Introduction
Exotic annual grass invasions such as downy brome (Bromus tectorum L.) and medusahead [Taeniatherum caput-medusae (L.) Nevski] have transformed landscapes in the Great Basin and surrounding ecoregions by displacing native perennials and other taxa, creating less stable and suitable habitat for wildlife, altering biogeochemical processes, and increasing wildfire recurrence (Balch et al. Reference Balch, Bradley, D’Antonio and Gomez-Dans2012; DiTomaso Reference DiTomaso2000). Following fire events, landscapes that previously had low or moderate levels of exotic annual grasses can be highly susceptible to a full-scale invasion, particularly in years with high rainfall and at midlevel elevations (Chambers et al. Reference Chambers, Bradley, Brown, D’Antonio, Germino, Grace, Hardegree, Miller and Pyke2013). Compared with perennial grasses, exotic annual grasses (1) have earlier emergence and condensed life spans, allowing for a phenological advantage in resource use (Verdu and Traveset Reference Verdu and Traveset2005); (2) create large seedbanks (Diamond et al. Reference Diamond, Call and Devoe2010; Facelli et al. Reference Facelli, Chesson and Barnes2005); and (3) result in feedbacks that reinforce their dominance, such as altered nutrient cycling or increased wildfire occurrence (D’Antonio and Vitousek Reference D’Antonio and Vitousek1992; Germino et al. Reference Germino, Belnap, Stark, Allen and Rau2016; Knapp Reference Knapp1996).
Management Implications
Spraying of the herbicide imazapic is becoming widely used to temporarily reduce exotic annual grasses in semiarid landscapes; release growth of deep-rooted perennial bunchgrasses, forbs, and shrubs; and prepare soils for seeding treatments. It is labeled for both pre- and postemergent treatment of exotic annual grasses, but information is needed on how its effectiveness varies with timing of application relative to plant phenology following disturbances such as wildfire. In sagebrush steppe rangelands, imazapic can provide temporary exotic annual grass control with little or no impact to deep-rooted perennial bunchgrasses when applied either directly following fire or if delayed a growing season, but also could potentially impact native shallow-rooted bunchgrasses and some coseeded species with the earlier application. Spraying in fall of the second post-fire year was less effective on sites that had high exotic annual cover before spraying (>42% cover), likely due to litter accumulation and lack of herbicide−soil contact. Aerially seeded shrubs and forbs generally had no or positive responses to spraying 9 to 10 mo after seeding. In summary, imazapic reduced exotic annual grasses when applied at various times during the first post-fire year, but had more effects on non-target plant species when applied in the first winter post-fire. Thus, a treatment sequence that staggers herbicide and seeding of desirable shrubs and forbs may be a useful approach toward integrated weed management.
Land managers have identified key methodologies to target exotic annual grasses, particularly for use in post-fire rehabilitation, to prevent their further spread and maintain or restore perennial grasses (Monaco et al. Reference Monaco, Hardegree, Pellant and Brown2016). Establishing and maintaining persistent communities of perennials, especially native deep-rooted bunchgrasses, are critical for providing long-term resistance against invasion and resilience to future disturbances (Chambers et al. Reference Chambers, Bradley, Brown, D’Antonio, Germino, Grace, Hardegree, Miller and Pyke2013). Preemergent herbicides are an important tool for control, because they can limit germination, growth, and seed production of targeted exotic annuals, thereby reducing resident seedbanks (Brisbin et al. Reference Brisbin, Thode, Brooks and Weber2012; Hirsch et al. Reference Hirsch, Monaco, Call and Ransom2012) and releasing growth of desirable perennials (Madsen et al. Reference Madsen, Davies, Mummey and Svejcar2014; Sbatella et al. Reference Sbatella, Wilson, Enloe and Hicks2011; Sheley et al. Reference Sheley, Jacobs and Lucas2001). However, herbicides also have the potential for negative effects on non-target species (Sheley et al. Reference Sheley, Carpinelli and Reever Morghan2007; Su et al. Reference Su, Sun, Wu, Ma, Wang, Xu and Lu2017; Wallace and Prather Reference Wallace and Prather2016). Several herbicides used to control exotic annual grasses in the semiarid Great Basin include flufenacet plus metribuzin, propoxycarbazone-sodium, sulfosulfuron, rimsulfuron, and imazapic (Wallace and Prather Reference Wallace and Prather2016).
Imazapic, in particular, has become widely used in rehabilitation to control B. tectorum (Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, David and Menalled2013), ventenata [Ventenata dubia (Leers) Coss.] (Wallace and Prather Reference Wallace and Prather2016), and T. caput-medusae (Kyser et al. Reference Kyser, Wilson, Zhang and DiTomaso2013; Sheley et al. Reference Sheley, Carpinelli and Reever Morghan2007). Imazapic is a broad-spectrum herbicide, though fairly selective when applied at low rates (≤70 g ai ha−1; Bahm and Barnes Reference Bahm and Barnes2011). Imazapic may act either as a preemergent germination inhibitor (Shinn and Thill Reference Shinn and Thill2004) or as a postemergent herbicide, inhibiting the enzyme acetolactate synthase (Su et al. Reference Su, Sun, Wu, Ma, Wang, Xu and Lu2017). Despite imazapic’s widespread use, its effectiveness at controlling exotic annual grasses is highly variable (Morris et al. Reference Morris, Monaco and Rigby2009; Wallace and Prather Reference Wallace and Prather2016), and more information is needed about outcomes in different management situations. The high phenotypic plasticity in phenology of winter annuals such as B. tectorum (Rice and Mack Reference Rice and Mack1991) causes year-to-year variability that complicates treatment planning, because the timing of emergence can vary greatly. Applications of imazapic via spraying from aircraft over large areas (more than thousands or even tens of thousands of hectares) are increasingly common, and logistical challenges in timing of spraying contribute to the variability in outcomes.
