


Introduction
The prospects of finding a habitable zone terrestrial planet are looking increasingly good. The fraction of Sun-like stars with Earth-like planets in their habitable zones (η⊕) is currently estimated to be between 1.4 and 2.7% (Catanzarite & Shao Reference Catanzarite and Shao2011), and there are tentative suggestions that η⊕ could be as high as 42% for M star planet hosts (Bonfils et al. Reference Bonfils2011). While knowing the conditions that make a planet habitable is clearly extremely useful when searching for life, it is equally useful, although perhaps more pessimistic, to ask the question: when will a planet cease to be habitable? It is entirely possible that some future discoveries of habitable exoplanets will be planets that are nearing the end of their habitable lifetimes, i.e. with host stars nearing the end of their main sequence lifetimes.
An impression of the stages a habitable Earth analogue planet (here taken to be an exact copy of Earth) passes through is given in Fig. 1, while the fraction of known terrestrial planets around late main sequence stars is illustrated in Fig. 2. Knowing the likely conditions on a far-future Earth analogue will help determine whether any life still present on such a planet will be remotely detectable. Due to the harsh environmental conditions such planets would face, it is expected that this life would be microbial.

Fig. 1. Time windows for complex and microbial life on Earth analogue planets orbiting Sun-like stars (F(7), G and K(1) stars) during their main sequence lifetimes. Assuming that the processes leading to multicellular life are the same as on Earth (i.e. ∼1 Gyr for life to emerge and ∼3 Gyr for multicellularity to evolve), the potential life spans of a more complex, multicellular biosphere are estimated. Multicellular life was assumed able to persist until surface temperatures reach the moist greenhouse limit for an Earth analogue planet in the continuously habitable zone of the star-type in question. Microbial life is then assumed to dominate until either the maximum temperature for microbial life is exceeded or until the star leaves the main sequence (whichever happens first). The average age of Earth-like planets was found by Lineweaver (Reference Lineweaver2001) to be ∼6.4 ± 0.9 Gyr based on estimates of the age distribution of terrestrial planets in the universe. This average age falls within microbial and uninhabitable stages for G- and F-type stars, respectively, but falls within the multicellular life stage for K stars.

Fig. 2. Main sequence evolution (relative to the Sun and accounting for stellar type) of currently known terrestrial planet (10ME and less) hosting stars. The left-shaded region represents early microbial biospheres and the right-shaded region represents late microbial biospheres for Earth analogue planets. The central unshaded region represents the stellar age-range within which Earth analogue planets would be more likely to have complex biospheres. The vertical line represents the current age of the Sun. Source: The Extrasolar Planets Encyclopaedia (Schneider Reference Schneider2010).
Life is thought to have emerged on Earth 3.8 Gyr ago (Schidlowski Reference Schidlowski1988; Rosing Reference Rosing1999) – and perhaps even as early 3.85 Gyr ago (Mojzsis et al. Reference Mojzsis, Arrhenius, McKeegan, Harrison, Nutman and Friend1996). Chemical evidence suggests that archaeal or bacterial unicellular organisms were present for at least 2.5 Gyr before body fossil evidence of multicellular life, as the red alga Bangiomorpha, appeared 1.2 Gyr ago (Butterfield Reference Butterfield2000; Strother et al. Reference Strother, Battison, Brasier and Wellman2011), while most animal phyla (groups of organisms with a degree of evolutionary relatedness) did not appear as body fossils until the Cambrian Explosion 530 Myr ago. Due to their greater metabolic and environmental versatility and ability to survive – including in a dormant state – under extreme physical and chemical conditions, microbes are likely to be the only forms of life capable of surviving in the hostile environments that will be found on the far-future Earth. They are, by many criteria, the most diverse, abundant and successful forms of life on Earth, notable for not only surviving, but often thriving in what, from an anthropocentric view point, would be considered extreme environments (Cockell Reference Cockell2003). Hence, it is reasonable to assume that microbial life will be the most abundant form of life (if it is found) in habitable extraterrestrial environments. In fact, their ability to survive in extreme environments (such as the microbes living at temperatures of 90 °C in Grand Prismatic Spring in Yellowstone National Park, or those that live in the highly acidic waters of the Rio Tinto, Spain; Dartnell Reference Dartnell2011) opens up the range of extra-terrestrial environments that could be classified as habitable.
On Earth such organisms exhibit more modes of life beyond the mechanisms of oxygenic photosynthesis and aerobic respiration used by ‘higher’ plants (those that are most structurally divergent in differentiation among their cells) and animals (Oren Reference Oren2009). Thus, microbial life produces biosignatures that are unlike those produced by the majority of life on Earth as a whole. For example, Kaltenegger et al. (Reference Kaltenegger, Traub and Jucks2007) show that the early Earth (a predominantly microbial world) exhibited very different atmospheric biosignatures to the present Earth. Potentially, remotely detected biosignatures from an exo-Earth with just a microbial biosphere may not be immediately considered to be caused by life due to the dissimilarities to the signatures of life on the present Earth. This work is a first step towards simulating these more novel microbial environments under a diverse range of radiation regimes and surface environmental conditions to ascertain the likely remotely detectable biosignatures that would be produced.
The host planets considered in these simulations will often not be exact Earth analogues, and given galactic statistics, they will often be found around red dwarf stars (Tarter et al. Reference Tarter2007) and in binary star-systems (Welsh et al. Reference Welsh2012). Each host star system will provide a unique set of planetary environmental conditions from varying irradiation and stellar flare activity to unusual surface compositions and temperature ranges.
When developing any model of this nature, it is useful to calibrate it using a well-known system; hence, initially the Earth itself was used to test the model.
Microbial niches on the far-future earth
The Sun's main sequence evolution will result in very extreme environments for life on the far-future Earth (taken here to be 2–3 Gyr from the present). As the Sun ages, its luminosity increases, resulting in surface temperature increases on Earth. Increased temperatures cause increased atmospheric water vapour – a greenhouse gas, the presence of which further increases surface temperatures. However, this simplified overview neglects the role of cloud cover. Clouds can reduce planetary heating by reflecting solar energy away from the planet, increasing the planetary albedo, but can also act to trap radiation, increasing a heating effect (McGuffie & Henderson-Sellers Reference McGuffie and Henderson-Sellers2005). More water vapour entering the atmosphere could lead to increased cloud formation, but there are uncertainties regarding the effect of clouds and greenhouse warming (Goldblatt & Watson Reference Goldblatt and Watson2012) as a result of the number of dynamical and thermodynamical factors that would need to be considered (McGuffie & Henderson-Sellers Reference McGuffie and Henderson-Sellers2005). It is beyond the scope of this investigation to model this. Therefore, a constant cloud cover will be assumed in this case. Boer et al. (Reference Boer, Hamilton and Zhu2004) found that cloud cover variations were small until the solar constant increases by 25% of its present value; hence, this should be a reasonable assumption for most of the modelled time period.
