INTRODUCTION
The story of human occupations in southeastern Mesoamerica begins at least 11 millennia before the first settled villages were surrounded by maize fields. Those first tropical foragers set in motion cultural adaptations and environmental impacts that continue today. Surprisingly, we still know relatively little about diet, technology, and relationships between people who lived in southern Mesoamerica and their neighbors. Despite decades of study, much of our understanding of the earliest populations in the neotropical rainforests is inferred from research in central and northern Mexico, North America, South America, and southern Central America, partially due to dense forest cover impeding identification of early sites and to a lack of archaeological focus on the earliest humans in the region.
In this article we examine the relationships between foraging and early horticultural populations living in the neotropics. During the Late Pleistocene (>12,000–10,000 b.c.), southeastern Mesoamerica was part of a mixing area for early Clovis-like and Fishtail-like technologies extending from southern Mexico to Northern Colombia (Pearson Reference Pearson2017). Terminal Pleistocene and Early Holocene (10,000–6000 b.c.) foragers in Belize likewise shared technological ties with their contemporaries in the neotropics of southern Mexico, Central America, and South America, evidenced by similarities in stone tools (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019; Ranere and Cooke Reference Ranere and Cooke2021). By the Middle Holocene (6000–2500 b.c.) these ties are less evident, as northern South America becomes a major locus of domestication and horticulture, with several early staple foods being exploited across vast swaths of northern South America between 6000 and 4000 b.c. (Iriarte et al. Reference Iriarte, Elliott, Yoshi Maezumi, Alves, Gonda, Robinson, de Souza, Watling and Handley2020), at least 1,000–1,500 years earlier than in southeastern Mesoamerica. Initial maize consumption, evidenced by elevated carbon stable isotopes values, is first evident in South America after 4000 b.c., with evidence of consumption of maize as a staple grain by some individuals around 3000 b.c. (Tung et al. Reference Tung, Dillehay, Feranec and DeSantis2020). In southeastern Mesoamerica the consumption of maize in sufficient quantities to be detected with stable isotopes begins 1,300 years later—again, as a gradual process over a period of seven centuries, leading to the adoption of maize as a staple grain by 2000 b.c. (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020).
The Neotropics are a biogeographical area marked by similar plant and animal species distributions (Kreft and Jetz Reference Kreft and Jetz2010) that extends roughly from the Isthmus of Tehuantepec southward to and including Central America and much of northern and central South America. We hypothesize here that this vast area was an important sphere of interaction prior to cultivation of domesticated plants during the Early and Middle Holocene (e.g., Piperno and Pearsall Reference Piperno and Pearsall1998). We present several lines of evidence to support our hypothesis of a complex history. First, genomic data indicate that early foraging populations in Belize, around 7600–5300 cal b.c., descended from ancient Late and Terminal Pleistocene neighbors to the north, but shared more alleles with neighbors in southern Central America and South America (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018). Second, after approximately 9000 cal b.c., people living in several areas of the neotropics begin to produce diverse types of bifacial tools that were stemmed and barbed, suggesting that hunting and animal-processing toolkits were adapted to emerging tropical forest ecosystems and that core concepts of form and function were shared across this vast region (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). By around 6000 cal b.c. our data and studies from the Isthmo-Colombian region indicate that, with few exceptions, people stopped producing formal bifacial tools, corresponding in time with an increasing reliance on plants and less reliance on hunting large mammals (Ranere and Cooke Reference Ranere and Cooke2021). This pattern contrasts with central Mexico, where there is abundant evidence for the production of bifacial points throughout the Holocene (MacNeish et al. Reference MacNeish, Johnson and Nelken-Terner1967), consistent with studies in North America (Justice Reference Justice2002). Third, very early domesticates or their wild precursors were being consumed in South America after approximately 7000 cal b.c., with Cucurbita (squash), Capsicum (chili pepper), Manihot (manioc), and Phaseolus (beans) appearing initially in the archaeological record (Iriarte et al. Reference Iriarte, Elliott, Yoshi Maezumi, Alves, Gonda, Robinson, de Souza, Watling and Handley2020). Zea mays (maize) underwent partial domestication in Mexico around 7000 cal b.c. (Piperno et al. Reference Piperno, Ranere, Holst, Iriarte and Dickau2009), was introduced into South America before approximately 6200 cal b.c., and, unimpeded by introgression from its wild precursor, underwent a set of secondary improvements there that resulted in the fixing of traits associated with modern maize (Kistler et al. Reference Kistler, Yoshi Maezumi, de Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Thomas, Gilbert, de Oliveira Freitas and Allaby2018). Genetic evidence suggests that improved landraces spread northward into Central America and were likely influential for the development of other highly productive varieties already under selective pressure in Mesoamerica (Kistler et al. Reference Kistler, Thakar, VanDerwarker, George, Harper, Allaby, Hirth and Kennett2020). We show direct evidence that in southeastern Mesoamerica, the initial shift to a diet focused on maize started around 2750 cal b.c., with some individuals consuming modest amounts of maize (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). By around 2050 cal b.c., maize was fully adopted as a staple grain in southeastern Mesoamerica. These data also suggest that earlier varieties of maize (Kistler et al. Reference Kistler, Yoshi Maezumi, de Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Thomas, Gilbert, de Oliveira Freitas and Allaby2018), which were grown across the neotropics, were not being consumed in appreciable quantities in southeastern Mesoamerica (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020), suggesting that movement of more productive maize northward may have involved sharing of both crops and technologies.
Below, we detail cultural developments in the neotropics from the Late Pleistocene though the Middle Holocene, with a focus on the genomic history, early stone tool technologies, and initial use of important plants that eventually defined the diets of people living in the region. We then describe two archaeological sites in southern Belize (Figure 1b) that were persistently used as mortuary sites throughout the Holocene and also contain significant evidence of stone tool processing and use, with attention to changes in the lithic toolkit, the genetic structure of the earliest inhabitants, and the results of isotopic studies of 27 directly dated skeletons dating from around 7650–2000 cal b.c.

Figure 1. Maps indicating (a) major pre-agricultural sites and study areas in the neotropics discussed in the manuscript; and (b) southern Belize and the locations of Mayahak Cab Pek and Saki Tzul rock shelters in the Bladen Nature Reserve, along with other important archaeological sites in southern Belize. Map by Prufer and A.E. Thompson.
Chronological Considerations
Generally lacking chronological data to understand when meaningful changes in subsistence and technology occurred in the Late Pleistocene and Early Holocene, archaeologists turned to North American time periods to delineate changes in Mesoamerica. Those chronologies have been based primarily on changes in stone tool technologies that in North America are anchored by absolute dating methods. In Mesoamerica, the generalized preceramic periodization consists of the Paleoindian Period, from 11,500 to 8000 b.c., followed by the vaguely defined Archaic (Early, Middle, Late), from 8000 to 900 b.c. (Lohse et al. Reference Lohse, Awe, Griffith, Rosenswig and Valdez2006). This article adopts a different generalized time framework based on cultural developments specific to the neotropics (Iriarte et al. Reference Iriarte, Elliott, Yoshi Maezumi, Alves, Gonda, Robinson, de Souza, Watling and Handley2020), encompassing southern Mesoamerica and Central America, the isthmian Colombian land bridge region, and Greater Amazonia. By Greater Amazonia we refer to the Amazon basin, an area as large as the continental United States, as well as Venezuela and northern Colombia, including inter-Andean mountain valleys. The terminology used here includes:
(1) Late Pleistocene >12,000–10,000 b.c. This includes sites suspected to date well before Clovis technology until the end of the South American Fishtail tradition that corresponds temporally to the end of the Younger Dryas, around 9750 cal b.c. (Broecker et al. Reference Broecker, Denton, Lawrence Edwards, Cheng, Alley and Putnam2010).
(2) Terminal Pleistocene/Early Holocene, around 10,000–6000 b.c. This is a period when stemmed and barbed bifaces were produced at many locations across the neotropics (Figure 2) suggest a continuing reliance on hunting larger animals (Ranere and Cooke Reference Ranere and Cooke2021), as well as the earliest evidence that foragers are experimenting with precursors of domesticates and some of the earliest domesticates, including Cucurbita and Capsicum (Chiou and Hastorf Reference Chiou and Hastorf2014; Kistler et al. Reference Kistler, Newsom, Ryan, Clarke, Smith and Perry2015).
(3) Middle Holocene, 6000–2500 b.c., is the period of increasing use of domesticates across the neotropics, including maize, and the period for which we have the earliest evidence of maize consumption in the human diet (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020; Piperno Reference Piperno2011a; Tung et al. Reference Tung, Dillehay, Feranec and DeSantis2020; Figure 3). Bifacial stone tool production appears to end prior to the beginning of this time period in many regions of the neotropics (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019; Ranere and Cooke Reference Ranere and Cooke2021), though some areas, such as the Llanos de Moxos, Bolivia, never produced bifaces, and other sites have very limited evidence for some production during the Middle Holocene (Castillo Espitia and Aceituno Reference Castillo Espitia and Aceituno2006).
(4) The Late Holocene, 2500 b.c. to the present, is characterized by the emergence of agricultural and increasingly sedentary communities. Across the boundary from the Middle to Late Holocene we see the first evidence for more intensive maize farming and the importance of this grain as a dietary staple in the centuries leading up to the first permanent villages in the Maya lowlands before 1000 b.c. (Pohl et al. Reference Pohl, Pope, Jones, Jacob, Piperno, deFrance, Lentz, Gifford, Danforth and Kathryn Josserand1996). Much of our data from this period is related to human dietary changes and the trajectory of human societies towards sedentary agricultural societies.

Figure 2. Terminal Pleistocene and Early Holocene stemmed and barbed points from lower Central America, northern Colombia, northwest Honduras, coastal Nicaragua, and Belize (not to scale). (a–d) Points from Finca Guardiria, Costa Rica; (e) point base from La Fábrica, Costa Rica (Messina Reference Messina2016); (f) point from La Ribera, Costa Rica (Messina Reference Messina2016); (g) point from Los Camachos, Costa Rica (Messina Reference Messina2016); (h–j) points from La Mula Central, Panama; (k) point from the Middle Magdalena Valley, Colombia (López Reference López2004); (l) point from El Mirador site, Palestina, Caldas, Colombia (Herrera Reference Herrera2019); (m) point from El Gigante, Honduras (Scheffler Reference Scheffler2008); (n) point from Ometepe Island, Nicaragua; (o) Lowe point from Tzibte Yux, Belize (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019); (p) Lowe point from Orange Walk, northern Belize (Lohse et al. Reference Lohse, Awe, Griffith, Rosenswig and Valdez2006); (q) Lowe point from Toledo, southern Belize (Prufer et. al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019); (r) unprovenienced Lowe point from western Belize; (s) Lowe point found in central Belize (Stemp et al. Reference Stemp, Awe, Prufer and Helmke2016); (t) Sawmill point from western Belize (Stemp and Awe Reference Stemp and Awe2013); (u) Sawmill point from northern Belize (Stemp et al. Reference Stemp, Awe, Prufer and Helmke2016); (v) Sawmill point from western Belize (Stemp and Awe Reference Stemp and Awe2013); (w–x) Sawmill-like points from western Belize (Lohse et al. Reference Lohse, Awe, Griffith, Rosenswig and Valdez2006; Stemp et al. Reference Stemp, Awe, Prufer and Helmke2016); (y) provisional Ya'axché point found in the Bladen Nature Reserve, southern Belize (Stemp et al. Reference Stemp, Awe, Prufer and Helmke2016); (z) provisional Allspice point from central Belize (Stemp et al. Reference Stemp, Awe, Prufer and Helmke2016). Figures 2a–2m courtesy of A. Ranere (Ranere and Cooke Reference Ranere and Cooke2021); Figure 2n courtesy of Museo El Ceibo, Nicaragua, Hector Neff, C. Brown, and M. Ghitis. Figures 2o–2z photographed by Prufer, with permission from the Belize Institute of Archaeology.