In areas where deep-rooted perennial bunchgrasses have been depleted, such as through some livestock-grazing regimes, seeding bunchgrasses by aerial broadcast or with rangeland drills is common. Seedings often fail because of early competitive pressures from exotic annual grasses (Francis and Pyke Reference Francis and Pyke1996), thus requiring later treatment with herbicide. Integrated weed management, which can combine herbicide spraying and seeding, is a common component of many rehabilitation or restoration plans, and there has been substantial research focused on methods to pair herbicide and seeding (Bahm and Barnes Reference Bahm and Barnes2011; Kyser et al. Reference Kyser, Wilson, Zhang and DiTomaso2013; Madsen et al. Reference Madsen, Davies, Mummey and Svejcar2014; Sheley et al. Reference Sheley, Jacobs and Lucas2001; Washburn and Barnes Reference Washburn and Barnes2000). While herbicides may reduce competing exotics and provide bare soil for seeded species, they may also have an antagonistic effect if they damage the seeded species. Some deep-rooted perennial bunchgrasses may be more susceptible to imazapic during the germination and establishment stage (Sheley et al. Reference Sheley, Carpinelli and Reever Morghan2007) rather than as established plants. Additionally, imazapic can reduce big sagebrush (Artemisia tridentata Nutt.) germination by up to 80%, while older shrub seedlings may have greater resistance (Owen et al. Reference Owen, Seig and Gehring2011).
The season and prevailing phenological condition during imazapic application can influence its effectiveness. For example, imazapic applied in fall before emergence reduced B. tectorum less than postemergent applications in Montana grasslands (Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, David and Menalled2013). However, postemergent application in spring was less effective than fall application for T. caput-medusae control in shrub-steppe of Utah (Monaco et al. Reference Monaco, Osmond and Dewey2005). Application rate (Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, David and Menalled2013; Morris et al. Reference Morris, Monaco and Rigby2009; Sheley et al. Reference Sheley, Carpinelli and Reever Morghan2007), soil type (Hirsch et al. Reference Hirsch, Monaco, Call and Ransom2012), and weather (Morris et al. Reference Morris, Monaco and Rigby2009) may all impact success rate. High precipitation before herbicide application can cause favorable synchronization of seedling growth and herbicide uptake, but high precipitation postapplication can increase risks of runoff and leaching (Martini et al. Reference Martini, Mezzomo, Antonio de Avila, Massey, Marchesan, Zanella, Peixoto, Refatti, Cassol and Marques2013; Morris et al. Reference Morris, Monaco and Rigby2009). While preemergent and very early postemergent effects of imazapic have been evaluated, much less is known about its effects when applied after the growing season of the initial exotic annual grass flush (e.g., with the goal of inhibiting a subsequent crop), a time when vast landscapes are often treated (Table 1).
Table 1 Previous publications on the effects of imazapic on different functional groups and the current findings of our study.Footnote a

a Blank cells indicate no publications or data.
b Seeded species abbreviations: ACMI, Achillea millefolium; ARTEM, Artemisia spp.; MESA, Medicago sativa; SAMI, Sanguisorba minor.
c The results indicate effects as follows: –, decreased abundance compared with control; =, abundance same as control; +, increased abundance compared with control.
Imazapic is used to target exotic annual grasses but can potentially have unintended effects on other functional groups (e.g., non-target species), particularly (1) short-lived, shallow-rooted species (such as Sandberg bluegrass [Poa secunda J. Presl]), (2) long-lived, deep-rooted species (native perennial forbs, deep-rooted perennial bunchgrasses), and (3) those species seeded in conjunction with herbicide treatment (Table 1). For preemergent application, non-target plants with short life histories are often susceptible to injury or reductions in cover (Davies and Sheley Reference Davies and Sheley2011; Davies et al. Reference Davies, Madsen, Nafus, Boyd and Johnson2014; Elseroad and Rudd Reference Elseroad and Rudd2011; Owen et al Reference Owen, Seig and Gehring2011), although Davies (Reference Davies2010) and Davies et al. (Reference Davies, Madsen, Nafus, Boyd and Johnson2014) found either neutral or positive responses (i.e., release from competition) of these species. Preemergent imazapic application effects on non-target deep-rooted perennials are highly variable, ranging from apparent release (Bahm and Barnes Reference Bahm and Barnes2011; Morris et al Reference Morris, Monaco and Rigby2009; Owen et al. Reference Owen, Seig and Gehring2011; Washburn and Barnes Reference Washburn and Barnes2000) to injury or mortality (Davies et al. Reference Davies, Madsen, Nafus, Boyd and Johnson2014; Hirsch et al. Reference Hirsch, Monaco, Call and Ransom2012; Sheley et al. Reference Sheley, Carpinelli and Reever Morghan2007; Shinn and Thill Reference Shinn and Thill2004; Wallace and Prather Reference Wallace and Prather2016) or no changes (Bahm and Barnes Reference Bahm and Barnes2011; Shinn and Thill Reference Shinn and Thill2004).
We assessed the effects on exotic and native plant communities of replicated and landscape-scale imazapic application made on portions of the 113,000 ha Soda wildfire (started August 10, 2015) along the southern Idaho–Oregon border. We asked the following questions about how the timing of imazapic application impacts native and exotic species:
1. What is the effect of spraying in the first winter or second fall post-fire compared with control (no spraying) on community composition and cover of dominant plant functional groups?
