length (TL); the first one had a screw-shaped caudal fin (Figure 2). Both of them showed marks of the umbilical cord, a spiracle in each side of the head, and the chondrocranium was flat and short.

Fig. 2. Two female blue shark (Prionace glauca) embryos with one nostril from the western coast of Baja California Sur, Mexico.
The second malformation was observed on board in one female embryo (22.5 cm TL) also caught in 2012, showing two mouths (Figure 3), one on the right side and the other one on the left side. The embryo had a hole that seemed to be a spiracle at the top of the mouth on the right side. The nictitating membrane of the left eye was partially developed, and the middle part of the spine was screw-shaped. After examination of the internal organs, this embryo was found to have four livers; one of them was atrophied and separated by a thin membrane (Figure 3A). The stomach, intestine and heart were developed normally.

Fig. 3. Female blue shark (Prionace glauca) embryo with two mandibles; (A) internal organs showing livers from the western coast of Baja California Sur, Mexico.
The third and fourth malformations were observed in specimens caught in 2013. The third one was in a female embryo with dicephaly, one head bigger than the other, the right one with 12.5 cm TL and the left one 12 cm TL. The fusion of the two heads was from the gill slits. The embryo had five gill slits (one on each side) (Table 1); five pelvic fins (one set with three fins and the other one with two; Figure 4). This embryo showed one pair of the first dorsal while the second pair was absent; it also had two joint caudal fins; two inferior lobules and the edges of the superior ones were separated, showing two vertebral columns that were joint below the last part of the caudal fin (Figure 4A). Dark colouration was observed on the top of the rostrum on the edge of the pectoral, anal and caudal fins while the rest of the body was pale. Some dermal denticles in the head and a distortion in the pre-caudal section were observed. The side of the embryo with the small head showed one undeveloped liver, and the other side had two. Both heads of the embryo shared the liver and intestine. Reproductive organs were not developed.
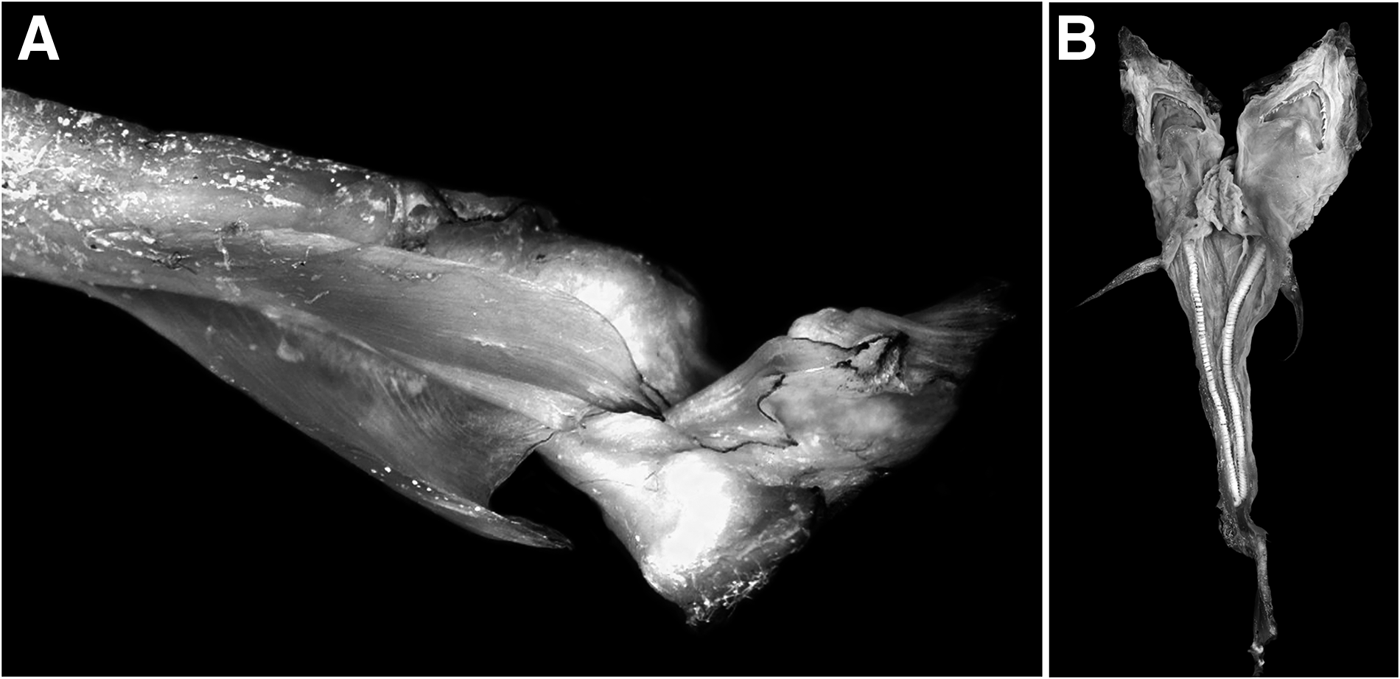
Fig. 4. Male blue shark (Prionace glauca) embryo with dicephaly, two pairs of caudal fins, and three anal fins; (A) caudal fin vertebrae from the western coast of Baja California Sur, Mexico.