Figure 3. Timeline showing earliest presence of domesticates in four regions of the neotropics in varying forms (microbotanical, macrobotanical), as well as timing of early maize consumption and adoption as a staple. See text for specific dates and references. Top and bottom bars are timing of Late Pleistocene Clovis and Fishtail lithic traditions in the neotropics and the timing of Early Holocene stemmed and barbed lithics found from Greater Amazonia to southern Mexico. Image by the authors.
EARLY HUMANS IN THE MESOAMERICAN NEOTROPICS (>10,000 b.c.)
Conservatively, it is now estimated that the initial New World colonists arrived in Central America by 12,500 cal b.c. (Waters Reference Waters2019; Williams and Madsen Reference Williams and Madsen2020). There they encountered a very different, and far less tropical, environment than today. At the time, the landscape was comprised of “heterogeneous, even patchy, vegetation across small distance scales; and stretches of forest alongside water courses in regions where forests were significantly reduced” (Piperno Reference Piperno2006:286). Pollen and macrofossil plant data suggest that the composition of forests certainly included tropical trees, but the extent of broadleaf forests was less than in the modern climate regime (Piperno and Pearsall Reference Piperno and Pearsall1998). Vegetation was more diverse than simple Pleistocene grassland/Holocene forest dichotomies would suggest (Piperno Reference Piperno, Bush, Flenley and Gosling2011b). Large tracts of the neotropics were forested, including across Greater Amazonia (Bush Reference Bush2017; Bush and de Oliveira Reference Bush and de Oliveira2006). Confronted with a greater diversity of large mammals and a wider range of riparian forest and grasslands, humans initially would have adapted to ecosystems that were similar to today, in that they were composed of broad-leaf tropical and dry highland forests and savannas (Hammond Reference Hammond and Harris1980; Iriarte et al. Reference Iriarte, Elliott, Yoshi Maezumi, Alves, Gonda, Robinson, de Souza, Watling and Handley2020; Magalhães et al. Reference Magalhães, Lima, Santos, Maia, Schmidt, Barbosa and da Fonseca2019; Morcote-Ríos et al. Reference Morcote-Ríos, Aceituno, Iriarte, Robinson and Chaparro-Cárdenas2021), but where the distribution of these landscapes was different compared with today.
Those migrants spread rapidly along the Pacific and Caribbean coasts, reaching southern Chile before 11,000 cal b.c. (Braje et al. Reference Braje, Dillehay, Erlandson, Klein and Rick2017; Dillehay et al. Reference Dillehay, Goodbred, Pino, Sánchez, Tham, Adovasio, Collins, Netherly, Hastorf, Chiou, Piperno, Rey and Velchoff2017). In the northern Yucatan peninsula, a near-complete human skeleton was found with extinct fauna in a submerged cave (Chatters et al. Reference Chatters, Kennett, Asmerom, Kemp, Polyak, Blank, Beddows, Reinhardt, Arroyo-Cabrales, Bolnick, Malhi, Culleton, Erreguerena, Rissolo, Morell-Hart and Stafford2014). The likely minimum age of the individual is approximately 10,000 b.c., based on Uranium-series dates of small calcite florets that had precipitated on bone before the skeleton was submerged by rising sea- and groundwater levels. Those dates are supported by radiocarbon dates and an abundance of Pleistocene faunal remains also found in the submerged chamber.
In southern Central America and across the isthmian region only a handful of stratified sites have Late Pleistocene occupations (Ranere and Cooke Reference Ranere and Cooke2021), and much of what we know comes from Clovis-like and Fishtail projectile points recovered from undated contexts in Panama (Ranere and Cooke Reference Ranere, Cooke, Bonnichsen and Turnmire1991) and Costa Rica (Snarskis Reference Snarskis1979; Swauger and Mayer-Oakes Reference Swauger and Mayer-Oakes1952). In North America, Clovis culture dates to 11,550–10,800 cal b.c. (Haynes et al. Reference Haynes, Anderson, Reid Ferring, Fiedel, Grayson, Vance Haynes, Holliday, Huckell, Kornfeld, Meltzer, Morrow, Surovell, Waguespack, Wigand and Yohe2007; Waters and Stafford Reference Waters and Stafford2007), though more recently constrained to 11,100–10,800 cal b.c. (Waters et al. Reference Waters, Stafford and Carlson2020), ending at the onset of the Younger Dryas. In South America, the stemmed Fishtail lithic tradition dates from 10,950–9950 cal b.c. (Politis and Prates Reference Politis, Prates, Harvati, Jäger and Reyes-Centeno2018; Suárez Reference Suárez2019; Waters et al. Reference Waters, Amorosi and Stafford2015). The Fishtail tradition spans the Younger Dryas and overlaps with Clovis by only 150 years. Evidence for both traditions is found across the isthmian region, including the iconic surface collections at Madden Lake, Panama (Ranere and Cooke Reference Ranere, Cooke, Bonnichsen and Turnmire1991; Sander Reference Sander1964).
Spatially, Clovis and Fishtail or Fishtail-like lithic traditions overlap in middle America from northern Venezuela to southern Chiapas, Mexico (Pearson Reference Pearson2017), with the largest numbers of examples found in Belize (Lohse et al. Reference Lohse, Awe, Griffith, Rosenswig and Valdez2006), Panama, and highland Guatemala (Brown Reference Brown1980; Coe Reference Coe1960; Gruhn et al. Reference Gruhn, Bryan and Nance1977; Ranere Reference Ranere, Morrow and Gnecco2006). This is not surprising given that the isthmian land bridge and surrounding tropical areas are a crossroads for technologies, sharing of knowledge, and contact between populations (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). The full distribution of undated Clovis and Fishtail points is well-summarized elsewhere (Cooke Reference Cooke1998; Lohse et al. Reference Lohse, Awe, Griffith, Rosenswig and Valdez2006; Ranere and Cooke Reference Ranere, Cooke, Bonnichsen and Turnmire1991; Stemp et al. Reference Stemp, Awe, Kathryn Brown, Harrison-Buck, Helmke, Wrobel and Yaeger2018). The only locations north of the Isthmus land bridge where a Clovis-like and a Fishtail-like projectile point were recovered from the same dated excavation context is from Los Grifos rock shelter in Chiapas, Mexico (Acosta Ochoa Reference Acosta Ochoa2010). In this rock shelter, the base of a putative Clovis and Fishtail points were found in contexts dating to 8428–7605 cal b.c.; however, these dates are inconsistent with the broad literature on the age of these two stone tool traditions (Waters Reference Waters2019). The largest-known Paleoindian site in lower Central America is Finca Guardiria, Costa Rica, a workshop spread over seven ha (Ranere and Cooke Reference Ranere and Cooke2021). At least 19 fluted projectiles, Fishtail points, and tens of thousands of other lithic artifacts have been recovered from surface collections and excavations into plowed contexts (Pearson Reference Pearson and Barton2004; Ranere Reference Ranere, Morrow and Gnecco2006). No contextual dates are associated with those excavations.
Genomic Evidence of Early Human Dispersal in the Neotropics
Recent paleogenomic studies help to track the region's earliest migrants. After humans arrived south of the North American ice sheets sometime between 17,450 and 14,550 b.c., they diversified into two main lineages, referred to as “Southern Native American” or “Ancestral A” (ANC-A) and “Northern Native American” or “Ancestral B” (ANC-B; Moreno-Mayar et al. Reference Moreno-Mayar, Vinner, de Barros Damgaard, de la Fuente, Chan, Spence, Allentoft, Vimala, Racimo, Pinotti, Rasmussen, Margaryan, Iraeta Orbegozo, Mylopotamitaki, Wooller, Bataille, Becerra-Valdivia, Chivall, Comeskey, Deviese, Grayson, George, Harry, Alexandersen, Primeau, Erlandson, Rodrigues-Carvalho, Reis, Bastos, Cybulski, Vullo, Morello, Vilar, Wells, Gregersen, Hansen, Lynnerup, Mirazon Lahr, Kjaer, Strauss, Alfonso-Durruty, Salas, Schroeder, Higham, Malhi, Rasic, Souza, Santos, Malaspinas, Sikora, Nielsen, Song, Meltzer and Willerslev2018; Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018; Raghavan et al. Reference Raghavan, Steinrucken, Harris, Schiffels, Rasmussen, DeGiorgio, Albrechtsen, Valdiosera, Avila-Arcos, Malaspinas, Eriksson, Moltke, Metspalu, Homburger, Wall, Cornejo, Moreno-Mayar, Korneliussen, Pierre, Rasmussen, Campos, de Barros Damgaard, Allentoft, Lindo, Metspalu, Rodriguez-Varela, Mansilla, Henrickson, Seguin-Orlando, Malmstrom, Stafford, Shringarpure, Moreno-Estrada, Karmin, Tambets, Bergstrom, Xue, Warmuth, Friend, Singarayer, Valdes, Balloux, Leboreiro, Vera, Rangel-Villalobos, Pettener, Luiselli, Davis, Heyer, Zollikofer, S, de Leon, Smith, Grimes, Pike, Deal, Fuller, Arriaza, Standen, Luz, Ricaut, Guidon, Osipova, Voevoda, Posukh, Balanovsky, Lavryashina, Bogunov, Khusnutdinova, Gubina, Balanovska, Fedorova, Litvinov, Malyarchuk, Derenko, Mosher, Archer, Cybulski, Petzelt, Mitchell, Worl, Norman, Parham, Kemp, Kivisild, Tyler-Smith, Sandhu, Crawford, Villems, Smith, Waters, Goebel, Johnson, Malhi, Jakobsson, Meltzer, Manica, Durbin, Bustamante, Song, Nielsen and Willerslev2015). A key component of identifying this branching was the analysis of a 10,950–10,750 cal b.c. child skeleton from the Anzick site in Montana (Rasmussen et al. Reference Rasmussen, Anzick, Waters, Skoglund, DeGiorgio, Stafford, Rasmussen, Moltke, Albrechtsen, Doyle, David Poznik, Gudmundsdottir, Yadav, Malaspinas, White, Allentoft, Cornejo, Tambets, Eriksson, Heintzman, Karmin, Korneliussen, Meltzer, Pierre, Stenderup, Saag, Warmuth, Lopes, Malhi, Brunak, Sicheritz-Ponten, Barnes, Collins, Orlando, Balloux, Manica, Gupta, Metspalu, Bustamante, Jakobsson, Nielsen and Willerslev2014). Anzick is archaeologically associated with Clovis culture, initially defined by distinctive stone tools found in close association with Pleistocene fauna at the Blackwater Draw site in New Mexico (Haynes Reference Haynes1995). Anzick is also on the ANC-A lineage, which is strongly represented in both early Central America (see below for descriptions of these skeletons) and South America (Nakatsuka et al. Reference Nakatsuka, Lazaridis, Barbieri, Skoglund, Rohland, Mallick, Posth, Harkins-Kinkaid, Ferry, Harney, Michel, Stewardson, Novak-Forst, Capriles, Durruty, Álvarez, Beresford-Jones, Burger, Cadwallader, Fujita, Isla, Lau, Aguirre, LeBlanc, Maldonado, Meddens, Messineo, Culleton, Harper, Quilter, Politis, Rademaker, Reindel, Rivera, Salazar, Sandoval, Santoro, Scheifler, Standen, Barreto, Espinoza, Tomasto-Cagigao, Valverde, Kennett, Cooper, Krause, Haak, Llamas, Reich and Fehren-Schmitz2020; Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018), as well as an individual from San Nicolas Island, California from around 2950 cal b.c. (Scheib et al. Reference Scheib, Li, Desai, Link, Kendall, Dewar, Griffith, Mörseburg, Johnson, Potter, Kerr, Endicott, Lindo, Haber, Xue, Tyler-Smith, Sandhu, Lorenz, Randall, Faltyskova, Pagani, Danecek, O'Connell, Martz, Boraas, Byrd, Leventhal, Cambra, Williamson, Lesage, Holguin, Soto, Rosas, Metspalu, Stock, Manica, Scally, Wegmann, Malhi and Kivisild2018). The Northern Native American (ANC-B) lineage is considered to be more closely associated with populations in eastern North America (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018).