2. What is the effect of spraying on occurrence frequency of seeded perennial shrubs and forbs?
3. Which environmental or landscape factors influence the plot-scale effectiveness (pre- and posttreatment exotic annual grass cover) of imazapic applied in the second fall post-fire?
We anticipated that imazapic application would reduce exotic annual grasses and have no effect on or increase perennial grass and forb cover, especially with recent post-fire application within the proper treatment window. We also expected that herbicide application integrated with shrub and forb seeding would decrease abundances of the seeded species, while staged interventions (seeding followed by waiting until the fall after seeding to apply herbicide) would increase abundances of seeded species.
Materials and Methods
Study Area
Mean annual precipitation ranges from 23.3 to 55 cm and mean annual temperature ranges from 6.8 to 10.8 C across the study area (800-m2 pixel PRISM data, 1950–2014). Elevation ranges from 701 to 2,054 m (U.S. Geological Survey’s Digital Elevation Model, 30-m pixel). Growing season precipitation (March to June) was 70 mm in 2016 and 132 mm in 2017, measured at the Idaho Department of Transportation (DOT; Ion station) weather station in the western section of the fire at 1,264-m elevation. Winter of 2016–17, with mean December and January daily temperatures of approximately −5 C, was colder than the winter of 2015–16, with mean December and January daily temperatures of approximately −1 C. The mean daily summer temperature was approximately 21 C between June and August 2016 and approximately 23 C between June and August 2017 (Idaho DOT).
Herbicide Applications and Seeding Treatments
Although additional herbicide applications were made at different times across the Soda wildfire, we limit our analysis to the following treatments: Imazapic (Plateau®, BASF, Research Triangle Park, NC) was applied at a rate of 100 g ai ha−1 with 1.7 kg ha−1 methylated seed oil (MSO) by the Idaho Bureau of Land Management (BLM) on different treatment areas in Idaho that were a combined 8,821 ha in February–March 2016, and 13,060 ha in October 2016 (Figure 1). Imazapic was applied to an additional 12,287 ha in Oregon at the same rate (100 g ai ha−1) with 1.1 kg ha−1 nonionic oil concentrate and 0.4 L 100 L−1 water softener (AccuQuest®, Helena Chemical, Collierville, TN) in September 2016.
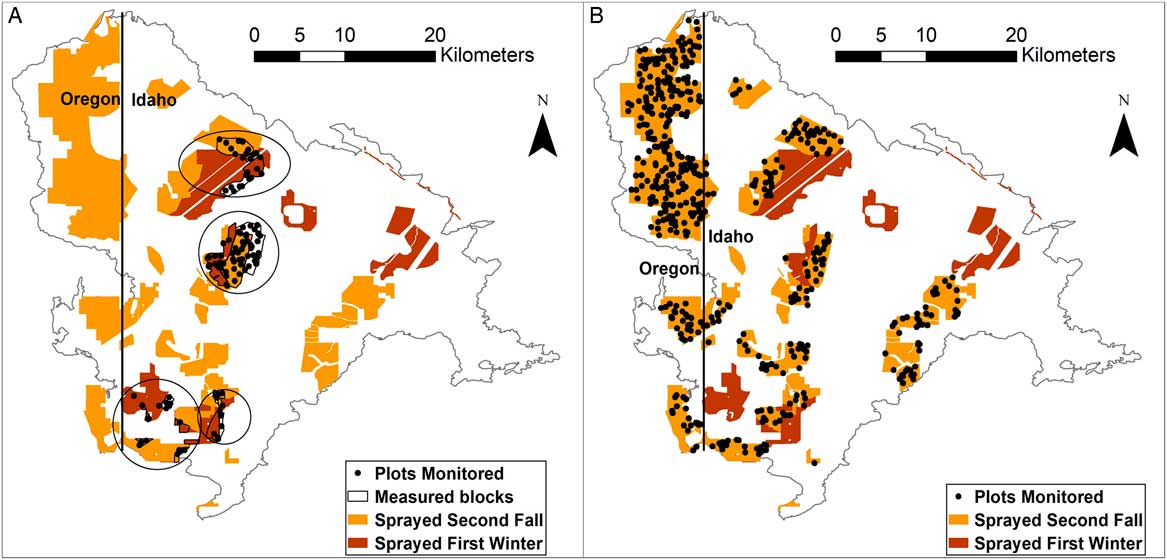
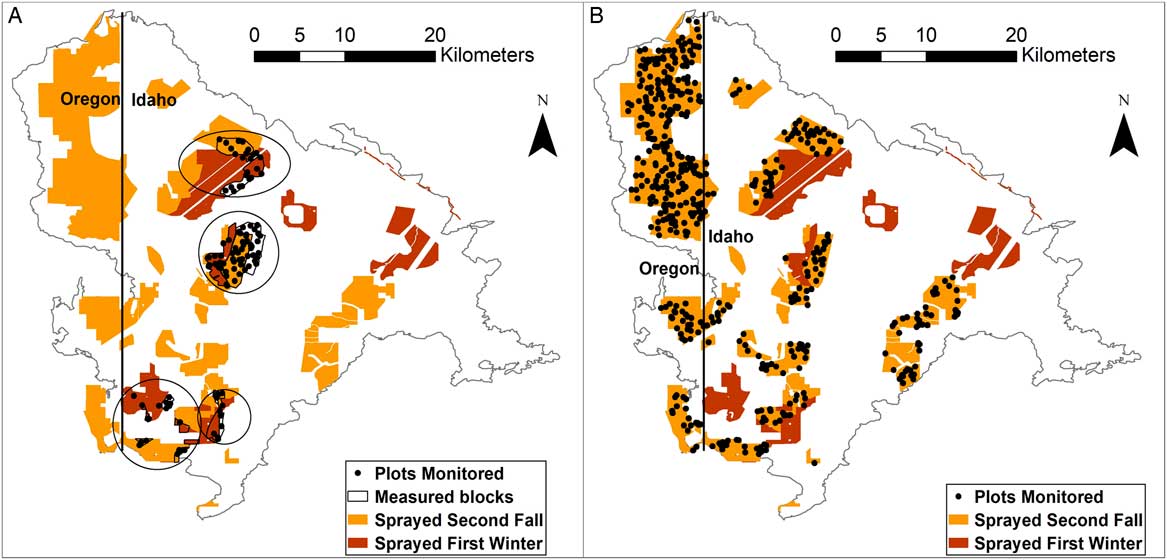
Figure 1 Location of herbicide spraying post-fire (sprayed first winter or sprayed second fall). (A) The location of the boundaries of post hoc analysis blocks (measured blocks) and plots within blocks (plots monitored) used to assess the effects of spraying herbicide on plant community and functional group cover between control (no herbicide) areas and areas sprayed either in the first winter or areas sprayed in the second fall post-fire. Regions in which the blocks occur are circled, but the plots monitored points show exact locations of data collection. (B) The location of all plots monitored in the second fall post-fire herbicide treatment that were used for the pre/posttreatment analysis of landscape factors on herbicide variability.