Higher temperatures lead to increased weathering of silicate rocks, drawing down more carbon from the atmosphere. Carbon is normally recycled through plate tectonics; however, increasing water loss eventually halts plate tectonics due to greater friction between the plates. Lower atmospheric CO2 levels make embryophytic plant (liverworts, hornworts, mosses and vascular plants) life unsustainable as, unless a more effective photosynthetic carbon acquisition mechanism is used (Bar-Even et al. Reference Bar-Even, Noor, Lewis and Milo2010, Reference Bar-Even, Noor and Mil2012), plants require a minimum atmospheric CO2 concentration of approximately 10 ppm (Caldeira & Kasting Reference Caldeira and Kasting1992). Even with a higher-affinity carboxylase, photosynthesis in air would be restricted by diffusion in the gas phase through the diffusion boundary layer at very low bulk phase CO2 concentrations. Once levels drop below this value higher plants will begin to die off. This in turn decreases oxygen production, which, with continued consumption by biota and by oxidation of kerogen (organic carbon in sedimentary rocks), leads to a steady decline in atmospheric oxygen to zero over a few million years (Walker Reference Walker, Schneider and Boston1991). Since multicellular (metazoan) animal life depends on oxygen for respiration, the end of animal life would occur a few million years after the end of plant life (Walker Reference Walker, Schneider and Boston1991). Large endotherms (mammals and birds) would likely be the first group to become extinct due to their higher oxygen requirements compared with smaller endotherms and exotherms. As temperatures increase alongside declining atmospheric oxygen levels, eutherians (placental mammals) would be particularly vulnerable. Not only do these have higher oxygen requirements than non-placental mammals, which is said to have delayed the evolution of large placental mammals until atmospheric oxygen reached a certain level (Falkowski et al. Reference Falkowski, Katz, Milligan, Fennel, Cramer, Aubrey, Berner, Novacek and Zapol2005), but embryo development is very sensitive to excess heat (McLean Reference McLean, Kainlauri, Johansson, Kurki-Suonio and Geshwiler1991). Large herbivorous mammals would suffer from the decrease in food supplies as plant abundance decreases. Smaller mammals would have a slight stay of execution in comparison, due to their lower oxygen requirements and their larger surface-to-volume quotient, which aids in the dissipation of heat (McLean Reference McLean, Kainlauri, Johansson, Kurki-Suonio and Geshwiler1991). This scenario is slightly complicated by the fact that eutherians have higher body temperatures than marsupials and monotremes (Clarke & Rothery Reference Clarke and Rothery2008), which may, therefore, be more susceptible to increasing temperatures. Birds may be better suited to surviving than mammals larger than the largest birds as their generally smaller sizes mean they require less oxygen than larger animals, but also, migratory birds especially would be better able than similarly sized mammals to travel long distances to find lower temperature refugia; however, the number of these refugia for such animals would decline as temperatures continue to rise. These refugia would also tend to be at higher elevations where less land surface area is available, restricting population sizes as species migrate upwards (Sekercioglu et al. Reference Sekercioglu, Schneider, Fay and Loarie2007).
Ectothermic vertebrates (fish, amphibians and reptiles) would be able to survive for longer than endotherms in this scenario due to their better heat tolerances and, in general, lower oxygen requirements (some ectotherms have been observed to consume oxygen in greater quantities than living endotherms of the same body weight) (Kemp Reference Kemp2006). Decreasing water availability would make some amphibian species more vulnerable in such a world (Aráujo et al. Reference Aráujo, Thuiller and Pearson2006). Fish species would be similarly vulnerable. It should be noted that rapid ocean evaporation would not yet have begun at this time. Marine species may be able to survive for longer than freshwater species due to the orders of magnitude greater volume of ocean water than freshwater. For ectotherms, external temperature influences metabolic rate. An increased environmental temperature results in an increased metabolic rate and therefore, an increased need for food. Hence, surviving species may be vulnerable to starvation (Dillon et al. Reference Dillon, Wang and Huey2010). Reptile species with temperature-dependent sex determination would be more susceptible to increased temperatures (Aráujo et al. Reference Aráujo, Thuiller and Pearson2006).
Invertebrates could be the final animals present on Earth before all animal species are lost. Some insects, such as the spider beetles Mezium affine and Gibbium aequinoctiale have been observed to survive (but not necessarily to complete their life cycle) in temperatures of up to 56 °C (Yoder et al. Reference Yoder, Chambers, Tank and Keeney2009). In general, life on land will be more vulnerable than marine life initially as a result of the temperature-buffering effects of water. However, the loss of plant life on land would lead to a reduction in nutrients reaching the ocean. Communities isolated from marine food chains, such as volcanic vent communities, would likely survive for the longest (Ward & Brownlee Reference Ward and Brownlee2002). Within approximately 0.1 Gyr after the end of higher (embryophytic) plant life, Earth will once more become a microbial world.
Microbial photosynthesis could continue for a further 0.1 Gyr. Some eukaryotic phytoplankton and benthic macroalgae, and cyanobacteria, can grow and persist in environments with large CO2 fluctuations (Maberly Reference Maberly1996) and can carry out net photosynthesis at very low CO2 concentrations (Birmingham & Colman Reference Birmingham and Colman1979; Maberly Reference Maberly1990). Hence, CO2 concentrating mechanisms in cyanobacteria and, to a smaller extent, some eukaryotic algae could allow photosynthesis to take place down to CO2 concentrations of 1 ppm; 10 times less than the minimum for terrestrial higher (C4 photosynthesis) plant life (Caldeira & Kasting Reference Caldeira and Kasting1992). A steady decline in habitable environments with time leads to a pruning of the tree of life, concentrated towards extremophilic life adapted to multiple extreme conditions (polyextremophiles; Mesbah & Wiegel Reference Mesbah and Wiegel2012), such as thermohalophiles and chemoautotrophs. Finally, rising temperatures cause rapid ocean evaporation (Kasting Reference Kasting1988; Bounama et al. Reference Bounama, Franck and von Bloh2001; Vásquez et al. Reference Vásquez, Pallé and Montañés Rodríguez2010), effectively sterilising most of the biosphere (see Fig. 3 and Fig. 4).

Fig. 3. Simplified sequence of events leading to the loss of plant and animal life on a rapidly warming planet and the resulting microbial world.