The Anzick child skeleton is ancestral to the oldest known genomes in Central and South Americans from Chile, Brazil, and Belize, all of which date to between 8000 and 7000 cal b.c. These ancient people were not the only source for later South American populations. Other ancient individuals derive from lineages without specific affinity to the Clovis-associated genome, suggesting some degree of population replacement that began by 7000 cal b.c. and that was followed by substantial population continuity in multiple regions (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018).
THE TERMINAL PLEISTOCENE, EARLY HOLOCENE, AND MIDDLE HOLOCENE (10,000–2500 CAL b.c.)
Following the end of the Clovis and Fishtail cultural traditions, a clearer picture emerges of human presence in southern Mesoamerica and surrounding regions. By 9000 cal b.c., conditions were becoming wetter (Haug et al. Reference Haug, Hughen, Sigman, Peterson and Röhl2001; Winter et al. Reference Winter, Zanchettin, Lachniet, Vieten, Pausata, Ljungqvist, Cheng, Lawrence Edwards, Miller, Rubinetti, Rubino and Taricco2020) and warmer (Hodell et al. Reference Hodell, Anselmetti, Ariztegui, Brenner, Curtis, Gilli, Grzesik, Guilderson, Muller, Bush, Correa-Metrio, Escobar and Kutterolf2008). In the Peten, Guatemala, there is evidence that closed-canopy forests were undergoing anthropogenic burning by 9000 cal b.c. (Anderson and Wahl Reference Anderson and Wahl2016), with mixed herbaceous and woody plants being represented in charcoal records. Burning peaks are evident much later, between 6000 and 4000 cal b.c. (Schüpbach et al. Reference Schüpbach, Kirchgeorg, Colombaroli, Beffa, Radaelli, Kehrwald and Barbante2015), during the Holocene Thermal Maximum, arguably the warmest and wettest period of the Holocene (Renssen et al. Reference Renssen, Seppä, Heiri, Roche, Goosse and Fichefet2009). From multiple studies there is evidence that after 8500 cal b.c. the abundance of higher-ranked plant and animal resources declined, as rainforest overtook many Pleistocene open areas where game would have fed on scrub and grasses (Piperno and Pearsall Reference Piperno and Pearsall1998).
In lower Central America and the isthmian land bridge region, where tropical vegetation makes detection of early sites difficult, at least two dozen sites have components dating to the Terminal Pleistocene and Early Holocene, though there are few temporally diagnostic bifacial tools that have been recovered from dated stratigraphic contexts (Cooke Reference Cooke1998; Ranere and López Reference Ranere and López2007). In a new regional synthesis of the isthmian region, Ranere and Cooke (Reference Ranere and Cooke2021) note that some Terminal Pleistocene and Early Holocene sites north and south of the lower isthmian land bridge contain similar stemmed, notched, and barbed bifacial points (Figures 2a–2j). They also note that in well-dated excavation contexts, bifacial thinning flakes are absent from all contexts younger than 5950 cal b.c. in the Isthmo-Colombian region (Aceituno and Loaiza Reference Aceituno and Loaiza2014; Ranere and Cooke Reference Ranere and Cooke2021). Across the neotropics there is a diversity of early lithic traditions that have been better-studied in the isthmian region and Greater Amazonia, where more stratified sites have been identified. In the latter, many sites do not show a transition from bifacial to unifacial industries, but from the very earliest records, starting around 11,000 cal b.c., they show a range of unifacial lithic industries associated with a diversity of environments (Dillehay Reference Dillehay2000; Dillehay et al. Reference Dillehay, Goodbred, Pino, Sánchez, Tham, Adovasio, Collins, Netherly, Hastorf, Chiou, Piperno, Rey and Velchoff2017). Across the neotropics unifacial technologies can be associated with reliance on plants, small terrestrial animals, and riverine resources (Iriarte et al. Reference Iriarte, Elliott, Yoshi Maezumi, Alves, Gonda, Robinson, de Souza, Watling and Handley2020; Morcote-Ríos et al. Reference Morcote-Ríos, Aceituno, Iriarte, Robinson and Chaparro-Cárdenas2021; Roosevelt et al. Reference Roosevelt, da Costa, Lopes Machado, Michab, Mercier, Valladas, Feathers, Barnett, da Silveira, Henderson, Silva, Chernoff, Reese, Holman, Toth and Schick1996).
In Greater Amazonia a variety of stemmed and sometime barbed bifacial tools date to the Terminal Pleistocene and Early Holocene (Dillehay et al. Reference Dillehay, Calderón, Politis and Beltrão1992), suggesting that northwest South America was a region of technological innovation (Pearson Reference Pearson2017). In Ecuador, El Inga and several related sites have Terminal Pleistocene and Early Holocene dates (mostly with large errors), and several varieties of long-stemmed bifacial points, some barbed (Constantine Reference Constantine2013). Across the continent, the El Jobo tradition of western Venezuela is characterized by lanceolate rather than stemmed forms (Pearson Reference Pearson2017).
In the Middle Cauca region of west-central Colombia, 11 Early Holocene sites have been identified by AMS radiocarbon dating to have components dating to before 8000 cal b.c. (Dickau et al. Reference Dickau, Aceituno, Loaiza, López, Cano, Herrera, Restrepo and Ranere2015). Several are associated with stemmed and barbed points (Aceituno and Loaiza Reference Aceituno and Loaiza2014; Cano et al. Reference Cano, López and Méndez2021; Herrera Reference Herrera2019). These stemmed and barbed points (Figures 2k–2l) are similar to others recovered from the Magdalena River, also in northern Colombia, dating from between 10,000 and 7000 cal b.c. (López Reference López2021), from rock shelters in the Brazilian Amazon Orinoco River Valley, thought to date to around 7000 cal b.c. (Barse Reference Barse1990; Scaramelli and Scaramelli Reference Scaramelli and Scaramelli2017), and multiple locales near to the Caverna da Pedra Pintada rock shelter at Monte Alegre (Roosevelt et al. Reference Roosevelt, da Costa, Lopes Machado, Michab, Mercier, Valladas, Feathers, Barnett, da Silveira, Henderson, Silva, Chernoff, Reese, Holman, Toth and Schick1996). At Caverna da Pedra Pintada, a stemmed and barbed triangular point was recovered from well-dated contexts between 9800 and 9300 cal b.c. (recalibrated by the authors using IntCal20; Roosevelt et al. Reference Roosevelt, da Costa, Lopes Machado, Michab, Mercier, Valladas, Feathers, Barnett, da Silveira, Henderson, Silva, Chernoff, Reese, Holman, Toth and Schick1996).
Both the northern and southern Amazon are now arguably major global centers of domestication (Lombardo et al. Reference Lombardo, Iriarte, Hilbert, Ruiz-Pérez, Capriles and Veit2020), with an array of crops linked to wild progenitors native to the region (Manihot, Cucurbita, Capsicum) and Zea mays, which was initially domesticated in the Balsas River Valley of south central Mexico and then likely underwent secondary improvement of several critical traits that characterize modern maize (Kistler et al. Reference Kistler, Yoshi Maezumi, de Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Thomas, Gilbert, de Oliveira Freitas and Allaby2018, Reference Kistler, Thakar, VanDerwarker, George, Harper, Allaby, Hirth and Kennett2020).
Across the isthmian region there is compelling evidence for agroforestry, early horticulture, and cultivation of domesticates in the Early Holocene (Figure 3). In Panama, analysis of starch and phytoliths recovered from stone tools demonstrates plant-processing of early Zea mays and Poaceae spp. (grasses) by 5976–5915 cal b.c. (Dickau et al. Reference Dickau, Ranere and Cooke2007), and Cucurbita moschata by 6600 cal b.c. (Piperno Reference Piperno2011a).
The earliest Zea mays in South America has been found at the Las Vegas site in southwestern coastal Ecuador and dates to 6214–5843 cal b.c. (recalibrated by the authors using SHCal20, OxCal 4.4; Stothert et al. Reference Stothert, Piperno and Andres2002) in contexts also containing the earliest known domesticated Cucurbita, dating to 10,282–10,013 cal b.c. and 9894–9383 cal b.c. (recalibrated by the authors using SHCal20, OxCal 4.4; Piperno and Stothert Reference Piperno and Stothert2003). In the Andean and sub-Andean Medellín-Porce and Middle Cauca river valleys of northwest Colombia, analysis of pollen, starch grains, and phytoliths from tools indicates processing of Phaseolus spp. by 7181–6701 cal b.c., Manihot spp. by 6000 cal b.c., Dioscorea spp. (yams) by 9874–9372 cal b.c., and Zea mays, a cultigen, by 5882–5641 cal b.c. (uncalibrated dates calibrated by the authors using SHCal20 in OxCal 4.4; Aceituno and Loaiza Reference Aceituno and Loaiza2014, Reference Aceituno and Loaiza2018; Santos Vecino et al. Reference Santos Vecino, Marín and Salas2015). In the Llanos de Moxos, Bolivia, in the lowlands of the southwestern Amazon basin, Zea mays first appears at 5034–4475 cal b.c., Cucurbita at 8205–7682 cal b.c., and Manihot spp. by approximately 8350 cal b.c. (Lombardo et al. Reference Lombardo, Iriarte, Hilbert, Ruiz-Pérez, Capriles and Veit2020). West of the Amazon Basin at Huaca de Prieta, Peru, the earliest known use of Capsicum sp. dates to 8480–8700 cal b.c. (Chiou and Hastorf Reference Chiou and Hastorf2014).
North of the Isthmus, southeastern Mesoamerica contends with a similar combination of high precipitation and extensive tropical foliage that obscures early sites. Very few stratified archaeological sites are known. One exception is El Gigante, a large rock shelter in western Honduras on the periphery of Mesoamerica. There, stratified deposits containing remarkably well-preserved faunal and floral assemblages document Early Holocene occupations dating from 9060–5480 cal b.c. (Kennett et al. Reference Kennett, Thakar, VanDerwarker, Webster, Culleton, Harper, Kistler, Scheffler and Hirth2017). Three complete and four fragments of stemmed bifacial points were excavated from the earliest deposits (Figure 2m). The points are triangular in shape, have expanding stems and indented bases, and some have basal thinning or flutes on one face (Iceland and Hirth Reference Iceland, Hirth, Jon C., Borejsza and Joyce2021; Scheffler Reference Scheffler2008; Scheffler et al. Reference Scheffler, Hirth and Hasemann2012), and fit well with the broad regional pattern of stemmed and barbed triangular points dating to this period (Ranere and Cooke Reference Ranere and Cooke2021). Bifacial points are absent from strata younger than 7040 cal b.c. (Kennett et al. Reference Kennett, Thakar, VanDerwarker, Webster, Culleton, Harper, Kistler, Scheffler and Hirth2017) and the ratio of chert debitage to groundstone debitage drops significantly after this time (Scheffler Reference Scheffler2008).
Contexts at El Gigante dating from 9060–5660 cal b.c. also contain exceptionally well-preserved plant remains, including agave, wild avocado (Persea), hog plum (Spondias), mamey sapote (Pouteria), mesquite bean (Prosopis), and acorns (Quercus), while later contexts, from 5660–5490 cal b.c., also contain Soursop (Annona), hackberry (Celtis), squash (Cucurbita), and gourd (Lagenaria), suggesting an expansion of agroforestry practices during the Early Holocene (Kennett et al. Reference Kennett, Thakar, VanDerwarker, Webster, Culleton, Harper, Kistler, Scheffler and Hirth2017). The earlier contexts also contain higher abundance of large mammals (deer) than the 5660–5490 cal b.c. contexts, including an increase in the relative abundance of smaller game, including turtles, birds, snails, and crabs (Scheffler Reference Scheffler2008; Scheffler et al. Reference Scheffler, Hirth and Hasemann2012). Unfortunately, a hiatus in use of the rock shelter from 5480 to 2390 cal b.c. obscures much of the Middle Holocene.
