


Introduction
It is well known that the duration of sperm–oocyte co-incubation, the sperm concentration and the presence of cumulus cells are simple but major factors for successful in vitro fertilization (Iwata et al., Reference Iwata, Shiono, Kon, Matsubara, Kimura, Kuwayama and Monji2008). It has been suggested that prolonged gamete co-incubation, in which high concentrations of spermatozoa are incubated in small volumes of medium, results in the production of high levels of hydrolytic (Rehman et al., Reference Rehman, Collins, Suh and Wright1994) and free radicals (Aitken, Reference Aitken1994) that damage the oocytes. Furthermore, increasing the sperm–oocyte incubation time has been found to correlate with a higher incidence of polyspermy (Sumantri et al., Reference Sumantri, Boediono, Ooe, Murakami, Saha and Suzuki1997; Gasparrini et al., Reference Gasparrini, De Rosa, Attanasio, Boccia, Di Palo, Campanile and Zicarelli2008), suggesting that it would be advisable to remove the presumptive zygotes from in vitro fertilization (IVF) medium as early as possible, once the spermatozoa–oocyte interaction has been accomplished to minimize the time the presumptive embryos are exposed to adverse conditions, improving even their implantation rate (Gianaroli et al., Reference Gianaroli, Magli, Ferraretti, Fiorentino, Tosti, Panzella and Dale1996). Moreover, an excessive gamete co-incubation period could increase the polyspermy incidence due to the ageing of the oocytes, reducing their capacity to block the polyspermy (Cheng, Reference Cheng1985; Pavlok et al., Reference Pavlok, Torner, Motlik, Fulka, Kauffold and Duschinski1988, Crozet, Reference Crozet1991a).
In the same way, a strong correlation between the incidence of polyspermy and the absolute number of capacitated spermatozoa present at the fertilization site suggests that the reduction in the number of spermatozoa could minimize in vitro polyspermy (Gil et al., Reference Gil, Almiñana, Roca, Vázquez and Martinez2008). However, in most cases, such changes are also accompanied by a low oocyte penetration rate. In fact, under in vitro conditions high sperm concentration is necessary to maintain the motility of the spermatozoa and also to obtain a sufficient number of capacitated sperm due of the low efficiency of the in vitro capacitation systems (reviewed by Crozet et al., Reference Crozet, Thibault and Levasseur1991b). In these situations, the ideal IVF system would be one that resulted in a high penetration rate with a low incidence of polyspermy. Specifically in goats, the sperm concentration in IVF medium normally used in the different laboratories is approximately 1 × 106 spermatozoa/ml (Jufen et al., Reference Jufen, Zhiming, Yong and Jianchen1991; Younis et al., Reference Younis, Zuelke, Harper, Oliveira and Brackett1991; Martino et al., Reference Martino, Palomo, Mogas and Paramio1994, De Smedt et al., Reference De Smedt, Crozet, Ahmed-Ali, Martino and Cognié1992; Bormann et al., Reference Bormann, Ongeri and Krisher2003; Koeman et al., Reference Koeman, Keefer, Baldasarre and Downey2003; Katska-Knazkiewicz et al., Reference Katska-Ksiazkiewicz, Rynska, Gajda and Smorag2004; Rodríguez-Dorta et al., Reference Rodríguez-Dorta, Cognié, González, Poulin, Guignot, Touzé, Baril, Cabrera, Álamo, Batista, Gracia and Mermillod2007), although concentrations of 2 × 106 (Pawshe et al., Reference Pawshe, Totey and Jain1994); 12–15 × 106 (Herrick et al., Reference Herrick, Lane, Gardner, Behboodi, Memeli, Blash, Echelard and Krisher2006) or even sperm concentrations up to 20 × 106 sperm/ml (Chauhan & Anand, Reference Chauhan and Anand1991) have been described.