Exotic annual grass phenology was not assessed before the February–March 2016 application (first winter application), but given that winter and fall precipitation was average, we assume that this treatment was made at a postemergent stage for B. tectorum. New growth of exotic annual grasses was not observed during the September or October 2016 (second fall) applications, indicating that this treatment was preemergent for exotic annual grass species. Forb and Artemisia spp. aerial seedings were conducted in January 2016 (see Table A.1. in the Supplementary Material for details). Additional treatments in 2016 included aerial seeding of perennial grasses and forbs using aircraft and drill seeding of perennial grasses (Table A.1 in the Supplementary Material). We did not include any areas that were drill seeded in our analysis.
Selection of Post Hoc Treatment Blocks (Herbicide in First Winter or Second Fall Compared with Control)
The following block selection was constrained to the two Idaho herbicide treatments in the first winter (February–March 2016) or second fall (October 2016); to isolate the effects of herbicide on plant communities and identify “control” areas, we conducted a post hoc site matching of areas where herbicide was applied versus areas where it was not applied based on similar soil-mapping units (USDA-NRCS 2017), elevation, and topography (Figure 1A). We also consulted with the BLM staff (personal communication 2018) who designed and oversaw implementation of treatments to eliminate areas that had potentially confounding factors and ensure that our assumptions about equivalency of (1) pre-fire community condition between treated and control areas and (2) imazapic application rates and conditions among treated areas (e.g., equipment used, wind drift) within statistical blocks were sound. Aerial grass seedings were included in some blocks, because the very small number of seedlings resulting from these treatments had a negligible effect on overall first- and second-year perennial grass cover (grass seedlings of seeded species were generally not observed in the SamplePoint photos in these treatments; Table A.1 in the Supplementary Material). To compare the effects of herbicide applied in the first winter or herbicide applied in the second fall with unsprayed areas, we identified four blocks that each contained an untreated site, a site sprayed in the first winter post-fire (February–March 2016), and a site sprayed in the second fall post-fire (October 2016) (Figure 1A). These blocks were generally in moderately invaded areas where control areas were available for comparison. Each block was considered one replicate and ranged from 407 ha to 2,069 ha in size.
To assess multiple treatment effects (shrub/forb seeding plus herbicide), we identified blocking units using the same site-matching criteria described earlier, but with the additional requirements of matching seed mixes and excluding blocks where no seeded shrubs or forb established (Figures A.1. and A.2. in the Supplementary Material). We could not create blocks with all herbicide-timing combinations, so blocks containing first winter applications and controls (3 blocks) were sometimes spatially separate from those containing second fall applications and controls (4 blocks).
Selection of Areas for Pre/Posttreatment Analysis
We selected all areas in the two second-fall, post-fire herbicide applications (September 2016 in Oregon and October 2016 in Idaho) to evaluate the influence of landscape factors on treatment effectiveness (Figure 1B).
Data Collection
Data collection occurred during the growing seasons of 2016 and 2017 at or near the time of peak biomass for each plot. Plot locations were determined via a stratified-random method, with ≥1 plot 54.5 ha−1, and were moved if they overlapped roads, had >20% trail area within a 18-m radius, or fell within 0.40 km of water troughs or ponds. The percent cover of each species was determined from 6-m2 rectangular aerial photographs at plot center captured from nadir at 2-m height (with Nikon Coolpix AW130, 16 megapixel, Nikon Inc., 1300 Walt Whitman Road, Melville, NY). We analyzed species cover using grid-point intercept (GPI) software (SamplePoint v. 1.43, 100 points image−1; Booth et al. Reference Booth, Cox and Berryman2006). Standing dead cover was classified in the same way as live, but detached cover was classified as litter. We then grouped species cover into functional groups (Table A.2. in the Supplementary Material). By 2017, plants had become taller, and we thus made a correction for parallax distortion in which only the central 3-m2 area of each aerial photograph was evaluated (49 points image−1). A second overhead photograph (5.5 m to the south) was collected for each plot to keep the total amount of area sampled similar to 2016.
In addition to the GPI analysis, we conducted frequency-density monitoring for seeded shrubs and forbs by counting seeded shrub and forb individuals at the center 1-m2 quadrat and progressing outward at radiuses of 5.5, 9, 13, and 18 m until three individuals of each species were found. Once three individuals of the species were found, we counted all individuals of that species within the given radius. This method is a concentric-rings adaptation of nested frequency density (Bonham Reference Bonham2013; Mueller-Dombois and Ellenburg Reference Mueller-Dombois and Ellenburg1974). Because A. tridentata and little sagebrush (Artemisia arbuscula Nutt.) were virtually indistinguishable in the field as seedlings, we did not distinguish between the two and subsequently refer to them both as Artemisia spp., although A. tridentata was seeded over a substantially larger area and at a higher rate (Table A.1. in the Supplementary Material).