In northern Mesoamerica (Mexico north of the Isthmus of Tehuantepec), drier conditions have facilitated the identification of early cave and surface sites (MacNeish et al. Reference MacNeish, Fowler and Cook1972b), including locales in central, west, and north Mexico (Gonzalez et al. Reference Gonzalez, Huddart, Israde-Alcántara, Domínguez-Vázquez, Bischoff and Felstead2015; Acosta Ochoa Reference Acosta Ochoa and Nichols2012; Sanchez and Carpenter Reference Sanchez, Carpenter, Bousman and Vierra2012). Among the best-known of these is the expansive effort by Richard MacNeish in the Tehuacan Valley. At Coxcatlán rock shelter, the earliest cultural contexts were initially thought to date to significantly before 7400 cal b.c. (MacNeish et al. Reference MacNeish, Johnson and Nelken-Terner1967, Reference MacNeish, Wittfogel, Woodbury and Johnson1972a, Reference MacNeish, Fowler and Cook1972b). More recent work suggests that the earliest contexts may not be older than 8000 cal b.c. (Smith Reference Smith2005) and that almost 50 percent of the radiocarbon dates from preceramic contexts can be rejected as being out of sequence. The early lithic Lerma assemblage at Coxcatlán has been linked to similar assemblages in Tamaulipas, Mexico, and artifacts from west Mexico have been opined to have originated in northeast Mexico (MacNeish and Nelken-Terner Reference MacNeish and Nelken-Terner1983). In the better-dated Coxcatlán phase there are some stemmed and barbed points dating to 5000–4000 cal b.c. (MacNeish et al. Reference MacNeish, Fowler and Cook1972b), but those are generally younger than those from the neotropics, and the dates with significant errors are imprecise.
Early humans in northern Mesoamerica were also deeply engaged in managing plant resources throughout the Holocene, and a wide variety of early domesticates have been identified in well-dated contexts across Mexico (Piperno Reference Piperno2011a). At Guilá Naquitz cave, domesticated Cucurbita pepo has been identified by 8085–7955 cal b.c. (Smith Reference Smith1997). Manioc appears much later in Mexico, with the earliest Manihot sp. pollen from tropical Tabasco in contexts estimated to be 4600 cal b.c. (Pope Reference Pope2001).
Maize, the most impactful crop in the New World, was first domesticated in the Balsas River Valley of western Mexico, by around 6900 b.c., in what was likely a single event (Matsuoka et al. Reference Matsuoka, Vigouroux, Goodman, Sanchez G., Buckler and Doebley2002) from lowland teosinte (Zea mays ssp. parviglumis), with contributions from highland teosinte (Zea mays ssp. Mexicana). The earliest evidence for maize comes from starch grains extracted from food-processing tools at the Xihuatoxtla shelter in the Central Balsas dated to 7010 cal b.c. (Piperno et al. Reference Piperno, Ranere, Holst, Iriarte and Dickau2009; Ranere et al. Reference Ranere, Piperno, Holst, Dickau and Iriarte2009). However, 4300 cal b.c. maize cobs from Guilá Naquitz were still small, with only two to four rows of seeds (Piperno and Flannery Reference Piperno, and Kent and Flannery2001). Maize from San Marcos cave, Tehuacán, Mexico, dating to 3350–3020 cal b.c. (Benz and Long Reference Benz and Long2000; Ramos-Madrigal et al. Reference Ramos-Madrigal, Smith, Víctor Moreno-Mayar, Gopalakrishnan, Ross-Ibarra, Thomas, Gilbert and Wales2016; Vallebueno-Estrada et al. Reference Vallebueno-Estrada, Rodríguez-Arévalo, Rougon-Cardoso, González, Cook, Montiel and Vielle-Calzada2016), was only partially domesticated for the full suite of characteristics setting a domesticated species apart from its wild counterpart (Kistler et al. Reference Kistler, Yoshi Maezumi, de Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Thomas, Gilbert, de Oliveira Freitas and Allaby2018). Very gradual changes in cob size in the Tehuacan Valley likely reflect introgression with teosinte in the natural range of parviglumis and Mexicana (Kennett et al. Reference Kennett, Thakar, VanDerwarker, Webster, Culleton, Harper, Kistler, Scheffler and Hirth2017).
Partially domesticated maize quickly radiated southward into Greater Amazonia by 6200 b.c. (Stothert et al. Reference Stothert, Piperno and Andres2002), and was followed by successive waves adding to crop diversity (Freitas et al. Reference Freitas, Bendel, Allaby and Brown2003). Early varieties underwent secondary improvements in South America (Kistler et al. Reference Kistler, Yoshi Maezumi, de Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Thomas, Gilbert, de Oliveira Freitas and Allaby2018), and a subsequent wave of maize introduced additional varieties that hybridized with the first wave. Once maize was outside the native range of introgression from parviglumis and Mexicana, selection for increased cob and seed size was enhanced and the result was the development of more productive staple grain varieties. It is not known when these improved varieties were reintroduced into Mesoamerica, but the improved genomes are present at El Gigante, Honduras by 300 cal b.c. (Kistler et al. Reference Kistler, Thakar, VanDerwarker, George, Harper, Allaby, Hirth and Kennett2020) and likely contributed to the development of more productive staple grain varieties in Central America by 2750 cal b.c. (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020).
In western Mesoamerica on the Pacific coast of Chiapas, maize phytoliths were found in sediment cores dating to 4550 cal b.c. (Kennett et al. Reference Kennett, Piperno, Jones, Neff, Voorhies, Walsh and Culleton2010). It is unknown if these were partially domesticated. Manihot, which was originally domesticated in Bolivia (Lombardo et al. Reference Lombardo, Iriarte, Hilbert, Ruiz-Pérez, Capriles and Veit2020), first appears in Tabasco at 4600 cal b.c. (Pope Reference Pope2001) and then, together with Zea mays and Capsicum sp., in northern Belize by 4550 cal b.c. (Rosenswig et al. Reference Rosenswig, Pearsall, Masson, Culleton and Kennett2014), and was becoming more widespread by 3500 cal b.c. (Pohl et al. Reference Pohl, Pope, Jones, Jacob, Piperno, deFrance, Lentz, Gifford, Danforth and Kathryn Josserand1996). Chili peppers (Capsicum spp.) were domesticated in Peru by 8350 cal b.c. (based on dated seeds), and in Mexico by approximately 3650 cal b.c. (Chiou and Hastorf Reference Chiou and Hastorf2014; Kraft et al. Reference Kraft, Brown, Nabhan, Luedeling, de J, Ruiz, Coppens d'Eeckenbrugge, Hijmans and Gepts2014); of the four species of Capsicum cultivated across Mesoamerica, three were first domesticated in South America (Perry and Flannery Reference Perry and Flannery2007). It is not clear if early southern Mesoamerican Capsicum starch grains (Rosenswig et al. Reference Rosenswig, Pearsall, Masson, Culleton and Kennett2014) derive from the South American or Mexican domesticates.
THE LATE HOLOCENE IN SOUTHEASTERN MESOAMERICA
Not surprisingly, there are more data for the period just before the rise of agricultural communities, suggesting that populations were larger and distributed across more and varied landscapes (Kennett et al. Reference Kennett, Piperno, Jones, Neff, Voorhies, Walsh and Culleton2010; Rosenswig et al. Reference Rosenswig, VanDerwarker, Culleton and Kennett2015; Voorhies et al. Reference Voorhies, Kennett, Jones and Wake2002). Semimobile farmers may have been practicing slash-and-burn maize agriculture in coastal Chiapas by 2750 cal b.c. (Kennett et al. Reference Kennett, Piperno, Jones, Neff, Voorhies, Walsh and Culleton2010), though full-scale maize agriculture may not have been adopted there before 1000 cal b.c. (Rosenswig et al. Reference Rosenswig, VanDerwarker, Culleton and Kennett2015), even though sedentary agricultural communities are present by 1500 cal b.c. An isotope study drawing on data from Mesoamerica and South America suggested that some individuals dating between 1450 and 900 cal b.c. do not show a clear signal of high maize consumption, while others, indirectly dated before 2000 cal b.c., were eating some C4 enriched foods (maize, CAM plants) or marine resources (Smalley and Blake Reference Smalley and Blake2003). The earliest maize cobs in neotropical Mesoamerica come from El Gigante, Honduras and date to after 2390 cal b.c. (Kennett et al. Reference Kennett, Thakar, VanDerwarker, Webster, Culleton, Harper, Kistler, Scheffler and Hirth2017); and cobs dated between 300 cal b.c. and 100 cal a.d. contain the signal of maize varieties that underwent secondary improvements in South America, and their reintroduction northward likely contributed to the development of more productive staple maize varieties (Kistler et al. Reference Kistler, Thakar, VanDerwarker, George, Harper, Allaby, Hirth and Kennett2020).
A wide range of cultural changes occurred during the transition to Late Holocene, with a movement towards intensive cultivation and reliance on plants as a source of food. Populations were likely a mix of small, seasonally residential groups relying on mixed hunting and foraging strategies, as well as sedentary horticulturalists that progressively intensified farming early. In Honduras, Costa Rica, Panama, and Brazil, distinctive ceramic traditions are present between 3000 and 2000 b.c., suggesting that sedentary agricultural communities were established by the start of the Late Holocene (Hoopes Reference Hoopes1994; Iizuka Reference Iizuka2017; Joyce and Henderson Reference Joyce and Henderson2001; Roosevelt et al. Reference Roosevelt, Housley, Da Silveira, Maranca and Johnson1991). Thus, social changes were likely mediated by subsistence changes. While the Middle Holocene from 8000 to 3500 cal b.c. is generally considered to have been wetter and warmer, it is followed by a drier interval lasting until 2200 cal b.c. (Mueller et al. Reference Mueller, Islebe, Hillesheim, Grzesik, Anselmetti, Ariztegui, Brenner, Curtis, Hodell and Venz2009; Winter et al. Reference Winter, Zanchettin, Lachniet, Vieten, Pausata, Ljungqvist, Cheng, Lawrence Edwards, Miller, Rubinetti, Rubino and Taricco2020), during which time maize becomes a more prominent crop in the paleoecological record and as a component in the human diet (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020; Piperno and Pearsall Reference Piperno and Pearsall1998). This scenario suggests that the long transition to agriculture spanned several phases of significant climate and environmental change, leading to the formation of cultural traits that can be considered Maya.
In the following section, we present data from southern Belize and excavations at two rock shelter sites that have excellent preservation of organic material. We describe the regional setting, the archaeological excavations, and results of laboratory analysis in relation to the trends outlined above.
SOUTHERN BELIZE AND THE MAYA MOUNTAINS ROCK SHELTERS
Southern Belize (Figure 1) is a geographically distinct region in Central America, with a diverse set of geological and biotic resources and the backdrop for a history of human occupation spanning the Late Pleistocence and Holocene (Prufer and Kennett Reference Prufer, Kennett, Houk, Aroyo and Powis2020). In addition to sites dating to the dawn of human impact in the region discussed here, this region was home to over 30 independent polities and a vastly productive agricultural landscape during the Classic Period (250–1000 cal a.d.), and has been continuously occupied by Maya people and their ancestors from the Pleistocene to the present. Physically, the region is circumscribed by the Maya Mountains to the west, a series of swampy bajos to the south, along the Temash and Mojo rivers, the Caribbean Sea to the east, and inhospitable pine barrens to the north. It is among the wettest places in Central America, receiving 3,500–4,000 mm of rainfall annually (Asmerom et al. Reference Asmerom, Baldini, Prufer, Polyak, Ridley, Aquino, Baldini, Breitenbach, Macpherson and Kennett2020; Ridley et al. Reference Ridley, Asmerom, Baldini, Breitenbach, Aquino, Prufer, Culleton, Polyak, Lechleitner, Kennett, Zhang, Marwan, Macpherson, Baldini, Xiao, Peterkin, Awe and Haug2015), more than double the precipitation of the Peten and seven times as much as the northern Yucatan Peninsula (Douglas et al. Reference Douglas, Pagani, Canuto, Brenner, Hodell, Eglinton and Curtis2015). It is also a seasonal desert (Haug et al. Reference Haug, Hughen, Sigman, Peterson and Röhl2001), where for several months each year there is little to no rainfall and evaporation exceeds precipitation.