On the other hand, it is well documented that the gamete interaction process allows for the penetration of oocyte by sperm and the efficacy of this process depends on the vitality and capacity of the sperm as well as on the maturational status of the oocyte at the time they encounter sperm. Regardless of the system used, spermatozoa must undergo capacitation and acrosome reaction (Sidhu & Guraya, Reference Sidhu and Guraya1989) and they need sufficient interaction time to accomplish penetration of the oocytes and to initiate the subsequent events. According to Dode et al. (Reference Dode, Rodovalho, Ueno and Fernandes2002), the earlier the spermatozoa are capacitated, the earlier the penetration will occur avoiding the aging process of the oocyte, which can also cause abnormal fertilization. However, the variations on the fertilization and embryonic development rates due to the different co-incubation times could also be a manifestation of the heterogeneity of the required time for the spermatozoa to complete the capacitation (Parrish et al., Reference Parrish, Susko-Parrish, Winer and First1988). Regardless of the method used, there is heterogeneity of the sperm population and the spermatozoa do not complete capacitation simultaneously. Actually, even under the same conditions some spermatozoa from the same ejaculate capacitated faster than others (Yanagimachi, Reference Yanagimachi, Knobil and Neill1994). Likewise, differences among males in terms of sperm capacitation times and acrosome reaction have also been reported (Parrish et al., Reference Parrish, Susko-Parrish, Leibfried-Rutledge, Critser, Eyestone and First1986; Yanagimachi, Reference Yanagimachi, Knobil and Neill1994). Therefore, the optimum time of interaction between the gametes could vary in function of different factors, from the efficiency of the in vitro capacitation method including the differences observed among spermatozoa, ejaculates and males on the required time to be capacitated to the efficiency of the maturation systems, when the necessity of additional time for the oocytes to achieve the cytoplasmatic maturation could also exist.
Generally, induced capacitation and acrosome reaction processes are evaluated in function of the obtained blastocyst rate, which is not the most suitable parameter to evaluate semen samples as this rate could be much more dependent on culture conditions during embryo development than the sperm penetration rate. The formation of two pronuclei is a criterion of IVF success. Such a criterion seems to be unambiguous and it clearly defines an in vitro fertilizing potential (Jedrzejczak et al., Reference Jedrzejczak, Pawelczyk, Taszarek-Hauke, Kotwicka, Warchol and Kurpisz2005). In fact, when IVF systems are employed, the penetration rate is the commonly used measure of the sperm fertilizing ability (Gadea, Reference Gadea2005). Furthermore, we suggest that to know the efficiency of the capacitation system used could offer considerable information about the IVF process, helping to determine the optimum conditions of the gamete co-incubation and improving enormously the embryo production in goats. As a successful method of capacitation can be assessed either by oocyte penetration in vitro or by making use of differential staining techniques, another objective in the present work was to obtain more information from our in vitro capacitation induction through the study of both aspects simultaneously. The evolution of the penetration rate and the different sperm parameters, with special attention to the acrosome reaction, along the different gamete co-incubation periods could explain their relationship.
On the other hand, even the acrosome reaction can take place independently from the oocyte, it has been demostrated that the cumulus oophorus as well as the zona pellucida can induce the acrosome reaction in the spermatozoa (reviewed by Yanagimachi Reference Yanagimachi, Knobil and Neill1988, Reference Yanagimachi, Knobil and Neill1994). In fact, Tanghe et al. (Reference Tangue, Van Soom, Mehrzad, Maes, Duchateau and de Kruif2003) showed evidence that the cumulus benefits the fertilizing ability of penetrating spermatozoa by creating a complex microenvironment of both cumulus secretions and metabolic products around the oocyte. In cattle, the beneficial effect of cumulus on the fertilization process can be explained partially by facilitating sperm capacitation, acrosome reaction and penetration (Fukui, Reference Fukui1990; Younis & Brackett, Reference Younis and Brackett1991; Cox et al., Reference Cox, Hormazabal and Santa María1993). In the present work, we have not evaluate this beneficial effect of the cumulus on the penetration rate comparing cumulus–oocytes complexes (COCs) versus denudated oocytes, simply we attempted to assess the effect of the COCs on the different sperm parameters in the gamete co-culture.