Candidate landscape variables were obtained from several databases: mean annual precipitation from PRISM (2017), elevation from U.S. Geological Survey Digital Elevation Models (10-m pixels), pre-fire exotic annual grass cover from the 2011 National Land Cover Database (Homer et al. Reference Homer, Dewitz, Yang, Jin, Danielson, Xian, Coulston, Herold, Wickham and Megown2015), and soil temperature moisture regime (subclass) from the USDA Natural Resources Conservation Service Web Soil Survey (USDA-NRCS 2017). Aspect, slope, heat load, and topographic wetness index (TWI) were calculated from elevation in ArcGIS. Heat load is an index ranging from 0 to 1 and is based on topographic effects on solar gain (McCune and Keon Reference McCune and Keon2009). TWI is used to explain the influence of topography on hydrological processes (Sorensen et al. Reference Sorensen, Zinko and Seibert2006). Species richness was calculated by summing all the species we found in each 13-m plot. Wildfire burn history was calculated by summing the total number of times a plot burned between 1960 and 2014 based on BLM fire records. Pretreatment exotic annual grass species dominance was calculated as the cover of B. tectorum minus the cover of T. caput-medusae from 2016 monitored plot values (pretreatment values). Pretreatment exotic annual grass cover and perennial bunchgrass cover were also included from 2016 monitored plot values.
Data Analysis
Herbicide in First Winter or Second Fall Compared with Control (Post Hoc Analysis)
Community response
We analyzed second-year (2017) data by species cover and by functional group cover for the 114 plots across four blocks (Figure 1A). For individual species cover, we ran two multiresponse blocked permutation procedures (MRBP) in the ‘Blossum’ package in R Studio to compare differences between the control and each herbicide application (R Core Team 2017; Talbert and Cade Reference Talbert and Cade2013). Additionally, we calculated the Shannon diversity index (SI) of cover for each plot and analyzed the differences between treatments with a two-way ANOVA with herbicide treatment and block as factors and performed a Dunnett’s post hoc test to compare the control and each herbicide application.
Functional group cover
Functional group cover data (exotic annual grass, perennial bunchgrass, P. secunda, and native perennial forb species classifications in Table A.2. in the Supplementary Material) were highly right-skewed and had unequal variances. Thus, we used separate beta generalized linear mixed models (GLMMs) to detect treatment differences in bare soil and cover of each functional group (‘glmmTMB’ package in R; Magnusson et. al Reference Magnusson, Skaug, Nielsen, Berg, Kristensen, Maechler, van Bentham, Bolker and Brooks2017), as suggested for plant abundance data (Damgaard Reference Damgaard2009; Eskelson et al. Reference Eskelson, Madsen, Hagar and Temesgen2011). We transformed cover data following the standard set by Smithson and Verkuilen (Reference Smithson and Verkuilen2006) for beta regressions. Herbicide treatment was considered a fixed effect and block was considered a random effect. Because we could not effectively control aspect during site matching, heat load index was considered as a covariate in each model but was only included in the exotic annual grass model, because likelihood ratio tests with the reduced model showed no improvement in fit for other functional groups. Dunnett’s post hoc tests were used to analyze the contrasts between treatment means and control means via the ‘lsmeans’ package (Lenth Reference Lenth2016). Fitted means from the beta GLMM regressions were within 2% cover of sample means. Estimated marginal means are reported (the effect of the herbicide treatments is adjusted for other model effects).
Frequency of Seeded Shrubs and Forbs
We compared seeded shrub and forb abundances within seventy-three 18-m frequency-density plots that were measured in 2016, in three blocks that each contained an area sprayed with herbicide directly after shrub and forb seeding (first winter) and a separate unsprayed area (Figure A.1. in the Supplementary Material). Some of these areas were seeded again in October 2016, so we did not assess 2017 data in these blocks. We also compared 2017 seeded shrub and forb abundance in 79 plots within four different blocks each containing an area sprayed with herbicide the season after shrub/forb seeding (second fall) and an unsprayed area (Figure A.2. in the Supplementary Material). We modeled presence/absence of Artemisia spp. and forbs using GLMMs with a binomial distribution in response to herbicide treatment as a fixed effect and block as a random effect (‘glmmTMB’ package; Magnusson et al. Reference Magnusson, Skaug, Nielsen, Berg, Kristensen, Maechler, van Bentham, Bolker and Brooks2017).
Influence of Landscape Factors on Treatment Effectiveness for Second Fall Herbicide Applications (Pre/Posttreatment Analysis)
Plots sprayed in September or October 2016 (second fall) were monitored before spraying in summer 2016 and after spraying in summer 2017. Thus, we could assess how landscape variability modified herbicide effectiveness at the plot scale on these 498 plots across the entire fire. We used the random forest algorithm (‘randomForest’ package; Liaw and Wiener Reference Liaw and Wiener2002) to analyze the effects of the following landscape variables on posttreatment exotic annual grass cover: application method (Oregon treatment conducted with nonionic oil concentrate or Idaho treatment conducted with MSO), pretreatment exotic annual grass cover, pretreatment perennial bunchgrass cover, pretreatment exotic annual grass species dominance, mean annual precipitation, elevation, 2011 National Land Cover Database modeled pre-fire exotic annual grass cover (from Homer et al. Reference Homer, Dewitz, Yang, Jin, Danielson, Xian, Coulston, Herold, Wickham and Megown2015), slope, TWI, heatload, species richness, aspect, wildfire burn history, and soil temperature moisture regime subclass. To identify the effects of the most important variables, we selected the top four variables in order of explanatory importance from the random forest output and ran a classification and regression tree (CART) model (‘rpart’ package; Therneau et al Reference Therneau, Atkinson and Ripley2017) using these variables and pruned the resulting tree using cost-complexity pruning.