The fourth malformation was observed as an incomplete development (i.e. hypoplasia) of a female embryo (10 cm TL). The embryo had a crushed snout with only three gill slits on each side of the body (Figure 5), and the internal organs were completely formed.

Fig. 5. Blue shark Prionace glauca embryo with incomplete growth from the western coast of Baja California Sur, Mexico.
DISCUSSION
The malformations reported in this study contribute to knowledge of new malformations that can take place in this blue shark species. It has been reported that elasmobranchs have immunity to disease by the high level of urea they usually produce (Dosay-Akbulut, Reference Dosay-Akbulut2006). Recent studies of malformations observed in sharks have suggested that malnutrition is associated with deformations in the spine or dicephalism (Delpiani et al., Reference Delpiani, Deli Antoni, Barbini and Figueroa2011) while caudal deformities and severe spinal curvatures have been associated with parasites, arthritis and osteosclerosis (Heupel et al., Reference Heupel, Simpfendorfer and Bennett1999; Olatunji-Akioye et al., Reference Olatunji-Akioye, Adeyemo and Akomolafe2010). Malformation can be associated in some shark species with a large number of embryos, environmental degradation and genetic abnormalities (Mancini et al., Reference Mancini, Casas and Amorim2006; Saïdi et al., Reference Saïdi, Bradaï, Marouani, Guélorget and Capapé2006). High concentration of heavy metals (i.e. Hg) previously found in the muscle of adult blue sharks may be involved in embryonic malformations (Barrera-García et al., Reference Barrera-García, O'Hara, Galván-Magaña, Méndez-Rodríguez, Castellini and Zenteno-Savín2012). Although the cause of embryonic abnormality is still unknown, the individuals reported in this study might not have survived after birth with these conditions due to difficulties in swimming, feeding and evading predators (Bengtsson, Reference Bengtsson1979; Mancini et al., Reference Mancini, Casas and Amorim2006). To our knowledge, the malformations reported here in blue shark embryos in the western Mexican Pacific coast have not been reported previously.
With respect to the first malformation, an embryo with one nostril and spiracles, absent in the adult blue shark (Compagno, Reference Compagno1984), would need its olfactory senses (ampullae of Lorenzini) for mating (Johnson & Nelson, Reference Johnson and Nelson1978; Kajiura et al., Reference Kajiura, Forni and Summers2005), to detect food and prey (Rasmussen & Schmidt, Reference Rasmussen, Schmidt, Doty and Muller-Schwarze1992) and for navigation (Edrén & Gruber, Reference Edrén and Gruber2005).
The second malformation in this study represents the second case for this species and first record for the western coast of BCS, Mexico. The only report of a shark with two jaws and morphological abnormality in the trunk and head region was found previously in a blue shark off the shore of Brazil (Mancini et al., Reference Mancini, Casas and Amorim2006). This malformation might have occurred due to genetic problems associated with development of morphological traits in sharks, especially in early stages of development (Heupel et al., Reference Heupel, Simpfendorfer and Bennett1999).