Based on these observations, the aims of the present study were to evaluate the influence of sperm concentration and length of the gamete co-incubation on fertilization and to investigate the kinetics of different semen parameters during the IVF, assessing the effect of the presence of COCs in these parameters, in order to determine the optimum conditions of the gamete co-incubation in prepubertal goats.
Materials and Methods
Recovery and IVM of oocytes from prepubertal goats
Ovaries were recovered from prepubertal goats (younger than 2 months old) at a commercial slaughterhouse. Once in the laboratory, the ovaries were washed three times in phosphate-buffered saline (PBS) plus gentamycin (50 mg/ml) at 38.5°C. Oocytes were obtained by slicing the ovaries in TCM199/HEPES medium (Sigma M-2520) supplemented with 2.2 mg/ml sodium bicarbonate and 50 μg/ml gentamicin. Oocytes with one or more complete and compact layers of cumulus cells and a evenly granulated cytoplasm were selected and cultured in TCM199/HEPES medium supplemented with 2.2 mg/ml of sodium bicarbonate, 50 μg/ml of gentamicin, 20% of inactivated estrus goat serum (EGS), 10 μg/ml ovine follicle stimulating hormone (FSH) (Ovagen, InmunoChemical Products Ltd), 10 μg/ml caprine luteinizing hormone (LH) (supplied by J.F. Beckers, IRSIA Research Unit, University of Liège, Belgium) and 1 μg/ml of 17-β-estradiol (Sigma, E-2257). Oocytes were matured in 50 μl microdrops of medium (eight oocytes per microdrop) covered with paraffin oil at 38.5°C for 27 h in an atmosphere of 5% CO2 in saturated humid air.
Spermatozoa processing
Ejaculates were collected via an artificial vagina from three Murciano-Granadina males of proven fertility and transported to the laboratory at 37°C. The percentage of progressive motility was assessed. All samples showing less than 60% motile sperm cells were rejected. Ejaculates were mixed in equal quantities in order to minimize any variation between males. Spermatozoa were selected by swim-up depositing 50 μl of ejaculate mixture in each of several tubes containing 2 ml of HEPES-TALP (Parrish et al., Reference Parrish, Susko-Parrish, Leibfried-Rutledge, Critser, Eyestone and First1986). After 1 h at 38.5°C, the top 0.6 ml from each tube was removed and pooled in a sterile 15-mL centrifuge tube and centrifuged (200 g) for 10 min. Immediately after, the supernatant was rejected and the sediment was diluted (1:1) in HEPES-TALP medium with 100 μg/ml heparin sodium salt (Sigma H-3393) with a final incubation concentration of 50 μg/ml of heparin and incubated at 38.5°C for 45 min.
IVF procedure
Three experiments were performed. The followed procedure before the insemination of the oocytes was the previous described methodology.
Experiment 1
In this experiment the effect of the sperm concentration in the IVF microdrop was studied. After the maturation culture interval, COCs were transferred to four groups of microdrops (14 to 16 oocytes per 95 μl) of fertilization medium (TALP; Parrish et al., Reference Parrish, Susko-Parrish, Leibfried-Rutledge, Critser, Eyestone and First1986), supplemented with 1 μg/ml of hypotaurine (Sigma, H-1384). After heparin treatment, sperm concentration was assessed in a hemacytometer. An aliquot of sperm suspension was added to each fertilization drop for a final concentration in each group of: (a) 0.5 × 106; (b) 1 × 106; (c) 2 × 106; and (d) 4 × 106 spermatozoa/ml, resulting in an approximate sperm:oocyte ratio of (a) 3500; (b) 7000; (c) 14,000; and (d) 28,000, respectively. Insemination was under paraffin oil in a humidified 5% CO2 atmosphere at 38.5°C. At 17 h of gamete co-incubation, the presumptive zygotes were fixed to evaluate penetration rate and a sperm sample from each treatment was saved in order to evaluate motility, viability and acrosome status.