Fig. 4. Simplified sequence of events leading from a microbial world with increasing temperatures and decreasing water availability to the end of all life on Earth.
It should be noted that it may be possible that the increase in atmospheric oxygen caused by the photodissociation of an ocean's worth of water vapour could cause the extinction of those final life forms that cannot tolerate high oxygen concentrations (similar to the extinction believed to have been caused by the Great Oxidation Event 2.4 Gyr ago) even before all the remaining pools of liquid surface water are lost. However, this future oxygen atmosphere would only persist for a geologically short period of time before oxygen is removed via oxidation, and surface reactions. Some oxygenic photosynthetic organisms are able to survive and grow at oxygen equilibrium gas phase partial pressures of 80 kPa in solution (at sea level) (Raven & Larkum Reference Raven and Larkum2007); therefore, organisms with similar tolerances would be better suited to surviving a brief high-oxygen period.
With the complete cessation of photosynthesis, but the continuation of all other biological activity at the present-day rate, the half-life of atmospheric oxygen would be approximately 3 Myr (Walker Reference Walker, Schneider and Boston1991); hence, the geologically short time-span for the continuation of animal life after higher plant life comes to an end.
On the far-future Earth, the only likely oxygen sink would be nitrogen fixation by lightning. At present there is approximately 8 × 1019 moles of atmospheric oxygen. With 5 × 1011 moles of nitrogen converted to nitric acid by lightning each year (Tie et al. Reference Tie, Zhang, Brasseur and Lei2002), atmospheric oxygen would have a half-life of approximately 100 Myr if fixation by lightning were the only oxygen removal process. However, lightning frequency is directly linked to a planet's climate, with the frequency of lightning strikes increasing with increased surface temperature (Williams et al. Reference Williams, Mushtak, Rosenfeld, Goodman and Boccippio2005; Price & Asfur Reference Price and Asfur2006a, Sekiguchi et al. Reference Sekiguchi, Hayakawa, Nickolaenko and Hobara2006). Lightning frequency has also been observed to increase with increased cloud cover in the tropics (Sato & Fukunishi Reference Sato and Fukunishi2005). Lightning modelling studies suggest that global lightning activity increases by 10% for every 1 K increase in global temperature (Price Reference Price, Betz, Schumann and Laroche2009). Hence, the rate of atmospheric oxygen removal on a warmer, far-future Earth would be much higher than present rates. For example, a global temperature 100 K higher than that at present would result in approximately 6 × 1015 moles of nitrogen converted into nitric acid per year. Even with a higher level of atmospheric oxygen than the present level (the ocean contains approximately 7 × 1022 moles of oxygen), the half-life of atmospheric oxygen would be of the order 6 Myr, suggesting a geologically short oxygen-rich period during the rapid photodissociation period. However, the question of whether this would be a planet-sterilizing event remains open.
As the climate evolves alongside solar main sequence evolution, life on Earth and its associated biosignatures will also evolve until the point at which the planet becomes uninhabitable. This raises the question of when biological activity on Earth would be reduced to such an extent that the planet no longer exhibits remotely detectable biosignatures.
Assuming that remotely detectable life resides on, or near the surface (signatures of deep biospheres tend to require direct sampling of the biosphere itself; Parnell et al. Reference Parnell, Boyce and Blamey2010) and that such life requires liquid water (Rothschild & Mancinelli Reference Rothschild and Mancinelli2001), a step towards answering this question was taken by constructing a model to find the most likely regions on the far-future Earth for the planet's ‘last’ life.
While solar evolution will ultimately be responsible for the expiration of the biosphere, there are many other factors that will affect the climate evolution on Earth along the way. The orbital parameters of the planet, specifically its eccentricity and obliquity, are known to vary over time, resulting in climate changes as they do so (Spiegel et al. Reference Spiegel, Raymond, Dressing, Scharf and Mitchell2010). Atmospheric composition also has a role to play. Changing concentrations of greenhouse gases in particular have a notable effect on surface temperatures (Kasting & Grinspoon Reference Kasting and Grinspoon1991; Pavlov et al. Reference Pavlov, Kasting, Brown, Rages and Freedman2000). Climate changes themselves can then alter other climate-influencing factors such as planetary albedo (Rosing et al. Reference Rosing, Bird, Sleep and Bjerrum2010). These factors were taken into account alongside an evolving Sun to create a toy latitude-dependent radiative balance model for Earth to better constrain where the last habitable areas on the planet may be.
Methods – factors influencing habitability
Solar luminosity
The primary driver for habitability change is the increase in solar luminosity over time. This can be accounted for by following the convention of Gough (Reference Gough1981) in which the luminosity at any time during the Sun's main sequence lifetime L(t) is given by:
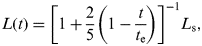 $$L(t) = \left[ {1 + \displaystyle{2 \over 5}\left( {1 - \displaystyle{t \over {t_{\rm e}}}} \right)} \right]^{ - 1} L_{\rm s}, $$
$$L(t) = \left[ {1 + \displaystyle{2 \over 5}\left( {1 - \displaystyle{t \over {t_{\rm e}}}} \right)} \right]^{ - 1} L_{\rm s}, $$where L s is the present solar luminosity, t e is the current age of the Sun and t is the time elapsed on the main sequence. The surface insolation depends on the solar constant S 0, which is influenced by solar luminosity such that
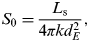 $$S_0 = \displaystyle{{L_{\rm s}} \over {4\pi kd_E^2}}, $$
$$S_0 = \displaystyle{{L_{\rm s}} \over {4\pi kd_E^2}}, $$where k is a constant (k≈1) and d E2 is the Earth–Sun distance. By incorporating a varying luminosity and assuming a total insolation S T = S 0/4 (as the surface of a sphere has an area four times that of a flat disc with the same radius), the effect of increased luminosity on mean global temperatures can be modelled.
The time evolution of this mean temperature can be calculated from the difference between incoming and outgoing radiation, i.e.