The Maya Mountains were formed by Devonian subaerial volcanos characterized by lava flows and pyroclastic and volcanoclastic activity, some locally altered hydrothermally, and by the Pennsylvanian-Permian Santa Rosa Group of argillaceous and arenaceous sediments and carbonates. The eastern slope is aproned by Tertiary and Cretaceous limestones of the Coban Formation (Bateson and Hall Reference Bateson and Hall1977). Combined with high precipitation during the Quaternary, the result is a hydrologically carved network of caves and cockpit karst overlaying earlier volcanics. The interior valleys, where people lived, have a unique geological history compared to the rest of the Maya lowlands. The upper reaches of tributaries contain volcanic and metamorphic material eroding off the spine of the mountains that are ubiquitous in geological float in streams and rivers. Soil pedogenesis in the alluvial valleys incorporates carbonate and volcanic materials, making them a rich agricultural landscape surrounded by near-vertical mountains, and hosting diverse biotic communities and economic resources that would have been of interest to farming and foraging communities (Dunham and Prufer Reference Dunham, Prufer, Laporte and Escobedo1998).
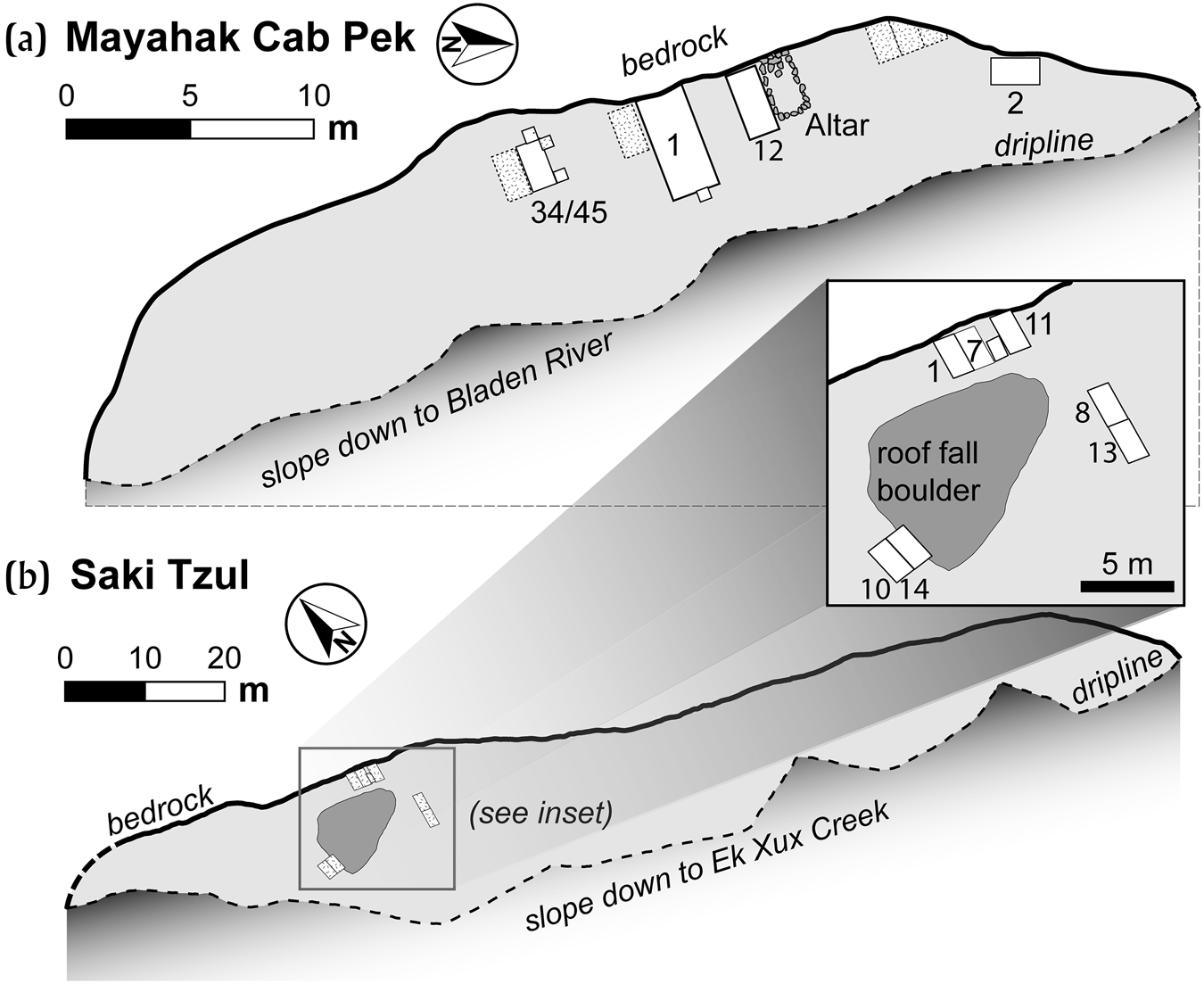
Mayahak Cab Pek (MHCP) and Saki Tzul (ST) are rock shelters located in an interior valley of the Maya Mountains (Figure 1b) in the Bladen Nature Reserve, a protected wilderness area where there has been minimal modern human disturbance of archaeological sites. Our work from 2014 to 2019 has demonstrated that the cultural use of these rock shelters began prior to 10,500 cal b.c. and continued through 1000 cal a.d. (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019), and even continues today, as debris from the occasional hunter or chiclero can be found. The two sites were first documented in 1997–1998 (Prufer Reference Prufer2002). At that time, shallow excavations at MHCP produced burials with excellent preservation of human and faunal remains (Prufer Reference Prufer2002; Saul et al. Reference Saul, Prufer, Saul, Prufer and Brady2005), but did not identify pre-agricultural contexts. Though the two rock shelters are located 1.4 km apart, they have similar stratigraphic sequences and contain similar assemblages of artifacts and biological remains. Both have dry sediments and large overhangs, reflecting that little if any direct rainfall affects cultural deposits. This circumstance also helps to explain the excellent preservation of unburned bone and other organic materials and only minor root presence, primarily close to the driplines. Additional information on the mortuary context for the 1998 burials at MHCP has been published previously (Saul et al. Reference Saul, Prufer, Saul, Prufer and Brady2005), though we have revised the chronological sequences of those burials (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020) after direct-dating of the skeletons. Below, we briefly describe each rock shelter, the stratigraphic sequences, and the assemblages recovered from excavations.
MHCP is an east-facing shelter under a limestone face, 20 m high, with easily discernable bedding planes, many marked by caramel-colored chert lenses and cobbles (Figure 4a). The ground surface of the rock shelter runs roughly south-southeast to north-northwest, with a slope gradient that varies between 6 and 14 degrees, steepening towards the south. The shelter surface is located approximately 20 meters above the present-day stream bed of an unnamed tributary of the Bladen branch of the Monkey River. The rock shelter overhang offers a substantial amount of shelter to the surface below, which is extremely dry and dusty. The dripline extends to approximately 8 meters from the cliff face. The floor of the rock shelter shows no signs of erosion from water flow or deflation. The wall shows a few indicators of travertine growth, likely from water seepage. The overall area of the floor is approximately 160 square meters. Disturbance from plant growth is not significant, since the ground surface is sheltered from rain. The east-facing aspect and high-canopy forest cover limits direct sunlight.
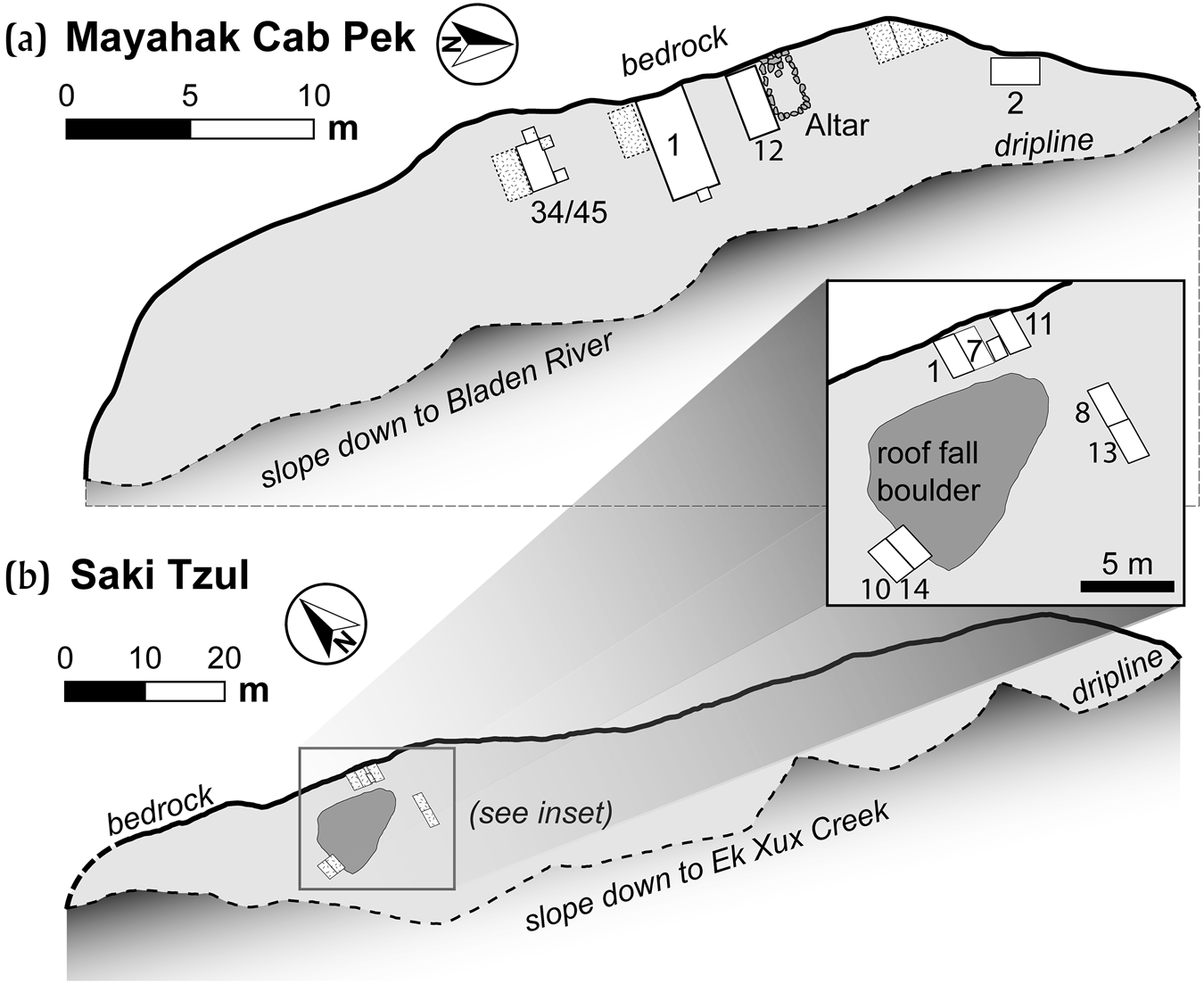
Figure 4. Plan views of (a) MHCP and (b) ST rock shelters and locations of excavation units. Image by Prufer, Thomas Harper, and Erin Ray, after Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020.