Experiment 2
In this experiment the IVF procedure was identical to the previous experiment, but all of COCs were inseminated with a final sperm concentration of 4 × 106sperm/ml approximately in the microdrops (14 to 16 oocytes per 95 μl) of fertilization medium. Immediately after, representative groups of 25–30 oocytes were fixed approximately at 2, 4, 6, 8, 10, 12, 16, 20, 24 and 28 h after insemination to evaluate nuclear status. At same time, samples of sperm which had been in contact with the oocytes were collected in order to evaluate motility, viability and acrosome status.
Experiment 3
In this experiment, groups of 14–16 oocytes/microdrop were inseminated following the same protocol that in the previous experiment. Moreover and in a parallel way, sperm samples were incubated under identical conditions, but without the presence of COCs. Immediately after, sperm samples were collected from the microdrops with or without COCs at 2, 4, 6, 8, 10, 12, 16, 20, 24 and 28 h after insemination in order to evaluate motility, viability and acrosome status.
Assessment of the results
To evaluate the nuclear stage at the different post insemination intervals, oocytes were separated from the spermatozoa attached to the zona pellucida by agitation in an isotonic solution of sodium citrate at 3% and fixed for 3 to 4 days in acetic acid 90% ethanol (1:3, v:v) at 4°C. Afterwards they were stained with 1% lacmoid and the nuclear stage was observed under a phase contrast microscope. The oocytes with a spermatozoa tail in the cytoplasm were considered penetrated.
The sperm parameters studied were progressive forward motility, viability and acrosome integrity of sperm samples collected from IVF microdrops. Sperm viability and acrosome integrity were evaluated using a simplified triple staining technique described in goats by Kusunoki et al. (Reference Kusunoki, Yasui, Kato and Kanda1984). This technique, described briefly in elsewhere (Palomo et al., Reference Palomo, Izquierdo, Mogas and Paramio1999), allowed for the differentiation of viable acrosome reacted spermatozoa from the total population. The percentage of progressive forward motile sperm was assessed subjectively in a 10 μl drop of sperm suspension placed on a warm slide covered with cover slip, using a phase contrast microscope (×200) fitted with a stage warmer at 38°C.
Statistical analysis
All the data expressed in percentages or fractions were subjected to arcsin transformation and then examined by analysis of variance of the treatment factor, with the date factor as the block factor, using the General Linear Model (GLM) of the statistics program SAS. Statistical significance was established as maximum probability value of 0.05% for all parameters. When a statistically significant effect was found in the treatment factor multiple comparisons using the least square deconvolution (LSD) method were made. The figures which refer to the different semen parameters were expressed as LS means (least square means) ± standard error (SE) of the LS means, while the figures which refer to the results of penetration rate were expressed as frequencies. Likewise, potential correlations among different parameters were analysed using the same statistics program described before.
Results
Experiment 1
No significant differences were found between the different sperm concentrations in the IVF microdrop on any of the sperm parameters evaluated at 17 hpi (Table 1). The total penetration rate and the polyspermy incidence were significantly higher as we used sperm concentration superior to 2 × 106 spermatozoa/ml (Table 2). However, the different sperm concentrations in the IVF microdrop tested in this experiment did not affect significantly the normal fertilization rate, although the maximum percentage was obtained with a sperm concentration of 4 × 106 spermatozoa/ml (oocyte: ratio of 28,000:1), not showing significant differences on the polyspermy incidence when we compared these results with the immediately lower sperm concentration.
Table 1 Sperm parameters* evaluated at 17 hpi according to the sperm concentration on IVF

*Least square (LS) means ± SE.