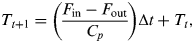 $$T_{t + 1} = \left( {\displaystyle{{F_{{\rm in}} - F_{{\rm out}}} \over {C_p}}} \right){\rm \Delta} t + T_t, $$
$$T_{t + 1} = \left( {\displaystyle{{F_{{\rm in}} - F_{{\rm out}}} \over {C_p}}} \right){\rm \Delta} t + T_t, $$where F in and F out are the incoming and outgoing radiation fluxes and C p is the heat capacity of the planet (at constant pressure). In this case, the incoming radiation is assumed to be a function of the solar insolation
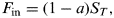 $$F_{{\rm in}} = (1 - a)S_T, $$
$$F_{{\rm in}} = (1 - a)S_T, $$where a is the temperature-dependent albedo and the outgoing radiation is given by
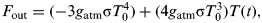 $$F_{{\rm out}} = ( - 3g_{{\rm atm}} {\rm \sigma} T_0^4 ) + (4g_{{\rm atm}} {\rm \sigma} T_0^3 )T(t),$$
$$F_{{\rm out}} = ( - 3g_{{\rm atm}} {\rm \sigma} T_0^4 ) + (4g_{{\rm atm}} {\rm \sigma} T_0^3 )T(t),$$where g atm is a variable factor accounting for the transmissivity of the atmosphere (0⩽g atm ⩽ 1), σ is the Stefan–Boltzmann constant and T 0 is the initial temperature of the system. Heat is then allowed to diffuse out from the equator to higher latitudes as described in Lorenz et al. (Reference Lorenz, Lunine, Withers and McKay2001), i.e. representing the convective heat flow from the tropics (a hot reservoir) to mid-latitudes and the poles (cold reservoirs). The magnitude of this heat flow is controlled by the latitudinal diffusion coefficient, which increases as temperatures rise and cause an increase in atmospheric density (Lorenz et al. Reference Lorenz, Lunine, Withers and McKay2001).
Greenhouse gases
The levels of greenhouse gases in the atmosphere affect the outgoing radiation lost to space and, therefore a planet's surface temperature. Atmospheric gases such as CO2, water vapour, methane, etc. absorb and emit radiation at infrared wavelengths contributing to a global greenhouse effect. The change in longwave radiation forcing due to CO2 and water vapour is proportional to the logarithm of their concentration in the atmosphere (Myhre et al. Reference Myhre, Highwood, Shine and Stordal1998; Rákóczi & Iványi Reference Rákóczi and Iványi1999; Allan Reference Allan2012).
In this model, particular attention is paid to the levels of water vapour in the atmosphere (a result of rapid ocean evaporation caused by a runaway greenhouse effect) and the effects of carbon draw-down (the removal of atmospheric carbon as a result of the weathering of silicate rocks).
This is included in the model by taking account of the partial optical thickness of the atmosphere due to both CO2 and H2O. This gives a total long-wave optical depth (τ) made up of the partial optical depths of CO2 and H2O of
 $${\rm \tau} = k_{{\rm CO}_{\rm 2}} P_{{\rm CO}_2} ^{0.53} + k_{{\rm H}_{\rm 2} {\rm O}} P_{{\rm H}_{\rm 2} {\rm O}}^{{\rm 0}{\rm. 3}} $$
$${\rm \tau} = k_{{\rm CO}_{\rm 2}} P_{{\rm CO}_2} ^{0.53} + k_{{\rm H}_{\rm 2} {\rm O}} P_{{\rm H}_{\rm 2} {\rm O}}^{{\rm 0}{\rm. 3}} $$(Levenson Reference Levenson2011), where k i is a proportionality constant and P i is the partial pressure, which evolves as the number density (n i) evolves with time such that
 $$P_i = \displaystyle{{n_i RT} \over {N_A}}, $$
$$P_i = \displaystyle{{n_i RT} \over {N_A}}, $$where R is the universal gas constant and N A is Avogadro's constant.
Orbital characteristics
Earth's position and orientation relative to the Sun are not static, but vary sufficiently over time to impact the planet's climate. In particular, eccentricity, precession and obliquity changes (the Milankovitch Cycles) are known to have a major impact on climate over geological time-spans (Spiegel et al. Reference Spiegel, Raymond, Dressing, Scharf and Mitchell2010). Precession (the rotation of the Earth's axial tilt) occurs on a cycle of approximately 19 000 years, obliquity varies between 22.1ο and 24.5ο approximately every 41 000 years and eccentricity cycles between 0 and 0.06 approximately every 100 000 years (Paillard Reference Paillard2010; Vásquez et al. Reference Vásquez, Pallé and Montañés Rodríguez2010).
The influence of obliquity variations on the insolation received at a given latitude can be found from
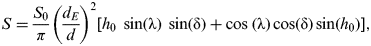 $$S = \displaystyle{{S_0} \over \pi} \left( {\displaystyle{{d_E} \over d}} \right)^2 \left[ {h_0 \;\sin ({\rm \lambda} )\;\sin ({\rm \delta} ) + \cos \;({\rm \lambda} )\cos ({\rm \delta} )\sin (h_0 )} \right],$$
$$S = \displaystyle{{S_0} \over \pi} \left( {\displaystyle{{d_E} \over d}} \right)^2 \left[ {h_0 \;\sin ({\rm \lambda} )\;\sin ({\rm \delta} ) + \cos \;({\rm \lambda} )\cos ({\rm \delta} )\sin (h_0 )} \right],$$where d is the Earth–Sun distance at a given point in the planet's orbit, h 0 is the solar hour angle at which insolation at a particular latitude becomes positive, λ is the latitude and δ is the solar declination, which depends on obliquity (ϕ) such that
 $$\sin ({\rm \delta} ) = \sin ({\rm \varphi} )\sin ({\rm \theta} - {\rm \omega} ),$$
$$\sin ({\rm \delta} ) = \sin ({\rm \varphi} )\sin ({\rm \theta} - {\rm \omega} ),$$where θ is the orbital longitude of the planet and ω is the longitude of perihelion. The eccentricity (e) of the planet's orbit influences the planet–star distance (d) at any point in the orbit following the relation:
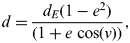 $$d = \displaystyle{{d_E (1 - e^2 )} \over {(1 + e\,\cos (\nu ))}},$$
$$d = \displaystyle{{d_E (1 - e^2 )} \over {(1 + e\,\cos (\nu ))}},$$where ν is the true anomaly of the orbit, i.e. the angle formed by the line joining a planet and star and the central axis of the ellipse formed by the planet's orbit.