Results and Discussion
Herbicide in First Winter or Second Fall Compared with Control (Post Hoc Analysis)
Community and Functional Group Responses
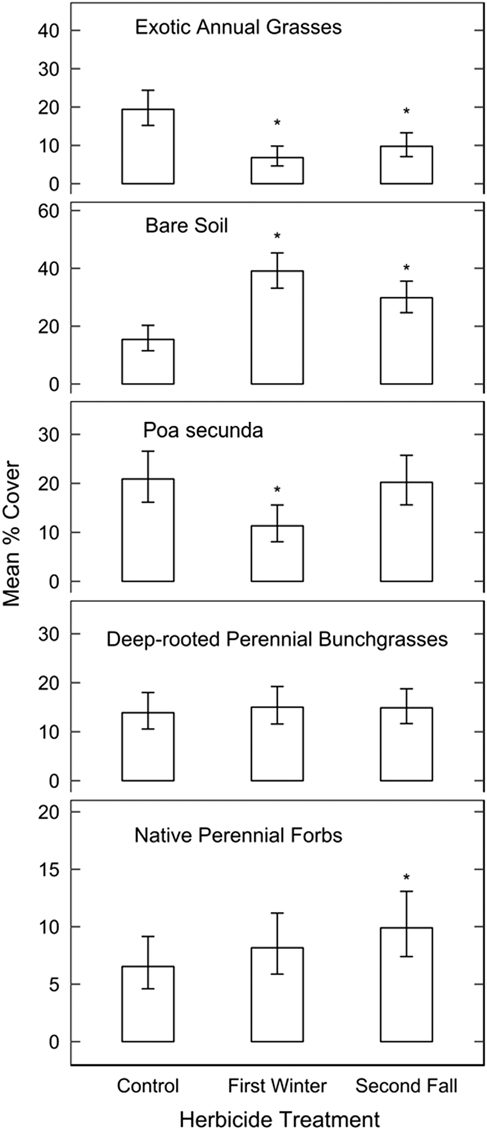
There were no significant differences in the overall species community compositions measured in 2017 (MRBP, P>0.05) or by the Shannon-Weiner diversity index between control plots and plots sprayed with herbicide in either the first winter or second fall post-fire. However, exotic annual grass cover was approximately 19% in the control compared with 7% in the areas sprayed in the first post-fire winter (P=0.0004) and 10% in the second fall application (P<0.0001; Figure 2). Compared with control areas, bare soil exposure was 2.5 times greater in areas sprayed in the first winter (P<0.001) and 2 times greater in areas sprayed in the second fall (P=0.0004). Poa secunda cover was 45% lower in areas sprayed in the first winter compared with control (P=0.005) but was not responsive to the second fall spraying. Cover of native perennial forbs was higher in the second fall treatment compared with control (P=0.05) but not different between the first winter treatment and control. Cover of deep-rooted perennial bunchgrasses was not affected by herbicide (P>0.05 for all contrasts).
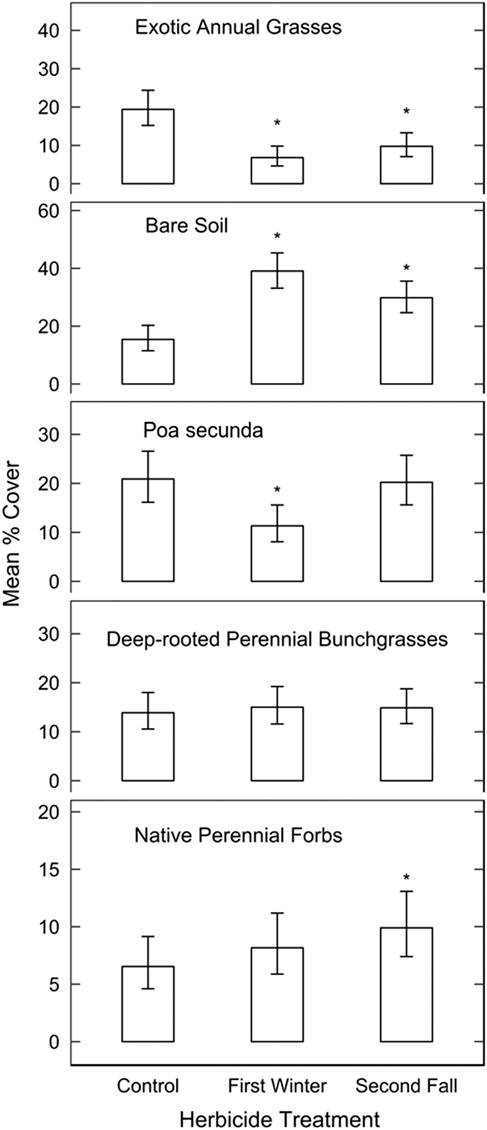
Figure 2 Estimated mean percent cover and 95% confidence intervals of different functional groups measured in 2017 from a mixed beta regression model in control plots and plots sprayed in the first winter or second fall post-fire. Exotic annual grass treatment effects are conditional on a constant heat-load index (not included in other functional group models). Asterisks denote treatments that are significantly different from untreated controls.