Table 2 Effect of different sperm concentration in the IVF microdrop on the penetration rated

a–cDifferent superscripts within columns indicate significant differences (p < 0.05).
dValues expressed as frequencies.
2PN, 2 pronuclei + sperm tail; PS, polyspermic; Abnorm, abnormally fertilized.
Experiment 2
The results concerning to sperm parameters studied in this experiment are showed in the Fig. 1. After the different gamete co-incubation periods, a negative effect on the sperm motility (p < 0.05) and viability (p < 0.0001) was observed as the length of the insemination period increase. Likewise negative coefficients were obtained when we correlated the percentages of motility (r = −0.55, p < 0.001), viability (r = −0.77, p < 0.0001) and even the percentage of acrosome integrity (r = −0.49, p < 0.005) with the length of the co-culture, confirming the detrimental effect of co-culture time on the spermatozoa.

Figure 1 Evolution of different sperm parameters during the gamete co-culture (least square (LS) means ± SE) in the presence of cumulus–oocytes complexes (COCs).
Analyzing the evolution of the different sperm categories obtained by the triple stain after different gamete co-culture time (Fig. 2), only a significant effect (p < 0.0001) on the percentages of intact acrosome sperm (viable and non-viable sperm) was observed, whereas the other categories were not significantly affected. Nevertheless, the percentage of non-viable acrosome-reacted sperm increased from values about 15% during the first 8 h of co-culture to values near to or higher to 20% in the next hours.

Figure 2 Evolution of the different sperm categories obtained by triple stain after different gamete co-culture time (p ≤ 0.0001, least square (LS) means ± SE) and the evolution of the penetration rate.
Related to the percentage of viable acrosome-reacted sperm, even not significantly (p = 0.0573), we could observe an effect of co-culture time on this parameter. In fact, we observed the maximum incidence of acrosome reaction between 8 and 12 h of co-culture, showing the maximum percentage (5.7%) at 10 h of the gamete co-culture, while the penetration rate was maximal at 12 h of co-incubation (Fig. 2), although not significant correlations were found between both parameters. In the same way, no significant correlationship was found among any of the sperm parameters analysed in any of the studied co-culture periods and the penetration rate.
The evolution of the penetration rate is showed in the Table 3. The first evidence of oocyte penetration (12.7%) was observed after 4 h of insemination. This rate increased until to reach values near or higher to 30% (37.2% at 12 h), although from 6 h of gamete co-culture, not significant differences were observed. In a similar manner, from the first 4 h of insemination, the percentage of normal fertilization obtained was similar after different insemination times reaching values higher to 20%, except in only at 16 hpi. The polyspermy incidence and the percentage of abnormally fertilized oocytes never exceeded 10% and 11.5%, respectively in any of the studied periods.
Table 3 Effect of different gamete co-culture length in IVF on the penetration rated

a–cDifferent superscripts within columns indicate significant differences (p < 0.05).
dValues expressed as frequencies.
2PN, 2 pronuclei + sperm tail; PS, polyspermic; Abnorm, abnormally fertilized.
Experiment 3
By analyzing the results obtained in this experiment, no significant interaction was observed between the co-culture time and the presence or absence of COCs on motility, viability and acrosome integrity percentages. Nevertheless, a positive effect of the presence of COCs was observed on viability and acrosomal integrity of the incubated sperm (p < 0.05 and p < 0.01, respectively), whereas the percentage of mobile sperm was significantly higher (p < 0.05) when the COCs were absent (Fig. 3).