While they provide a guide as to the likely behaviour of the planet's orbital characteristics from the present onwards, the Milankovitch cycles are unlikely to remain constant for the entire lifetime of the planet. The presence of a large moon helps to stabilize Earth's obliquity, preventing large obliquity swings like those experienced by Mars for example (Laskar et al. Reference Laskar, Joutel and Robutel1993, Reference Laskar, Correia, Gastineau, Joutel, Levrard and Robutel2004). While the presence of a large moon may not be essential for a stable obliquity range for Earth-like planets (Lissauer et al. Reference Lissauer, Barnes and Chambers2012), the recession of an already present large moon could potentially induce larger obliquity swings, effecting latitudinal temperature values. Earth's moon is receding due to tidal interactions with Earth at a current rate of approximately 4 cm/yr (Néron de Surgy & Laskar Reference Néron de Surgy and Laskar1997), placing it 40 000 km further away within 1 Gyr (if this recession rate remains constant). This gives it an orbital distance of 67R ⊕ (R ⊕ = Earth radii), exceeding the critical point described by Tomasella et al. (Reference Tomasella, Marzari and Vanzani1996) for stabilizing planetary obliquity. Exceeding this distance places the Earth into a chaotic obliquity regime permitting large obliquity swings between 30ο and 60ο (Tomasella et al. Reference Tomasella, Marzari and Vanzani1996) and possibly allowing even larger obliquities of up to 90ο (Laskar et al. Reference Laskar, Joutel and Robutel1993). This would effect global temperature distribution, causing shifts in the latitude zones that receive maximum and minimum insolation. As the Moon recedes, Earth's rotation rate will slow. This reduces the equator-to-pole temperature gradient by reducing the magnitude of the mid-latitude eddies responsible for heat transport (Feulner Reference Feulner2012), again influencing the global temperature distribution.
Similarly, eccentricity will not remain stable over geological timescales. Laskar & Gastineau (Reference Laskar and Gastineau2009) suggest that a resonance between Mercury and Jupiter may eventually increase Mercury's eccentricity, which would destabilise the inner planets in approximately 3 Gyr – a scenario that could lead to Earth's eccentricity reaching a value as high as 0.3. However, this change is likely to come too late to influence Earth's final life.
Therefore, the evolution of orbital characteristics will influence climate evolution and hence, the evolution of planetary habitability. Thus, these factors need to be included in a habitability evolution model.
Hydrogen escape and the runaway greenhouse effect
Surface temperatures and incoming radiation can drive positive feedback cycles that eventually send a planet's climate out of radiative equilibrium, potentially causing abrupt environmental changes. In particular, runaway and moist greenhouse effects can put an end to a planet's habitable life.
As the luminosity of a star increases, the radiation intercepted by an orbiting planet increases. This increases evaporation rates of surface water, raising the atmospheric water vapour content. Water vapour being a greenhouse gas, this results in further temperature increase, beginning a positive feedback loop (McGuffie & Henderson-Sellers Reference McGuffie and Henderson-Sellers2005; Goldblatt & Watson Reference Goldblatt and Watson2012). A moist greenhouse state describes the state in which water vapour dominates the troposphere while the water vapour content of the stratosphere starts to increase. A runaway greenhouse scenario occurs when water vapour becomes a dominant component of the atmosphere. At this point, the moist adiabatic lapse rate (the net rate of temperature increase with height for an atmosphere saturated with water vapour) tends towards the saturation vapour pressure curve for water (i.e. the pressure at which water vapour becomes saturated for a given temperature) resulting in a fixed temperature-pressure structure for the atmosphere (Barnes et al. Reference Barnes, Mullins, Goldblatt, Meadows, Kasting and Heller2012; Goldblatt & Watson Reference Goldblatt and Watson2012).
A fixed temperature-pressure structure means that the energy radiated back into space by the planet is also fixed. If the incoming radiation exceeds this limit, there is a net gain of energy by the planet and, provided that water vapour is not lost from the atmosphere, runaway heating occurs, eventually resulting in the evaporation of surface oceans. The oceans are finally lost to space through hydrogen escape. Ordinarily, on Earth, little water vapour reaches the stratosphere as the current temperature-pressure structure leads to water condensing at lower altitudes. However, during a runaway greenhouse, the entire atmosphere becomes water saturated, causing photo-dissociation of water molecules in the upper atmosphere to contribute significantly to the hydrogen escape.
In this case, hydrogen loss to space would be approximately 6.6 × 1011 atoms per cm2 per second, limited only by the solar extreme UV heating rate (Caldeira & Kasting Reference Caldeira and Kasting1992), the intensity of which should be reasonably similar to the present flux for an ageing sun-like star (see, for example Ribas et al. Reference Ribas, Guinan, Gúdel and Audard2005). Hence, assuming Hydrogen loss is uniform across the surface area of the outer atmosphere (assuming the surface area of a sphere of radius 1R ⊕), 3.06 × 1030 hydrogen atoms would be lost per second. There are approximately 4.4 × 1046 water molecules in the ocean (8.8 × 1046 H atoms), therefore all water would be lost to space in approximately 1 Gyr.
Results and discussion
Figure 5 shows the general mean surface temperature trends predicted by the model for the next 2.5 Gyr. The steep increase in temperature at around 1 Gyr from the present represents the onset of rapid ocean evaporation. Assuming an upper temperature bound for life of 420 K (allowing some increase over the currently known upper temperature tolerance of thermophiles along the lines of Ward & Brownlee (Reference Ward and Brownlee2002)) and assuming no changes to obliquity or eccentricity cycles, life could persist 0.7 Gyr longer at surface levels at the poles than at the equator.

Fig. 5. Change in global mean temperatures over time with increasing solar luminosity. The dashed line represents equatorial temperature and the solid line represents polar temperature. After about 1 Gyr a moist greenhouse begins when temperatures reach 330 K allowing the water vapour content of the stratosphere to increase rapidly. When temperatures reach approximately 420 K life would likely no longer be able to survive. A runaway greenhouse regime begins after approximately 2.8 Gyr. Initially, the poles warm noticeably less rapidly than the equator; however, as the planet heats up, the equator-to-pole temperature gradient decreases due to an increased latitudinal heat diffusion coefficient, caused by the increase in atmospheric pressure.
Rapid ocean loss as a result of a moist greenhouse effect would likely represent the end-point of a planet's habitable lifetime. Assuming ocean loss was not uniform across the globe due to regional temperature variations, there could potentially be pockets of liquid water that remain for a brief time before total loss of liquid surface water occurs. A source of liquid water is a prerequisite for life as we know it; hence, these last pools of water would represent the final habitable regions on a dying planet. In this section, potential locations for these last habitable regions are discussed.
Ocean floor trenches
The present mean ocean depth is approximately 4 km; however, deep ocean trenches, depressions in the sea floor caused by the subduction of one tectonic plate under another at divergent plate boundaries can extend the ocean depth to 6 km on average and up to 11 km for the deepest known trench, the Mariana Trench in the Western Pacific Ocean.
One could imagine that as surface water is lost these would provide sheltered regions, cooler than the ambient temperatures outside the trench, much like the shadowing effect of craters on the Moon can reduce the temperatures within them by as much as 295 K compared to surface temperatures (Margot et al. Reference Margot, Campbell, Jurgens and Slade1999). However, the presence of an atmosphere on Earth complicates the situation. Movement of air into a trench from above would cause that air to be compressed the lower it goes. This compression increases the air temperature, which could lead to trenches actually being some of the warmest regions on a runaway greenhouse planet and thus, making them unlikely candidates for the location of some of the last liquid surface water on the planet.