Spraying in either the first winter or second fall appeared to provide temporary reductions in exotic annual grass cover compared with control areas in moderately invaded areas (generally <50% exotic annual grass cover). In contrast to the reduction in P. secunda cover we observed following the first spraying, P. secunda was not reduced by imazapic application at the small-plot level in previous studies (Davies Reference Davies2010; Davies and Sheley Reference Davies and Sheley2011; Davies et al. Reference Davies, Madsen, Nafus, Boyd and Johnson2014). A key distinction of these other studies was their use of fall or spring prescribed burning before herbicide application, which likely resulted in lower P. secunda injury before the herbicide was applied. In our study, P. secunda response to the early herbicide application may have been relatively sensitized by several years of drought conditions combined with a higher-intensity fire. Given a growing season to recover before herbicide spraying, P. secunda appeared resistant to imazapic. As with any post hoc analysis, we are limited in our knowledge of pre-fire vegetation and cannot completely rule out the possibility of preexisting differences in P. secunda between areas sprayed and not sprayed.
The resistance of deep-rooted perennial bunchgrass and forb cover to imazapic may relate to their being largely comprised of regrowth of plants established before the fire, rather than seedlings. Our focus on measurements of cover may not capture or reveal injury that could be caused by imazapic application (as in Shinn and Thill Reference Shinn and Thill2004), but our results indicate that even if injury did occur, it did not significantly impact perennial native cover as a community component. Established perennial grasses and forbs were also unaffected by imazapic in other studies (Davies and Sheley Reference Davies and Sheley2011; Elseroad and Rudd Reference Elseroad and Rudd2011; Kyser et al. Reference Kyser, Wilson, Zhang and DiTomaso2013; Monaco et al. Reference Monaco, Osmond and Dewey2005).
Seeded Shrub and Forb Frequency
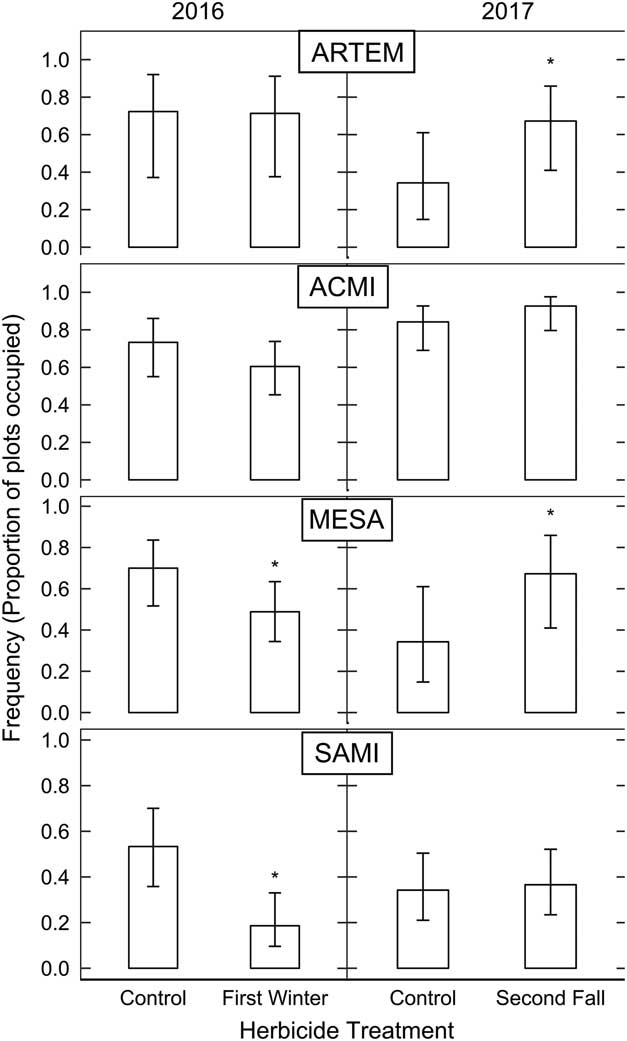
Abundances of Artemisia spp. and yarrow (Achillea millefolium L.) did not differ in areas sprayed in the first winter compared with control areas (Figure 3). Alfalfa (Medicago sativa L.) and small burnet (Sanguisorba minor Scop.) were less common in plots sprayed in the first winter compared with the control (P=0.07 and 0.003, respectively). Artemisia spp. were approximately 1.5 times as likely and M. sativa was approximately twice as likely to occur in areas sprayed in the second fall compared with control areas (P=0.01 and 0.008, respectively). Sanguisorba minor and A. millefolium were equally likely to occur in plots sprayed in the second fall as in the control (P>0.05; Figure 3).

Figure 3 Estimated frequency of 2016 species occupancy in January 2016 between unsprayed control plots and plots sprayed in the first winter post-fire from a mixed-effects logistic regression with 95% confidence intervals (left). Estimated frequency of 2017 seeded species occupancy between unsprayed control plots and plots sprayed in the second fall post-fire is displayed on the right. ACMI, Achillea millefolium; ARTEM, Artemisia spp.; MESA, Medicago sativa; SAMI, Sanguisorba minor. Asterisks denote treatments that are significantly different from untreated controls.
These results agree with Owen et al. (Reference Owen, Seig and Gehring2011), who found that preemergent herbicide application decreased germination but not seedling survival (4-mo-old) of A. tridentata. Our findings indicate that other perennial seeded forbs may be susceptible to preemergent imazapic application but can withstand exposure after a growing season and even increase, presumably due to release from competition. Sanguisorba minor was present in fewer plots than the other seeded species in general, but was further reduced by herbicide application in the first winter and received no benefit from application in the second fall. The low overall occurrence of S. minor is unsurprising, because it was seeded at a rate that was less than 20% that of the other seeded species.