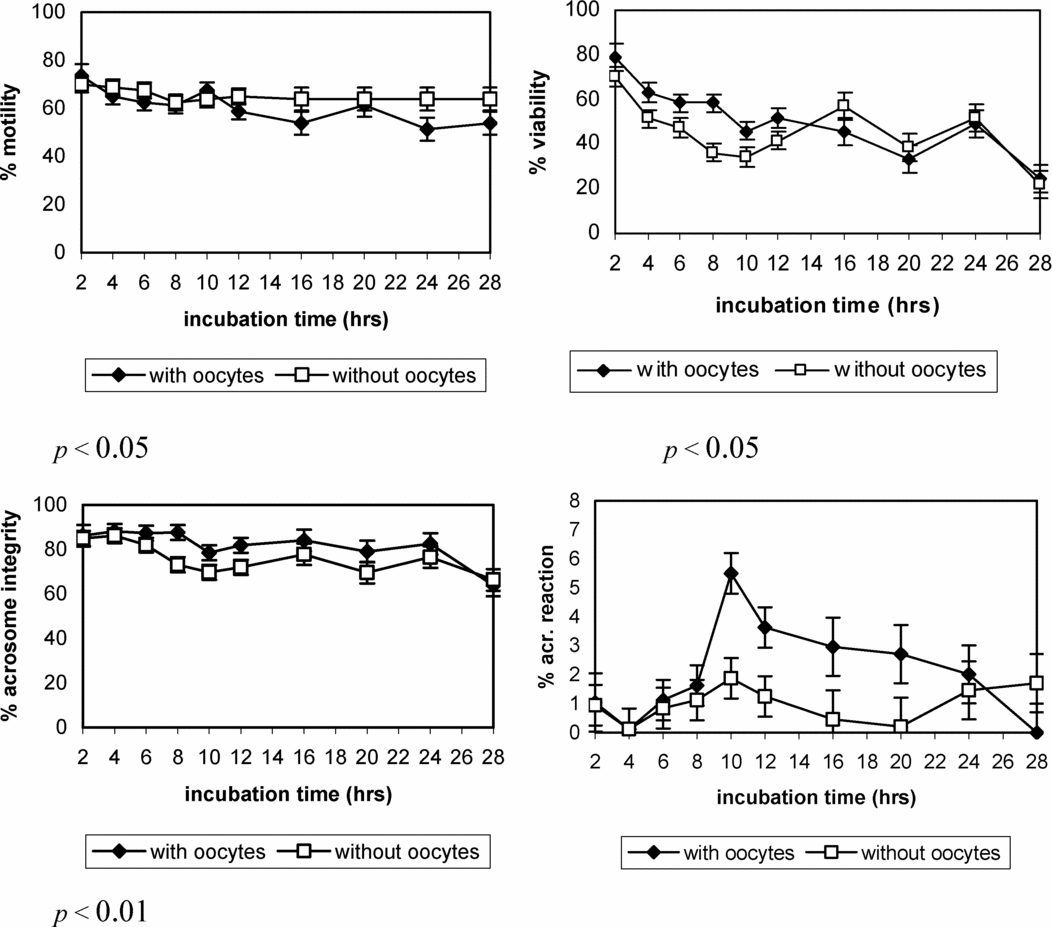
Figure 3 Evolution of different sperm parameters (least square (LS) means ± SE) during the gamete co-culture in the presence or absence of cumulus–oocytes complexes (COCs).
No significant interaction was found between the incubation time and the presence of COCs on the percentage of viable acrosome-reacted sperm. Likewise, even the presence or absence of COCs had no significant effect on this sperm parameter, the incidence of acrosome reaction was higher when the COCs were present. In the same way that the previous experiment, not significant effect (p = 0.0502) of the sperm incubation time was found on the percentage of viable acrosome–reacted sperm, showing a similar evolution along the incubation time in both experiments.
Discussion
In the present study we found that the balance between having a sufficient number of sperm in order to obtain acceptable fertilization rates ensuring a minimal incidence of polyspermy is achieved with the concentration of 4 × 106 spermatozoa/ml (sperm:oocyte ratio 28,000:1), as we obtained the best rate of total penetration (43.4%) and normal fertilization (15.7%), showing values of polyspermy incidence (15.6%) even lower than those obtained by De Smedt et al. (Reference De Smedt, Crozet, Ahmed-Ali, Martino and Cognié1992) with adult goat oocytes (20%) using a sperm concentration of 1 × 106. Although no significant differences were found between sperm concentrations of 2 and 4 × 106 spermatozoa/ml used in this experiment.