Saki Tzul (ST) rock shelter is located in the Ek Xux Valley of the Bladen Nature Reserve, along and 70 m above the Ek Xux Creek, less than 300 m from the Classic period Maya center Ek Xux. ST is located on the eastern side of the valley, 1.4 km north from MHCP, and is a starkly white, limestone outcrop, and one of the most impressive geological features on the local landscape. The massive limestone overhang is approximately 80 meters high, 145 meters long, and shelters an area between 8 and 15 meters wide, covering 1700 square meters (Figure 4b). The rock shelter runs along a generally east–west axis, with differences in the elevation of the ground surface, with the most level area being the location of our investigations. With a south-facing exposure, the rock shelter receives abundant sunlight throughout the day. We have noted that during large storms some limited rain blows in from heavy winds, resulting in enough moisture for some scrubby vegetation to grow within the dripline, but not enough to generate fluvial effects. Eight excavations were conducted between 2016 and 2018, in only a small area of the rock shelter, around a large breakdown boulder that fell from the overhanging cliff roof prior to human use (Figure 4b). Multiple excavation units reveal a similar stratigraphic sequence to MHCP, also spanning the past 12,500 years.
Across all units at both rock shelters, the stratigraphy is characterized by a sequence of cultural midden and mortuary deposits approximately 3 meters deep (Figures 5a–5b). The ceramic-bearing upper portions of units in both rock shelters can be characterized generally as repeating two sub-stratigraphic soil units of varying depth, which include midden fill overlying dense concentrations of cobbles that likely represent occupation surfaces. This sequence of midden deposits and cobbles is repeated at least two or three times in the ceramic-bearing contexts. These deposits date after approximately 2000 cal b.c. and contain pottery fragments, flaked stone chert and other stone tools, and the remains of mammals, birds, and reptiles. Artifact density is high in the upper ceramic-bearing strata, with concentrations of burned and unburned animal bone, burned wood and charcoal, and disarticulated human remains. The cultural levels of the Classic Maya start just a few centimeters below a covering of modern windblown leaves, suggesting that sediments have aggraded little since the region was abandoned around 1000 cal a.d.

Figure 5. Composite profiles of (a) MHCP and (b) ST, showing location of burials. Image by Prufer, Thomas Harper, Erin Ray, and Asia Alsgaard, modified from Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020.
Two dense freshwater shell lenses occur just below these Classic period deposits. These contain a few pottery sherds, confined to the top of the shell midden. Jute (Pachychilus sp., a freshwater snail) concentrations comprise up to 50 percent of the matrix and are densest toward the contact with ceramic levels and continuing into aceramic levels. Almost all the jute shells appear to be spire-lopped, which has been suggested elsewhere as an indicator that jute was prepared and consumed as a food product (Halperin et al. Reference Halperin, Garza, Prufer and Brady2003; Hammond Reference Hammond1975). These levels are also characterized by dense, cobble-rich horizons, primarily consisting of sub-round to sub-angular limestone and porphyritic igneous clasts. Given the generally rounded nature of the clasts, they likely derived from river float and would have been transported to the rock shelter by humans.
The lowest stratigraphic units are organic-rich silt to silty loams and contain some debris from the limestone cliff outcrop, igneous flaked stone tools of local origin, several large chert bifaces (Lowe points), animal, riverine mollusks (Pachychilus spp.), and human remains. Sediments are poorly sorted and likely represent the natural accumulation of exogenic (silt) and endogenic (roof fall) with anthropogenic cultural midden materials. Artifact and faunal concentrations are moderate, and jute snail concentrations also drop to less than 10 percent of the matrix in some contexts. The size and the number of cobbles, both limestone and porphyritic igneous rocks and expedient tools, tend to increase with depth. The lowest cultural levels are silty. Within the silty matrix we found a smaller frequency of chert tools, but they were larger and have a high concentration of expedient blades, some of which are modified bifacial thinning flakes. The silty matrix directly above decaying limestone likely represents the first extensive human use of the rock shelters. These silty matrices terminate on bedrock in some units (likely roof fall breakdown) over reddish clays and decaying limestone. These basal cultural layers are uneven, with pockets of silts containing cultural materials and, in more than one case, a human mandible. Probes deep into these non-cultural contexts yielded primarily the remains of rodents and small birds, at least one example of Pleistocene megafauna, as well as small pieces of charcoal and possibly a few simple chert flakes. These date to the Late Pleistocene and are the subject of ongoing research. These contexts cannot be defined as anthropogenic with any certainty.
Chronology
AMS radiocarbon dates bracket our pre-agricultural contexts between 10,670 and 2050 cal b.c. Older dates are generally in excellent stratigraphic order, but numerous shallow burial features, fire pits, and other cultural intrusions over time have resulted in some reversals (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). There is no evidence for fluvial impacts, site deflation, or unconformities. In general, the Classic Maya period dates are not in sequence, as more intensive use of the rock shelter during that time is reflected in increased mixing within contexts. It is likely that the dense jute middens just below the Classic period contexts protected the earlier materials below. All radiocarbon dates were prepared at The Pennsylvania State University (PSU) Human Paleoecology and Isotope Geochemistry Lab and the University of New Mexico Center for Stable Isotopes, and processed either at the PSU-AMS facility or the University of California Irvine KECK-AMS facility. Charcoal was prepared using standard ABA methods (Kennett et al. Reference Kennett, Culleton, Voorhies and Southon2011), while bone collagen was prepared using both ultrafiltration and XAD amino acid separation (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). All dates have been calibrated using the IntCal20 (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Lawrence Edwards and Friedrich2020) terrestrial curve.
We developed three Poisson distribution age models at MHCP and ST that illustrate the pre-agricultural sequences for both sites (Figures 6a–6c). Poisson models take as an argument (k) the number of postulated events per unit length and for deposition sequences the event scale, usually in the 0.1–1.0 cm range. In order to generate an age-depth model, the dated events can be interpolated automatically (Ramsey Reference Ramsey2008). Stratigraphic depths were adjusted to reflect depths below the ground surface and to account for surface and basal slope in excavation units. We selected charcoal from unit fill and not from within intrusive features for analysis and inclusion of models. Bayesian Poisson depth models were produced in OxCal 4.4 using published parameters and a Poisson parameter ko value of 1 (depth-1), resulting in interpolation at every 1.0 cm. One AMS date from each age model was excluded as out of sequence. Model agreements are high and suggest that human use of the rock shelters begins by 10,600 cal b.c. (Figures 6a–6b) and continues throughout the Holocene.

Figure 6. Poisson depth-age models from three excavations units: (a) MHCP Unit 13(34/35); (b) MHCP Unit 1; and (c) ST Unit 13. Note: Dates for stratigraphic positions of Lowe-type bifaces recovered in 2019 (a) and 2017 (b). Model generated in Oxcal v4.4.2 (Ramsey Reference Ramsey2016) using the Intal2020 calibration curve (Reimer et al. Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey, Butzin, Cheng, Lawrence Edwards and Friedrich2020) Labels show AMS sample ID (14C age, error). Images by Prufer.
Second, we use a simple summed probability distribution (SPD) for all direct dates on human skeletons (Table 1) as an indicator of frequency and intensity of use of the two rock shelters as mortuary sites (Figure 7). SPDs are aggregates (sums) of calibrated radiocarbon dates within a defined chronological range. They are frequently a proxy for demographic change (Shennan et al. Reference Shennan, Downey, Timpson, Edinborough, Colledge, Kerig, Manning and Thomas2013); however, given the uncertainties in mobility patterns of foraging populations, we consider these a better indicator of the frequency or intensity of the use of the sites for mortuary activities. The SPD was calculated from calibrated dates using the package rcarbon (Crema and Bevan Reference Crema and Bevan2020). The distribution of these dates suggests episodic use of the rock shelters before 3800 cal b.c. Between 3800 and 2000 cal b.c., sustained mortuary activity increases. This rise may reflect a more persistent population in the region or increased regional attention on these sites as preferred locales for interring the dead. Gaps in the overall distribution of when individuals were buried may be a consequence of limited subsurface testing of less than 5 percent of ST and 20 percent of MHCP and are certainly reflected in the difficulty in dating these early burials. An additional 15 burials not included in this study have poor collagen preservation, but likely date to the Early and Middle Holocene based on stratigraphic position.
Table 1. Human skeletal remains and results of direct dating and isotopic analysis. After Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020.


Figure 7. Summed probability distribution for burials dating to 7600–1950 B.C. Graph by Prufer.
The Lithic Toolkit in the Bladen
The stone tool assemblages from MHCP and ST are large, and analysis is ongoing. Still, we can make several observations based on the existing chronological data. In 2017 we recovered two large bifaces from MHCP in levels dating between 8750 and 6500 cal b.c. (Figure 5b; Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). These bifaces show a suite of characteristics that place them into a category of points designated as Lowe (Stemp et al. Reference Stemp, Awe, Prufer and Helmke2016). Lowe points are large, stemmed, and barbed bifaces, with many examples showing unifacial beveling on alternate edges, longitudinal flake thinning on one side of the base only, concave-to-flat bases, and stem sides that range from straight to slightly contracting from the base to the neck. We also recovered a Lowe biface from excavations at Tzib te Yux rock shelter, located approximately 40 kilometers from the MHCP less than 2 km from the Maya center Uxbenka (Figure 1) in a stratigraphic context dated to 8275–7980 cal b.c. From that same site we excavated the distal end of a large biface with steep, unifacial beveling on alternate edges from a stratigraphic context dated to 10,450–10,085 cal b.c. (Figure 8; Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). Here we report the recovery of one additional Lowe point from MHCP in 2019. This point was recovered from contexts dating to 6910–6650 cal b.c. (Figures 6a and 8).

Figure 8. Lowe-type point recovered from MHCP in 2019 and dated to 6910–6650 cal. b.c. Drawing by Erin Ray for the Bladen Paleoindian and Archaic Archaeological Project; photograph by Prufer.
Additional chipped stone materials show a change in lithic technology at around 6000 cal b.c. Contexts dated earlier contain abundant large bifacial thinning flakes, many of them utilized as blades, based on edge damage, and some show evidence of microflaking. Most have prepared striking platforms, and a few show evidence for pointed platforms. Overall, the oldest levels have fewer chert artifacts, but those that are present are larger (Figure 8) and suggest a significant amount of cobble testing, as evidenced by many decortication flakes, as well many large, tested but abandoned cobbles where the cortex has been partially removed. Unifacial dome-shaped scrapers are also present in the earliest levels, some of which are quite large (approximately 10 cm in diameter).
In contexts younger than approximately 6000 cal b.c., we continue to see cortical flakes and cobble testing, but no longer see any evidence of bifacial thinning. Most of the chert tools are expedient, made from cherts of varying quality and including small and medium modified flakes, crude awls, hammerstones with battered edges, and the debitage from processing those expedient tools. Some expedient tools appear in all time periods and thus are not considered temporally diagnostic (Figures 9a–9d). These include end scrapers with cutting edges, and crude keel end scrapers, discoidal scrapers, and choppers, although large, dome-shaped scrapers only appear in Late Pleistocene through Early Holocene contexts.

Figure 9. Non-bifacial lithics. (a) Chert chopper, Early Holocene; (b) chert unifacial scraper, Early Holocene; (c–d) chert discoidal dome-shaped unifacial scrapers, both Late Pleistocene; (e) a variety of Early Holocene end-battered grinding stones and choppers, all porphyritic river cobbles. Photographs by Prufer and Erin Ray.