Landscape Factor Effects on Treatment Effectiveness for Second Fall Herbicide Applications (Pre/Posttreatment Analysis)
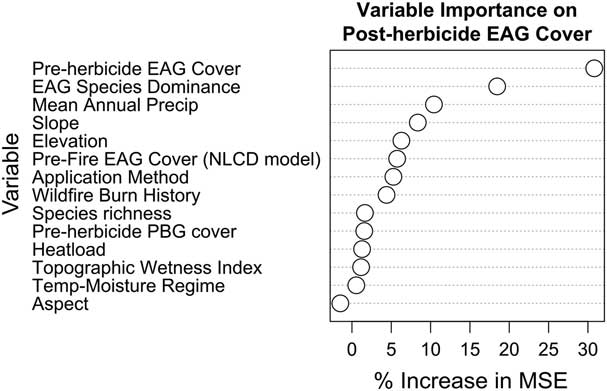
The landscape factors analyzed did not explain much of the variability in changes in exotic annual grasses associated with imazapic applied in the second fall after fire. First-year exotic annual grass cover (measured in 2016 before treatment) accounted for the vast majority of model variance explained (Figure 4), and the entire random forest model only accounted for 38% of the variance in posttreatment exotic annual grass cover. A CART model that included the top four random forest variables resulted in a single threshold: exotic annual grass cover was reduced to 10% where imazapic had been applied to plots initially (pretreatment) having up to 42% exotic annual grass cover. However, posttreatment exotic annual grass cover was much greater (39%) following the spraying of plots that had >42% cover pretreatment. Previous studies have reported very different effects of imazapic at different sites (Morris et al. Reference Morris, Monaco and Rigby2009; Wallace and Prather Reference Wallace and Prather2016), and we also observed a large degree of variability in exotic annual grass cover that was unaccounted for by any factors we tested (62%). The 42% threshold in pretreatment herbicide cover we identified indicates that small-scale patches of high invasion may resist post–growing season herbicide application. This may be due to first-year litter accumulation, which could (1) intercept herbicide and (2) reduce our ability to detect herbicide effects on emergence when senesced stems are still standing and rooted into the subsequent growing season and measurement period following herbicide application. Previous studies have shown that removing litter by mowing or burning can help with exotic annual grass suppression by imazapic when litter cover is high (Kyser et al. Reference Kyser, DiTomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007; Morris et al. Reference Morris, Monaco and Rigby2009).

Figure 4 Random forest variable importance plot showing the importance of each landscape variable on predicting postherbicide exotic annual grass (EAG) cover in 2017. EAG species’ dominance is a metric calculated by subtracting the 2016 (preherbicide) cover of Taeniatherum caput-medusae from the 2016 cover of Bromus tectorum. Pre-fire EAG cover (NLCD model) is derived from 2011 National Land Cover Database (Homer et al. 2015). Preherbicide EAG cover and preherbicide perennial bunchgrass cover (PBG) are also given from 2016 field monitoring. The x-axis quantifies the percent mean increase in mean squared error (MSE) as a result of a given variable being permuted. The higher the percent increase in MSE as a result of a variable being permuted, the more important that variable is to explaining variability in postherbicide EAG cover.
Synthesis
The use of imazapic as a post-fire rehabilitation tool has typically focused on immediate treatment after disturbance, specifically application in the first fall after fire. Our results provide a landscape-scale example in which exotic annual grass cover was lower in areas where herbicide application was delayed until the second fall post-fire when compared with untreated areas.
Herbicide is usually applied as a temporary first step to reduce competition before seeding (especially seeding with rangeland drill), and this sequence is typically conducted by spraying in the first fall post-fire so that drill seeding can occur after the growing season. Herbicide alone will often not provide long-term reductions in exotic annual grasses without establishment of deep-rooted perennial bunchgrasses and the resistance to invasion they provide (Davies et al. Reference Davies, Boyd, Johnson, Nafus and Madsen2015). In areas where both aerial shrub and forb seeding and herbicide occurred, delaying herbicide 9 to 10 mo after seeding led to a more favorable outcome than co-application within a 1-mo period. Nonetheless, applying herbicide within a month after seeding did not eliminate the seeded species, and when this degree of co-application must occur (e.g., due to operational or logistical constraints), the small losses in non-target species might be mitigated with increased seeding rates. More commonly, a treatment sequence of herbicide followed by seeding is used, as it poses less risk to the seeded species, and while this sequence was originally intended following the Soda wildfire, late-fall freezing precluded its implementation. This multiple-entry method has been effective with establishment of seeded deep-rooted perennial bunchgrasses (Davies et al. Reference Davies, Madsen, Nafus, Boyd and Johnson2014; Sbatella et al. Reference Sbatella, Wilson, Enloe and Hicks2011).
Acknowledgments
Funding provided by the BLM, Great Basin Landscape Conservation Cooperative Program, and the Joint Fire Science Program. Logistics, support, and review comments were graciously provided by Cindy Fritz, Peter Torma, Amy Stillman, Robert Bennett, and Cara Hastings at the Boise BLM. Review comments were also provided by Dave Barnard at U.S. Geological Survey. Field data were collected by Julia Aaronson, James Albertson, Andrew Blomberg, Vanessa Callahan, Rebecca Donaldson, Emma Donohoe, Brian Fischer, Lauren Hunsaker, Castilleja Kuzis, Andrew Lague, Thomas Laird, Jessica Moran, Harold Pearson-Nadal, Rae Robinson, Calene Thomas, Jeffery Titcomb, Mamadou Toure, Merry V. M. Davidson, Patrick Weinberg, and Wilson Yates. No conflicts of interest have been declared. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. government.
Supplementary materials
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2018.18