Related to this finding, an interesting analysis of Rath (Reference Rath1992) in pigs showed a high correlation between polyspermy and the absolute number of spermatozoa and oocytes (i.e. sperm:oocyte ratio), but not between polyspermy rate and sperm concentration per millilitre. This suggests that the sperm:oocyte ratio is more relevant in IVF outcomes and could be one of the explanations of the different results obtained in the different research groups on caprine IVF. In fact, among the different works which have used the concentration of 1 × 106 spermatozoa/ml, the sperm: oocyte ratio could vary from values of 5–10 × 103:1 (Kenkistepe et al., Reference Keskintepe, Simplicio and Brackett1998; Katska-Knazkiewicz et al., Reference Katska-Ksiazkiewicz, Rynska, Gajda and Smorag2004) to 100–200 × 103:1 (De Smedt et al., Reference De Smedt, Crozet, Ahmed-Ali, Martino and Cognié1992; Martino et al., Reference Martino, Palomo, Mogas and Paramio1994) or even values of 800 × 103:1 (Chauhan & Anand, Reference Chauhan and Anand1991). In the present work, in spite of have been using higher sperm concentrations, our sperm:oocyte ratios not exceed of values of approximately 28,000:1.
On the other hand, it has been described that an excessive generation of free radicals by the high concentration of spermatozoa that are incubated during gamete co-incubation has many detrimental effects on embryos cells. However, these free radicals also have some beneficial effects, such as they are needed for acquisition of fertilizing ability by spermatozoa and hydrogen peroxide is required for induction of capacitation in some species (Watson, Reference Watson2000), which could have occurred in our experiment as we compared the penetration rates obtained between the lower versus the higher sperm concentrations. Likewise, as First & Parrish (Reference First and Parrish1987) described, if the capacitation is not sufficient, very few spermatozoa will be able to fertilize oocytes and the polyspermy will not be a problem, suggesting that probably our capacitation method allowed few spermatozoa to reach the fertilizing ability to penetrate the oocytes, even from a high sperm concentration. However, in preliminary studies, where the compared sperm concentrations were more disparate quantitatively, we found that as we increased the concentration from 5 to 10 × 106 sperm/ml, the normal sperm fertilization rate decreased to the half or even less and the polyspermy incidence increased more than twice without affecting the total sperm penetration (data not shown). Therefore, a delicate balance between the risk the polyspermy, the efficiency of the capacitation system and the amount of free radicals generated on the gamete co-culture could determine whether the final outcome in detrimental or beneficial. Moreover, no significant differences were found between the different sperm concentrations in the IVF microdrop on any of the sperm parameters evaluated at 17 hpi, demonstrating that neither a high dilution in the lowest sperm concentration nor a high reactive oxygen species (ROS) generation in the highest sperm concentration have had any effect on the motility, viability or acrosome integrity of the spermatozoa.
It is well known that in the case of in vitro fertilization, the duration of sperm–oocyte co-incubation is a simple but an important factor in the production of embryos. Unfortunately, few studies are found about the kinetics of the fertilization events and the optimum co-incubation period in the goat IVF. In the present study, the minimum time of gamete co-culture necessary for observing fertilized oocytes was 4 h, whereas in human IVF a spermatozoa-oocyte interaction of 1 h was sufficient, even in semen samples showing defects (Gianaroli et al., Reference Gianaroli, Magli, Ferraretti, Fiorentino, Tosti, Panzella and Dale1996). Different minimum times of in vitro matured oocyte penetration have been described as 3 h (Saeki et al., Reference Saeki, Kato, Hosoi, Miyake, Utsumi and Iritani1991), 6 h (Xu & Greve, Reference Xu and Greve1988) and 8 h (Hyttel et al., Reference Hyttel, Xu and Greve1988) in the bovine, 2 h in the ovine (Cheng, Reference Cheng1985) or 1 h in the caprine (Cox et al., Reference Cox, Saravia, Briones and Santa María1995). Likewise, these differences could affect the necessary time for obtaining the maximum penetration rate. In fact, Xu & Greve (Reference Xu and Greve1988) obtained the maximum penetration rate in bovine oocytes with an interval of 28 h, whereas Saeki et al. (Reference Saeki, Kato, Hosoi, Miyake, Utsumi and Iritani1991) obtained this maximum after 5 h of co-culture. In our conditions, the maximum penetration rate was showed at 12 h, although from 6 h of gamete co-culture not significant differences were observed. Similar timing of goat sperm penetration was shown by Cox et al. (Reference Cox, Saravia, Briones and Santa María1995) using bovine matured oocytes.