The third malformation, known as dicephaly (or axial bifurcation), is a developmental anomaly that appears in a single fertilized ovum during gastrulation, often resulting from duplication of the rostral aspect of the neural tube and subsequent development of two heads (De Lahunta et al., Reference De Lahunta, Glass and Marc2014). Previous reports on dicephaly in shark species indicated that the union of the heads occurs in the cephalic region or in the gills (Hevia-Hoarmazábal et al., Reference Hevia-Hoarmazábal, Pastén-Marambio and Vega2011; Wagner et al., Reference Wagner, Rice and Pease2013). Previous malformation reports with dicephalic embryos were made in the fishing area of Punta Arenas in the Gulf of California and San Lazaro Island in the Pacific Ocean, suggesting that the abnormality could have been caused after or during fertilization by an incomplete zygote division (Galván-Magaña et al., Reference Galván-Magaña, Escobar-Sánchez and Carrera-Fernández2011). Other reports of malformation in the cephalic region of three female embryos of blue shark were reported in San Lazaro fishing camp in BCS (Bejarano-Alvarez et al., Reference Bejarano-Alvarez, Galván-Magaña and Ochoa-Báez2011). The liver was shared in one dicephalic embryo. The other two embryos did not have eyes, and part of their snout was not well developed with malformations in cranial cartilage and gill slits. This study suggested that malformations in blue shark embryos happen because this species is the most fecund shark worldwide with the largest number of embryos produced, and not all embryos can develop well in their small uteri. Hevia-Hoarmazábal et al. (Reference Hevia-Hoarmazábal, Pastén-Marambio and Vega2011) described conjoined twins in a specimen of blue shark north of Chile with a partial craniofacial duplication and presence of four eyeballs and an equal number of nostrils. Oral cavities were interconnected, converging in a single pharynx. The trunk was simple; the column presented thoracic lordosis and a helical twist in its caudal portion. Dicephaly was also reported in Galeorhinus galeus Linnaeus, 1758 at Argentina Sea and showed two separate vertebral columns; the first and second dorsal fins were duplicated as well as the dorsal lobe of the caudal fin (Delpiani et al., Reference Delpiani, Deli Antoni, Barbini and Figueroa2011). This embryo was similar to the third embryonic malformation of blue shark reported in our study.
Galván-Magaña et al. (Reference Galván-Magaña, Escobar-Sánchez and Carrera-Fernández2011) deduced that embryonic development continued due to its implantation in the uterus, providing nutrients for the embryo. However, the transfer of nutrients could have been too low to provide sufficient food for ‘two specimens’ (dicephalic shark); therefore, a different growth was observed in both anomalous embryos when compared with normal ones; the small size of anomalous blue sharks was evident.
Malformations associated with incomplete development have been reported for internal organs such as liver and heart (Galván-Magaña et al., Reference Galván-Magaña, Escobar-Sánchez and Carrera-Fernández2011), incomplete external development of eyes and snout (Bejarano-Alvarez et al., Reference Bejarano-Alvarez, Galván-Magaña and Ochoa-Báez2011), anophthalmia (absence of one or both eyes), and the presence of single clasper (Ribeiro-Prado et al., Reference Ribeiro-Prado, Oddone, Gonzalez, Amorim and Capapé2008). In this study, the fourth malformation belonged to incomplete embryonic development and three gill slits while members of the family Carcharhinidae have five (Compagno, Reference Compagno1984). Incomplete development could have been associated with low nutrient input from the mother to the embryo, since sufficient nutrients are essential for the complete development of the offspring (Otake & Mizue, Reference Otake and Mizue1986).
The malformations observed in embryos of blue shark emphasize that the space in the uterus is not enough for embryos to have an optimal development, and that available nutrients are not enough for all the embryos (Galván-Magaña et al., Reference Galván-Magaña, Escobar-Sánchez and Carrera-Fernández2011) although further research is necessary. During the first few weeks of feeding the embryo, nutrients are obtained via the yolk sac, after which the enfeebled vitelline membrane will lead to the creation of the placenta (Carrier et al., Reference Carrier, Pratt, Castro, Carrier, Musick and Heithaus2004).
The blue shark is an important species in fisheries; thus, further studies on heavy metals, genetics and nutrimental contribution of the egg to the embryo are necessary to understand the cause of malformations in embryonic development.
ACKNOWLEDGEMENTS
The authors would like to thank the crew of the ‘Damasta’ and fisherman Antonio Simeón for sampling collection; Claudia J. Perez for helping with the embryo pictures; and Diana Fischer for editorial services.
FINANCIAL SUPPORT
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.