Additionally, it is possible that trenches may not even be present 1 Gyr from now. Plate tectonics is driven by the transfer of heat from the Earth's core to the surface. As described in more detail in Meadows (Reference Meadows2007), heat is transferred from the core through the mantle via convection (and sometimes conduction). As these plumes of heat reach the crust they give up their heat and are pushed aside by new hot uprisings. This sideways motion in the upper mantle is the driving force for plate movement. However, the Earth is cooling down over time, resulting in a gradually solidifying outer liquid core. This slows down convection within the mantle, leaving less power available to move plates. Eventually (approximately 3 Gyr from now), convection will stop altogether, ending plate motions.
This slowing of tectonic processes may mean that there may not be any deep ocean trench features on the far future Earth. Additionally, regardless of whether convection is still occurring in the mantle, once the oceans boil away, tectonic plate movements will stop. The subduction of one plate under another is aided by the presence of liquid surface water, which acts as a lubricant. Without it, plate movements cease due to increased friction (Meadows Reference Meadows2007).
High altitude pools
Another potential refuge could be high altitude lakes. Currently, temperatures in the troposphere follow a mean linear decrease of ∼6.5 K per km with increasing altitude (an average of the lapse rates calculated for dry and moist air) due to the fact that solar radiation is absorbed by the planet's surface, which re-radiates it, heating the lower atmosphere. Assuming this negative trend still holds on the far future Earth, life could be expected to migrate upwards to more comfortable, liquid water permissible temperatures as surface temperatures rise.
An estimate of the likely value of the (adiabatic) lapse rate Γw during the planet's moist greenhouse phase can be obtained from
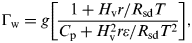 $$\Gamma _{\rm w} = g\left[ {\displaystyle{{1 + H_{\rm v} r/R_{{\rm sd}} T} \over {C_{\rm p} + H_{\rm v}^{\rm 2} r\varepsilon /R_{{\rm sd}} T^2}}} \right],$$
$$\Gamma _{\rm w} = g\left[ {\displaystyle{{1 + H_{\rm v} r/R_{{\rm sd}} T} \over {C_{\rm p} + H_{\rm v}^{\rm 2} r\varepsilon /R_{{\rm sd}} T^2}}} \right],$$where g is the gravitational acceleration, H v is the heat of vaporization for water, R sd is the specific gas constant for dry air, ε is the ratio of the specific gas constant of dry air to that for water, T is the surface temperature and r is the ratio of the mass of water vapour to dry air, which depends on the saturated vapour pressure and atmospheric pressure.
At an altitude of 10 km (with no changes to obliquity or eccentricity cycles) life could persist for approximately 0.7 Gyr longer at polar latitudes than in equatorial regions (assuming an upper temperature tolerance of 420 K) – see Figure 6. Polar surface life could outlast equatorial high-altitude life by approximately 0.2 Gyr.

Fig. 6. Mean temperature evolution at the equator (black) and the poles (grey) with increasing altitude and estimated tropospheric lapse rate from surface level (solid line) to an altitude of 10 km (dashed line).
The case for high altitude pools may be complicated by the slowing, or halting of plate tectonics. This would mean that, except in regions where magma plumes continue to rise to the surface, continental uplift could slow down or stop, allowing the weathering rate to exceed the mountain-building rate, potentially resulting in less high altitude land area (Meadows Reference Meadows2007). However, the lack of plate movements could allow volcanoes surrounding magma plumes (hot spot volcanoes) to grow taller than those on the present Earth as material is able to accumulate in one location for a longer period of time – the driver behind very tall martian volcanoes, such as Olympus Mons, which stands at a height of 21 km (Wood Reference Wood1984).
Additionally, an airborne biosphere of micro-organisms could also take advantage of the cooler temperatures at higher altitudes. Micro-organisms are known to currently be present in the atmosphere (Bowers et al. Reference Bowers, Lauber, Wiedinmyer, Hamady, Hallar, Fall, Knight and Fierer2009; Womack et al. Reference Womack, Bohannan and Green2010), some of which remain metabolically active and have residence times in the atmosphere that are long enough for them to reproduce (Womack et al. Reference Womack, Bohannan and Green2010).
Caves
It can be argued that life on the far future Earth would retreat underground and into cave systems (the subsurface biosphere) (Irwin & Schulze-Makuch Reference Irwin and Schulze-Makuch2011). Many microbial communities are already known to exist independently of solar energy, for example, obtaining the means to metabolize directly from the rocks (chemolithotrophs) and in the case of one strain of green sulphur bacteria, even using the extreme short-wave tail of the geothermal black body radiation (at not more than 600 K) around a deep sea vent for photosynthesis (Beatty et al. Reference Beatty, Oevrmann, Lince, Manske, Lang, Blankenship, Van Dover, Martinson and Plumley2005).
Caves are generally assumed to have constant interior environments, with internal temperatures closely approximating the local mean surface temperature (Tuttle & Stevenson Reference Tuttle and Stevenson1977; Howarth Reference Howarth1983). This, combined with the general trend of increasing temperatures with depth would not bode well for organisms on a planet with surface temperatures exceeding the boiling point of water. However, some cave systems may be better suited to sheltering life than others. In particular, caves that have their greatest volume below their entrance would act as cold reservoirs (Tuttle & Stevenson Reference Tuttle and Stevenson1977) as colder, denser air would flow downward into the cave, but warmer lighter air would not.
These cold trap caves (also known as ice caves due to the presence of year-round ice within them) are often formed from collapsed lava tubes (Williams et al. Reference Williams, McKay, Toon and Head2010) and generally have a narrow single entrance with a large chamber below, which has a large volume-to-wall surface ratio, because it is conduction through the cave walls that would restore the temperature within the cave to the mean surface temperature.
Cold, dense air enters the cave during winter. In-falling snow is compacted into layers of ice, or inflowing water freezes. When temperatures outside increase, the warmer, less dense air cannot enter the cave, leaving the colder air trapped within. This allows ice to remain within the cave throughout the year. However, the ice mass within the cave is not the ice that originally formed there. The ice is continually being melted as a result of heat conduction from the surrounding rocks that are in contact with the ice (which tend to reflect the annual mean temperature of a region) and dripping water inside the cave (Ohata et al. Reference Ohata, Furukawa and Osada1994; Leutscher et al. Reference Leutscher, Jeannin and Haeberli2005). The melt water is then lost from the cave with the entire ice mass being replaced after between 100–1000 years (Leutscher et al. Reference Leutscher, Jeannin and Haeberli2005), depending on the geometry of the particular cave in question. Hence, new water needs to be supplied (Δw) such that
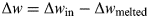 $${\rm \Delta} w = {\rm \Delta} w_{\rm in} - {\rm \Delta} w_{{\rm melted}} $$
$${\rm \Delta} w = {\rm \Delta} w_{\rm in} - {\rm \Delta} w_{{\rm melted}} $$remains constant for permanent ice to remain.