The other major component of the lithic assemblage are non-chert choppers and hammerstones, and what are likely crude grinding stones (Figure 9e). These tools are found in all levels and contexts. Many of these were worked by crude splitting and flaking, particularly of the volcanic rocks for expedient chopping and cutting tools, with little, if any, evidence of retouch. In the Bladen River, below MHCP, and in the Ek Xux Creek, below ST, cobbles are dominated by limestone and porphyritic igneous cobbles, suggesting that the latter are a source for expedient tools manufactured at or carried to the rock shelters.
Skeletal Population
Mortuary contexts at both rock shelters reflect similar burial practices and include both primary and secondary burials. Isolated elements were recovered from many stratigraphic contexts, and while some are the result of intrusive disturbance into earlier contexts, others are intentional deposits of individual skeletal elements or multiple elements. Burials in varying degrees of flexure were the most common and are found in all time periods. Less common are extended burials, several of which are face down.
In total, 52 inhumations have been described from both rock shelters (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020), not including isolated remains and individuals that we have not yet directly dated. Here we discuss only 27, reflecting those skeletons who are pre- or transitional maize agriculturalists and for whom we have been able to obtain direct dates (Table 1). That population consists of nine males, five females, and 13 for whom sex could not be determined, either because they were infants or young children, or because the remains were too fragmentary to determine sex. The age structure consists of 15 adults, nine infants or children, and three indeterminate individuals.
It is worth giving extra attention to three of the earliest skeletons, one from MHCP and two from ST (burials MHCP.14.1.6, ST.16.1.2, and ST.16.1.3; Table 1), for whom we obtained direct dates ranging between 7350 and 5350 cal b.c. and genome-wide data. Individual MHCP.14.1.6 is the remains of an older adult female as determined through osteological analysis and DNA. She likely died elsewhere, and her remains were then disarticulated and transported to MHCP and buried in a shallow pit. Much of what remains of the skeleton (<75 percent complete) has hundreds of shallow cut-marks, primarily on the axial skeleton and limbs. The shallow burial pit in which she was placed contained a higher density of chipped stone than the surrounding matrices (n = 71 artifacts), including utilized blades, flakes with sharpened (micro-flaked) edges, and bifacial thinning flakes. MHCP.14.1.6 belongs to the mitochondrial haplogroup D4h3a, a rare matriline which, among ancient individuals, has been identified so far only in two individuals from the North American northwest (Scheib et al. Reference Scheib, Li, Desai, Link, Kendall, Dewar, Griffith, Mörseburg, Johnson, Potter, Kerr, Endicott, Lindo, Haber, Xue, Tyler-Smith, Sandhu, Lorenz, Randall, Faltyskova, Pagani, Danecek, O'Connell, Martz, Boraas, Byrd, Leventhal, Cambra, Williamson, Lesage, Holguin, Soto, Rosas, Metspalu, Stock, Manica, Scally, Wegmann, Malhi and Kivisild2018), the 10,950–10,750 cal b.c. Anzick-1 individual from Montana (Rasmussen et al. Reference Rasmussen, Anzick, Waters, Skoglund, DeGiorgio, Stafford, Rasmussen, Moltke, Albrechtsen, Doyle, David Poznik, Gudmundsdottir, Yadav, Malaspinas, White, Allentoft, Cornejo, Tambets, Eriksson, Heintzman, Karmin, Korneliussen, Meltzer, Pierre, Stenderup, Saag, Warmuth, Lopes, Malhi, Brunak, Sicheritz-Ponten, Barnes, Collins, Orlando, Balloux, Manica, Gupta, Metspalu, Bustamante, Jakobsson, Nielsen and Willerslev2014), and four ancient individuals from Brazil and Chile (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018). This haplogroup has been hypothesized to be a marker of human dispersal along the Pacific coast, where it is found in modern populations (Perego et al. Reference Perego, Achilli, Angerhofer, Accetturo, Pala, Olivieri, Kashani, Ritchie, Scozzari, Kong, Myres, Salas, Semino, Bandelt, Woodward and Torroni2009). The initial diversification of the D4h3a haplogroup is estimated to have occurred around 14,000 b.c., which overlaps with the coalescence of haplogroups A2, B2, C1, and (importantly) D1 in the new world (Llamas et al. Reference Llamas, Fehren-Schmitz, Valverde, Soubrier, Mallick, Rohland, Nordenfelt, Valdiosera, Richards, Rohrlach, Romero, Espinoza, Cagigao, Jiménez, Makowski, Reyna, Lory, Torrez, Rivera, Burger, Ceruti, Reinhard, Spencer Wells, Politis, Santoro, Standen, Smith, Reich, Ho, Cooper and Haak2016). Since it is known now to be present in MHCP.14.1.6 and in ancient individuals from Brazil and Chile, it seems likely to have spread eastward along the Atlantic coast (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018), at least starting the neotropics. The early individuals from ST (ST.16.1.2 and ST.16.1.3) are both middle-adult males who were interred together in flexed positions within the same burial feature, the base of which is located almost 2 m below the modern ground surface. Both of these individuals belong to the mitochondrial haplogroup D1. The D1 and D4h3a haplogroups are both rare today in Mesoamerica. Both of the ST individuals also belong to the same Y-chromosome haplogroup Q1a2a1a1, the main subclade for all Native Americans and a predominant haplogroup (51–86 percent) throughout the region (Perez-Benedico et al. Reference Perez-Benedico, La Salvia, Zeng, Herrera, Garcia-Bertrand and Herrera2016).
We analyzed dietary isotopes of carbon and nitrogen on bone collagen for 25 skeletons and on bone apatite from a different subset of 25 individuals (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). All individuals dating prior to approximately 2750 cal b.c. (Table 1; Figure 10) show no evidence that they were consuming maize. These individuals derived both the carbohydrate and protein fractions of their diet from C3 plants and from animals consuming C3 plants. There is no evidence that they were relying to any measurable extent on marine resources in their diet. The data do not exclude the possibility that freshwater protein sources (i.e., snails or crabs) were a component of the diet. The bone collagen δ13C values for these individuals range between −21.6 and −20.3‰ (Mean = −20.8‰; SD = 0.3) and represent the isotopic values of protein, which are biased primarily towards meat consumption, but also reflect some contribution from vegetable protein (Froehle et al. Reference Froehle, Kellner and Schoeninger2012; Schoeninger and DeNiro Reference Schoeninger and DeNiro1984). The data are consistent with the diet of people living in a tropical rainforest and eating locally available plants and animals, where C4 resources are not found in any abundance (Wright Reference Wright2006). The δ15N values range from 6.4 to 9.9‰, which is also consistent with a population consuming C3 plants and terrestrial animals that are consuming C3 plants. The δ15N values are approximately 3–6‰ higher than published values for herbivores in the region (Wright Reference Wright2006). This figure is high, considering that the normal trophic discrimination of nitrogen is 1–2‰ (Robbins et al. Reference Robbins, Felicetti and Sponheimer2005). This suggests that people may have been consuming relatively more omnivores or carnivores, and potentially supplementing the consumption of mammals with freshwater fish or gastropods. Since carbon isotope enrichment in these early humans indicates there is little if any consumption of marine foods, it is likely that the enrichment of δ15N values is a result of a diet high in protein from higher-ranked terrestrial animals.

Figure 10. Carbon and nitrogen isotope values from MHCP and ST. Symbols represent foragers, transitional agriculturalists, and farmers. Data are plotted over published values for Maya Classic period maize farmers from the Maya lowlands region (Somerville et al. Reference Somerville, Fauvelle and Froehle2013). (a) Bivariate plot of δ13C vs δ15N from bone collagen; (b) δ13C of apatite plotted against collagen with dietary regression lines derived from experimental feeding studies (Kellner and Schoeninger Reference Kellner and Schoeninger2007); (c) Function 1 versus Function 2 discriminant analysis plotted against isotopic dietary niche data derived from experimental feeding studies (Froehle et al. Reference Froehle, Kellner and Schoeninger2012). Figure modified after Kennett et al. (Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020:Figure 3).
The first evidence that people are shifting their diets towards maize consumption (approaching 30 percent of total diet) appears in individuals between 2750 and 2050 cal b.c., though these diets are highly varied during this 700-year period, and three of the 14 individuals dating to this period have some overlap with the earlier pre-maize population (Figure 10a). Consequently, the δ13C values for collagen values are variable with a mean of −18.3‰, reflecting carbon primarily from consumption of animal protein. We further measured δ13C for bone apatite and these values reflect the consumption of C3 and C4 resources for the total diet, including carbohydrates, lipids, and protein (Ambrose and Norr Reference Ambrose, Norr, Lambert and Grupe1993). These values range from −14.9‰ to −6.6‰, with a mean of −11.2‰. This result indicates maize consumption averaging 25.8 percent of the total diet (Figures 10b and 10c), based on a simple linear mixing model (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). However, since the signal is much less pronounced in δ13C collagen, it is not clear that this is related to significant maize seed consumption. δ15N values during this time period are similar to the pre-maize period, suggesting a similar reliance on local fauna and freshwater gastropods and not on marine resources. It does not appear that animals consuming maize are a significant part of most of these diets, though the elevated δ15N values for bone collagen could indicate some contribution.
After approximately 2050 cal b.c., all our isotopic data show that maize was a staple grain, with consumption comparable in dietary significance to Classic period farmers (1000 cal b.c. to 1000 cal a.d.), with maize contributing around 70 percent of their total diet (Figure 10b). The δ13C values on collagen range between −13.5‰ and −8.2‰, which are consistent with diets high in maize documented across the Maya lowlands (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020; Somerville et al. Reference Somerville, Fauvelle and Froehle2013). The δ15N values are also comparable. Both collagen and apatite δ13C values suggest that a significant proportion of the diet includes the consumption of animals that are consuming C4 plants, which may indicate that management of domesticated animals (i.e., dogs, turkeys) is an important part of the human ecosystem. Again, we do not see evidence for any significant consumption of reef fish or other marine resources.
DISCUSSION
Genomic data from our earliest individuals support the proposition that the first humans in southeastern Mesoamerica were part of a distinctive Native American lineage that is distantly related, though still ancestral, to present-day populations in southeastern Mesoamerica (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018). These individuals show affinities to early founding lineages in the Americas, and one (burial MHCP.14.1.6) is a direct matrilineal descendent of the Anzick lineage, which includes the Anzick 1 skeleton found in Montana. Combined with data from ancient individuals in South America, our data indicate that there were several rapid radiations of humans from North to South America prior to 5300 cal b.c., and that an expansion of the population associated with the Clovis culture was the primary source of the ancestry of many later Central and South Americans (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018).
The genetic data are also consistent with an early splitting lineage that arose from populations that were radiating southward through the neotropics in the Terminal Pleistocene and Early Holocene (Posth et al. Reference Posth, Nakatsuka, Lazaridis, Skoglund, Mallick, Lamnidis, Rohland, Nagele, Adamski, Bertolini, Broomandkhoshbacht, Cooper, Culleton, Ferraz, Ferry, Furtwangler, Haak, Harkins, Harper, Hunemeier, Lawson, Llamas, Michel, Nelson, Oppenheimer, Patterson, Schiffels, Sedig, Stewardson, Talamo, Wang, Hublin, Hubbe, Harvati, Nuevo Delaunay, Beier, Francken, Kaulicke, Reyes-Centeno, Rademaker, Trask, Robinson, Gutierrez, Prufer, Salazar-Garcia, Chim, Gomes, Alves, Liryo, Inglez, Oliveira, Bernardo, Barioni, Wesolowski, Scheifler, Rivera, Plens, Messineo, Figuti, Corach, Scabuzzo, Eggers, DeBlasis, Reindel, Mendez, Politis, Tomasto-Cagigao, Kennett, Strauss, Fehren-Schmitz, Krause and Reich2018). Early skeletons from MHCP and ST date to 7300–5350 cal b.c. and are genetically related equally to Terminal Pleistocene and Early Holocene individuals from North and South America and to modern descendent communities in lower Central and South America.
During the Terminal Pleistocene and Early Holocene, we see a suite of stemmed and barbed bifacial points being produced from southern Mexico through the Isthmo-Colombian region. These points started being produced at the end of the Younger Dryas, following Clovis-like and Fishtail traditions, and continue until approximately 7000–6000 cal b.c., when they largely disappear. While we have recovered three Lowe points and two large bifaces with traits shared with Lowe (large bifaces with alternate edge beveling), several other large and stemmed bifacial tool types are known from Belize and are thought to be of Terminal Pleistocene or Early Holocene age (Sawmill, Ya'axché, Allspice), but remain undated (Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). These artifacts all bear resemblance to Terminal Pleistocene and Early Holocene points (Figures 2a–2l) found in the isthmian region (Ranere and Cooke Reference Ranere and Cooke2021), suggesting they are roughly contemporaneous and were adaptations to local tropical environments at a time when humans were still engaged in more large animal hunting and processing than in subsequent periods.
After 6000 b.c. there is little evidence for bifacial tool production in neotropical Mesoamerica or the isthmian region until the Formative period (Paris Reference Paris2012; Prufer et al. Reference Prufer, Alsgaard, Robinson, Meredith, Culleton, Dennehy, Magee, Huckell, James Stemp, Awe, Capriles and Kennett2019). In Panama and at MHCP and ST, bifacial thinning flakes are common in contexts before 6000 b.c. and largely absent in younger contexts prior to the Formative period. We are aware of only one published bifacial point from northern Colombia found within a well-dated Middle Holocene context (Castillo Espitia and Aceituno Reference Castillo Espitia and Aceituno2006). This decline in bifacial tool production is likely related to the shift in food procurement strategies, away from hunting larger animals towards an increasing focus on smaller animals and plants during the thermal maximum, the wettest and warmest period of the Holocene (Renssen et al. Reference Renssen, Seppä, Heiri, Roche, Goosse and Fichefet2009). This change is also seen in a shift towards smaller mammals in the faunal assemblages at MHCP (Orsini Reference Orsini2016).
During the Terminal Pleistocene and Early Holocene (8000–6000 cal b.c.), foraging populations across the neotropics and adjacent regions were increasingly consuming some of the earliest cultigens or their wild progenitors. In southwest Mexico the earliest maize domesticates were exploited only briefly prior to diffusing into South America, where they were widely distributed across Greater Amazonia and the Pacific coast. There, maize underwent selection prior to improved varieties moving northward (Kistler et al. Reference Kistler, Thakar, VanDerwarker, George, Harper, Allaby, Hirth and Kennett2020). However, there we lack strong evidence for when maize grain became a significant component of the diet. Without isotopic data from directly dated skeletons from Greater Amazonia or the isthmian region, we can make only a couple of limited statements: (a) The earliest data come from the north coast of Peru, where maize initially became a significant part of the human diet as a weaning gruel after 4000 b.c. and was a staple in the diets of some individuals after 3000 b.c. (Tung et al. Reference Tung, Dillehay, Feranec and DeSantis2020); and (b) in southern Belize, humans dating prior to 2750 cal b.c. are not consuming maize in any quantities detectable by bulk stable isotope measurements of bone collagen or apatite. The shift from limited to staple consumption of maize occurred over a 700-year period from 2750 to 2050 cal b.c. (Kennett et al. Reference Kennett, Prufer, Culleton, George, Robinson, Trask, Buckley, Moes, Kate, Harper, O'Donnell, Ray, Hill, Alsgaard, Merriman, Meredith, Edgar, Awe and Gutierrez2020). Currently available data suggest that this shift to staple consumption of maize happened 800 years prior to the first documented settlers (Iceland and Hirth Reference Iceland, Hirth, Jon C., Borejsza and Joyce2021) and communities in the Maya lowlands (Castellanos and Foias Reference Castellanos and Foias2017; Inomata et al. Reference Inomata, Triadan, López, Fernandez-Diaz, Omori, Bauer, Hernández, Beach, Cagnato and Aoyama2020). This may indicate that while early farmers of maize (around 2050–3200 b.c.) in the region were consuming the crop in quantities comparable to the Classic period, they were engaged in much more flexible social relationships and living in mobile or semisedentary communities in some places, and perhaps living alongside groups building incipient public architecture by around 1000 cal b.c. (Ebert et al. Reference Ebert, Hoggarth, Awe, Culleton and Kennett2019; Inomata et al. Reference Inomata, MacLellan, Triadan, Munson, Burham, Aoyama, Nasu, Pinzón and Yonenobu2015). This might indicate that maize, already a domesticate for 5000 years and available in its fully domesticated form for over 1000 years, was not an immediate driver of changes in mobility as it became ensconced in the human diet. Alternately, it may be that early settlements in southern Mesoamerica were ephemeral, built from perishable materials, and produced simple ceramic vessels, which were widely in use by 2000 b.c. across lower Central America and even earlier in parts of Greater Amazonia (Iizuka Reference Iizuka2017; Joyce and Henderson Reference Joyce and Henderson2001; Roosevelt et al. Reference Roosevelt, Housley, Da Silveira, Maranca and Johnson1991).
Interpreting the complex history of the humans in the neotropics has been hampered by a general lack of data, particularly in southeastern Mesoamerica, where few well-preserved sites have been identified, particularly for the Early and Middle Holocene. Our data from MHCP and ST fill a substantial gap, while raising other questions. Belize was a mixing zone for two Terminal Pleistocene foraging traditions, with evidence that it was on the margin of both the North American Clovis tradition and the South American Fishtail tradition. However, until Clovis-like or Fishtail tools are recovered from well-dated contexts, we will not know more about the timing of this overlap or how these traditions diffused into middle America. Subsequently in the Early Holocene the tradition of stemmed and barbed bifacial tools was widely shared across Central America and the Isthmo-Colombian region, and from Belize we have well-dated estimates of when these were produced and when this tradition ended. Ancient DNA suggests that people living in Belize at that time also shared ancestry with people living today in lower Central America, South America, and North America, though in the absence of additional data from ancient skeletons south of Belize it is not possible to assess the degree to which they overlap. In the Middle Holocene we see increasing developments in plant management and likely early horticulture. Archaeological data suggest that maize was widely dispersed across the neotropics before 5000 b.c., but that farming of more productive varieties of maize likely appeared earlier in South America (Kistler et al. Reference Kistler, Yoshi Maezumi, de Souza, Przelomska, Costa, Smith, Loiselle, Ramos-Madrigal, Wales, Ribeiro, Morrison, Grimaldo, Prous, Arriaza, Thomas, Gilbert, de Oliveira Freitas and Allaby2018), with the evidence of consumption of maize grain as an integral part of the diet coming from coastal Peru as early as 4000 cal b.c. (Tung et al. Reference Tung, Dillehay, Feranec and DeSantis2020), some 1,300 years earlier than in Belize.
The adoption of maize in the neotropics as a dietary staple long preceded the development of social relationships and institutions associated with surplus production. It is almost certain that maize agriculture was a catalyst that, once it was established, propelled the rise of permanent villages and, eventually, the political and economic institutions that characterize the next two millennia, but this process was not immediate and likely dovetailed with the gradual abandonment of reliable mobile foraging-farming modes of subsistence. For the earliest investors in intensifying maize production, its elevation to the status of the primary staple of the diet was likely contingent on a long period of risk assessment and a re-ranking of economically important crops. Thus, the complex changes of the Terminal Pleistocene through the Middle Holocene set the stage for the complex economic and political institutions that followed.
ETHICS STATEMENT
All ancient skeletons from MHCP and ST rock shelters were excavated by the Bladen Paleoindian and Archaic Archaeological Project (BPAAP) under permits issued by the Belize Institute of Archaeology (IA) and the Belize Forest Department. Skeletons of ancient individuals were exported under permits issued by the IA to Prufer, in accordance with the laws of Belize and permission granted to conduct molecular analyses on bulk tissues extracted from skeletons of ancient individuals. This research was conducted in close collaboration with the Ya'axché Conservation Trust, an internationally recognized Belizean NGO that is the co-manager of the Bladen Nature Reserve with the Government of Belize. Ya'axché is locally managed and largely staffed by members of descendent Maya communities. As part of this collaboration, BPAAP research proposals are annually reviewed by the Ya'axché administrative and scientific staff. In 2016 and 2018 K.M.P. gave consultation presentations to the local staff of Ya'axché and other interested community members on our research, including discussions of isotopic and genomic studies on ancient humans from Belize. In 2020, in coordination with Ya'axché, K.M.P. and D.J.K. invited indigenous leaders and community members from villages proximate to the Bladen Nature Reserve to consult on this research. We provided advance invitations and arranged transportation for 46 people from five villages to attend the public consultation. K.M.P. delivered a one-hour presentation detailing the fieldwork, laboratory work, and the results of this study, and an additional hour was devoted to answering questions and clarifying the data and interpretations. Community members requested future public consultations to update them on additional research results, as well as copies of all study results in English, with translations into Mopan and Q'eqchi' languages. Additionally, in 2017 and 2018–2019, results of this research were presented by K.M.P at the annual Belize Archaeology Symposium (BAS), a widely publicized venue sponsored by the IA that is promoted as archaeology for the public. The BAS affords the opportunity for both the presentation of research results and feedback from professional and public communities (including members of indigenous, Maya-speaking communities).
RESUMEN
Los datos de los abrigos rocosos del sur de Belice muestran evidencias de fabricación de herramientas, caza y explotación de recursos acuáticos hacia 10.500 cal a.C., y funcionaron como sitios mortuorios entre 7600 y 2000 cal a.C. Los contextos del holoceno temprano contienen bifaces con vástago y púas como parte de una tradición que se encuentra ampliamente en todo el neotrópico. Después de 6000 cal a.C., las herramientas bifaciales desaparecen del registro, reflejando los cambios en la subsistencia cuando la gente se volvió más dependiente de los alimentos vegetales y aparecieron los primeros domesticados. Sugerimos que los habitantes del sur de Belice mantenían estrechos vínculos con sus vecinos del sur. Los esqueletos de 7650–5350 cal a.C. están relacionados genéticamente con individuos antiguos de 11.000–7000 cal a.C. de América del Norte y del Sur, y con poblaciones modernas de la parte baja de América Central y del Sur, pero en menor medida con los mexicanos modernos. Esto sugiere una historia compleja para el neotrópico, un área de información y tecnologías compartidas. El cultivo del maíz en Belice estuvo probablemente influenciado por las variedades de maíz del noroeste de Sudamérica que se extendieron hacia el norte antes de 3000 cal a.C. Situamos la adopción del maíz como alimento básico de la dieta entre 2750–2050 cal a.C., basándonos en los isótopos estables del carbono de un notable transecto de esqueletos humanos, fechados por radiocarbono directamente en estos abrigos rocosos—esto es, 1000–1500 años más tarde que en Sudamérica.
ACKNOWLEDGMENTS
Research presented here was funded by the National Science Foundation (BCS: 1632061 Prufer, 1632144 Kennett) and the Alphawood Foundation (Prufer). We acknowledge the dedicated efforts of all our collaborators at Harvard University (Reich Lab), Pennsylvania State University (B. Culleton, Institute for Energy and the Environment), and the University of New Mexico (N.V. Atudorei, S. D. Newsome, Center for Stable Isotopes), and all of our field- and lab-based students and team members since 2014. Special thanks to Julie Saul and the late Frank Saul for initiating this study more than two decades ago, as part of the Maya Mountains Archaeological Project directed by P.S. Dunham. We also thank the Belize Institute of Archaeology and the Forest Department for permits to conduct fieldwork. Our research was conducted in collaboration with the Ya'axché Conservation Trust (https://yaaxche.org/), whose mission to protect the biodiversity and cultural history of the Bladen Nature Reserve is a core goal of this project. We thank all our collaborators from the Mopan and Q'eqchi' communities of Southern Belize, especially Jose Mes, Raymundo Sho, Oligario Sho, Francisca Bardalez, Mateo Rash, and Sylvestre Rash.