Therefore, for this mechanism to provide a useful refuge for life on the far-future Earth, there would need to be some form of water input into the system (perhaps through seepage from the surrounding rock or via water vapour entering from the external atmosphere at times when the outside air density matches, or exceeds that within the cave) for liquid water to remain within such a cave.
Ice caves are found not only in cold regions but also in temperate and tropical latitudes on Earth today. The question of how long such caves would be able to retain water when exposed to rapidly increasing surface temperatures remains open. Leutscher et al. (Reference Leutscher, Jeannin and Haeberli2005) focused on ice caves in the Jura Mountains and found that the temperatures within cold trap caves were dependent on the outside winter temperatures in the region and, more particularly relevant to this work, that the ice mass within the ice caves studied decreased as annual mean winter temperatures increased (most noticeably since the 1980s). This suggests that cold trap cave temperatures on the far-future Earth would only be as (relatively) cool as the coolest temperatures in a region in any given year, which, given the high temperatures expected, may not provide a long stay of execution for life.
However, for Earth, after the Moon recedes beyond the critical distance of 67R ⊕ described by Tomasella et al. (Reference Tomasella, Marzari and Vanzani1996) (approximately 1 Gyr from now), Earth's obliquity enters a regime in which it can vary chaotically between much higher values than the present obliquity range. If obliquity cycling in the model is altered to vary between 30ο and 60ο (and even up to 90ο), the equator-to-pole temperature gradient is effectively reversed by the new, higher obliquity range, which causes the poles to receive more insolation than the equator (in agreement with Tomasella et al. (Reference Tomasella, Marzari and Vanzani1996)). As such high obliquity planets do not necessarily preclude habitability (Williams & Kasting, Reference Tomasella, Marzari and Vanzani1997) this suggests that equatorial regions may be more accommodating to life than polar regions on the far-future Earth.
These large obliquity swings raise the possibility that the cold trap cave mechanism could extend the lifetime of liquid water habitats further than expected. The more extreme temperature differences between seasons that result from increased obliquity would cause cooler winter temperatures. For example, as illustrated in Figure 7, an obliquity of 60ο would allow winter temperatures that are up to 40ο lower than summer temperatures in some regions, whereas an obliquity of 90ο could potentially allow very extreme seasonal temperature variations of more than 200ο. This, of course, neglects latitudinal heat transport, which acts to lower the temperature range (especially as atmospheric density increases). By including simple heat transport, an estimate of the magnitude of the lower temperature values over latitude for different obliquity values was found (Figure 8). A high obliquity far-future Earth with a cold-trap cave could extend the stay of execution for life as far as +2.8 Gyr from present.

Fig. 7. Seasonal temperature variations between equatorial and polar latitudes for obliquities of 60 and 90ο for a solar luminosity of 1.1 Ls. The solid lines represent the upper temperature range and the dashed lines represent the lower-temperature range.

Fig. 8. Change in lower-temperature range over latitude for obliquities of 30, 60 and 90° at 2.0, 2.5, 2.8 and 3.0 Gyr from present. These suggest that, for high obliquities, temperatures may permit liquid water at some or all latitudes at certain points in the planet's orbit for at least 2.8 Gyr from present, which would allow for a cold trap cave mechanism to harbour liquid water year-round in these cases. While the +3 Gyr example shows a slight dip below the boiling point of water for high/low latitudes for obliquities of 60/90°, the onset of a runaway greenhouse would lead to much less predictable climate dynamics and water availability would be very limited.
The deep subsurface
The deep-subsurface environment would likely provide more stable refuges for life. It has been claimed that the deep biosphere may contain the vast majority of Earth's microbial biomass, with 50–80% of this estimated to occur in the deep marine subsurface (Orcutt et al. Reference Orcutt, Sylvan, Knab and Edwards2011). It is officially defined as depths of 50 m or more below the continental surface or the ocean floor (Bell Reference Bell2012) although, in practice, deep biosphere research can encompass any depths below 1 m (Bell Reference Bell2012; Edwards et al. Reference Edwards, Becker and Colwell2012). The lower depth limit is currently unknown (Kieft et al. Reference Kieft2005; Reith Reference Reith2011). The limiting factor is likely to be temperature. It is known that temperature increases with depth, with the mean geothermal temperature gradient for Earth's crust being ∼30ο per km depth (Bohlen Reference Bohlen1987). This would place the lower limit at the depth where temperatures reach the upper-limit for life (122 °C) – approximately 3.5 km for the present Earth.
However, the geothermal gradient varies depending on rock type (Pedersen Reference Pedersen2000), increasing or decreasing the maximum habitable depth in a given region. The deepest microbial life found to date was found in igneous rock aquifers at depths of 5.3 km. Therefore, although assuming a mean geothermal gradient of 30ο would lead to predicted temperatures that preclude habitability once surface temperatures reach the maximum for life, the variable nature of the crustal temperature gradient could allow the deep subsurface biosphere to persist, perhaps for longer than any other refuge.
In the deep subsurface biosphere life has been characterized that operates independently of surface photosynthesis (Kieft et al. Reference Kieft2005; Lin et al. Reference Lin, Slater, Lollar, Lacrampe-Couloume and Onstott2005, Reference Lin2006; Chivian et al. Reference Chivian2008). From a remote detection perspective it may be more challenging to detect the presence of life in these environments; however, life existing independently of photosynthesis might produce methane, which could be detectable (Parnell et al. Reference Parnell, Boyce and Blamey2010).
Future life in hot, saline environments?
Wherever these last pools of water may be found, they are likely to share some common characteristics. They are likely to be warm, isolated and (at least for ocean remnants) highly saline. This suggests that the last life forms would be those that can tolerate, or require high salt concentrations and temperatures (thermohalophiles).
Life on the far-future Earth will likely be found in microbial mat communities resembling some present day hydrothermal vent communities (Ward & Brownlee Reference Ward and Brownlee2002). High temperature, high salinity hydrothermal vent environments are found on Earth today. For example, the shallow-water Milos submarine hydrothermal field in the central Hellenic Volcanic Arc provides such an environment. The high salinity of some waters in this region (as much as 2.5 times higher than ordinary sea water; Valsami-Jones et al. Reference Valsami-Jones, Baltatzis, Bailey, Boyce, Alexander, Magganas, Anderson, Waldron and Ragnarsdottir2005) means that only microbial mats can thrive in those areas immediately surrounding hydrothermal vents (Thiermann et al. Reference Thiermann, Akoumianaki, Hughes and Giere1997).
It is not possible to say with any certainty what the likely pH of the final pools of liquid water would be. Alkaliphilic and moderately acid-tolerant forms of halophilic micro-organisms are known on Earth today, spanning an optimal pH range from 4 to 10 (DasSarma & DasSarma Reference DasSarma and DasSarma2001), whereas thermophiles are found across the pH range from 1 to 10 (Ferrera & Reysenbach Reference Ferrera and Reysenbach2007). Therefore, although the pH of these far-future environments cannot be determined, the required survival mechanisms for high temperature, high salinity environments should allow organisms to adapt to these environments.
Organisms living in this future epoch would also be subjected to elevated UV radiation levels due to the low abundance of atmospheric ozone. Ozone is produced from O2 and when atmospheric O2 decreases, as a result of the lack of O2 production when temperatures exceed the upper limit for oxygenic photosynthesis, the steady state level of ozone decreases, resulting in less attenuation of biologically damaging UV radiation below 290–300 nm (Beaty et al. Reference Beaty2006).
Recently characterized microbiological communities from some of the highest Atacaman volcanoes (Lynch et al. Reference Lynch, King, Farías, Sowell, Vitry and Schmidt2012) may reflect the style of microbial communities expected on a hot, dry Earth. These exist under a very intense solar radiation regime, with large (ΔT>60 °C) daily temperature variations and under very arid conditions. Known examples of microbes adapted to high salinities and warm temperatures are Halothermothrix orenii, Thermohalobacter berrensis and Bacillus aeolius: these provide plausible models for the last life on Earth. Halobacteria and many other salt-dwelling Archaea and Bacteria often inhabit salt crusts, which confer protection from deleterious UV radiation (Landis Reference Landis2001) and one could imagine salt-encrusted communities of thermohalophiles inhabiting the last remaining bodies of water. The supposition that such life would be halophilic depends, of course, on the chemical composition of the water.
Future biosignatures?
This leads to the question of what the dominant by-products of such an extremophile biosphere would be and whether this would allow for any remotely detectable atmospheric biosignatures. The micro-organisms from the Atacaman volcanoes appear to use the oxidation of carbon monoxide to obtain energy (Lynch et al. Reference Lynch, King, Farías, Sowell, Vitry and Schmidt2012). While the slowing of plate tectonics may decrease the amount of available CO, hotspot volcanoes (those caused by upwellings from the deep mantle rather than at plate boundaries) could still exist. Mars (which is not tectonically active) exhibited this kind of volcanism up to as early as 2 Myr ago (Neukum et al. Reference Neukum2004), while hotspot volcanism may still be actively ongoing on Venus (Smrekar et al. Reference Smrekar, Stofan, Mueller, Treiman, Elkins-Tanton, Helbert, Piccioni and Drossart2010). The thermohalophilic organisms listed above are all anaerobic organisms which produce hydrogen, CO2, ethanol and acetate as by-products of their metabolic processes. Acetates are not promising as remotely detectable biosignatures, only managing to escape into the atmosphere (at a very slow rate) if they are formed within very acidic (pH > 4.7) water (Seager et al. Reference Seager, Schrenk and Bains2012).
On a planet with relatively low CO2 levels compared to the present Earth, it may be possible that the biological production of CO2 would produce a disequilibrium of the gas in the atmosphere that would indicate the presence of life, especially if it can easily be deduced that the planet is likely to be near the end of its habitable lifetime. Similarly, if enough hydrogen is produced biologically, there could be a noticeable excess of atmospheric hydrogen, if biological hydrogen production is not comparatively negligible to that produced abiotically.
Global biological productivity on the early (pre-photosynthetic) Earth was much lower than that present-day value, as illustrated by the values in Table 1. Similarly, it is likely that productivity on the far-future Earth will be much lower than at present. An estimate for the likely productivity of the planet towards the end of its habitable lifetime is obtained by looking at the productivity values for environments similar to the proposed refugia for life (summarized in Table 2).
Table 1. Net global biological productivity values for modern and early (pre-photosynthetic) Earth

Table 2. Biological productivity values for environments similar to the proposed refugia on the far-future Earth

As a best-case scenario, assuming a global distribution of geothermal pools equal in size to the largest on the present-day Earth (Frying Pan Lake, New Zealand – 200 000 m3; Scott Reference Scott1994), a distribution of ice caves similar to that in the Jura Mountains (40 caves; Leutscher et al. Reference Leutscher, Jeannin and Haeberli2005) for 10 mountain ranges, a similar number of active hydrothermal vent fields (there are estimated to be approximately 1000 currently active vent fields; Baker & German Reference Baker and German2004) and a microbially inhabited basaltic crust, a guideline figure for global net productivity of 0.23 (×1012 moles of C yr−1) is obtained for the far-future Earth; two orders of magnitude lower than the pre-photosynthetic Earth. This suggests that the biosignatures produced by the far-future biosphere would be less intense than those on a world with higher global net biological productivity.
Conclusions
As the luminosity of the Sun increases during its main sequence lifetime, surface temperatures on Earth will increase, leading to complete evaporation of the Earth's oceans and an end to the Earth's habitable lifetime. As the planet nears the end of its habitable lifetime, it will likely only support microbial life; more specifically, life adapted to survive in harsh high-temperature, high-salinity environments. A maximum lifetime for life on Earth of 2.8 Gyr from present was found, given the presence of sheltered, high-altitude or high-latitude environments. These niches should accommodate life for about 1 Gyr beyond other surface environments.
A model was developed to investigate the effects such life will have on its environment. Future work will extend this to investigate the likely biosignatures produced by life in these localized specific environments, both on Earth and on any habitable exoplanet in the late stages of its development to answer the questions of whether a vastly reduced biosphere could produce remotely detectable biosignatures and, by extension, at what point an Earth-like planet would stop exhibiting remotely detectable biosignatures.
Acknowledgements
The authors wish to thank Jelte Harnmeijer, Christiane Helling and Anja Bauermeister for helpful discussions and Shawn Domagal-Goldman and an anonymous reviewer for comments that significantly improved the manuscript. Support from the Science and Technology Facilities Council (STFC) Aurora scheme is gratefully acknowledged. The University of Dundee is a registered Scottish Charity, No. SC015096.