Nevertheless, the speed of sperm penetration into the zona pellucida in vitro is highly dependent on several factors such the efficiency of capacitation systems, in addition to the own differences of each specie. In bovine, when the duration of sperm–oocyte incubation was less than 4 h, the fertilization rate was much lower (Saeki et al., Reference Saeki, Kato, Hosoi, Miyake, Utsumi and Iritani1991; Pavlok, Reference Pavlok2000). In a similar manner, also in cattle, it has been shown that the oocyte cleavage rate is reduced by decreasing the co-incubation time from 10 to 5 h (Sumantri et al., Reference Sumantri, Boediono, Ooe, Murakami, Saha and Suzuki1997; Ward et al., Reference Ward, Enright, Rizos, Boland and Lonergan2002; Kochhar et al., Reference Kochhar, Kochhar, Basrur and King2003). These results were confirmed by the significant increase in both penetration and cleavage rates observed at 6 h of co-incubation, where fertilization rates continued to increase up to 12 h of incubation no showing differences between 12 and 18 h of incubation (Dode et al., Reference Dode, Rodovalho, Ueno and Fernandes2002), demonstrating that, regardless the method used, there is heterogeneity of the sperm population and spermatozoa from the same ejaculate do not complete capacitation simultaneously (Parrish et al., Reference Parrish, Susko-Parrish, Leibfried-Rutledge, Critser, Eyestone and First1986; Yanagimachi, Reference Yanagimachi, Knobil and Neill1994). However, several researchers suggest that a shortened time course of oocyte exposure to spermatozoa could improve fertilization rates and subsequent embryo viability, suggesting that a gamete co-culture period of 9–10 h could be sufficient to ensure maximal cleavage and blastocyst yields in cattle (Ward et al., Reference Ward, Enright, Rizos, Boland and Lonergan2002; Kochhar et al., Reference Kochhar, Kochhar, Basrur and King2003).
Also, in ovine IVF, Morton et al. (Reference Morton, Catt, Hollinshead, Maxwell and Evans2005) observed that reducing the gamete co-incubation length increased the speed of oocyte cleavage. The latter observation may have been due to reduced exposure of oocytes to dead spermatozoa during short co-incubation, but was not attributable to earlier penetration of spermatozoa, as oocyte penetration was similar after a short (for 2 to 3 h) and long (18–20 h) gamete co-incubation. In human, Gianaroli et al. (Reference Gianaroli, Magli, Ferraretti, Fiorentino, Tosti, Panzella and Dale1996) suggested that the advanced cellular stages resulting from embryos generated by a shortened exposure to spermatozoa may be associated with not only the removal of spermatozoa but also its potentially damaging by-products, indicating that embryos exhibiting a faster cleavage rate are more viable and therefore capable of implantation. Moreover, a longer co-incubation period increased the incidence of polyspermic fertilization (Chian et al., Reference Chian, Nakahara, Niwa and Funahashi1992; Sumantri et al., Reference Sumantri, Boediono, Ooe, Murakami, Saha and Suzuki1997; Iwata et al., Reference Iwata, Shiono, Kon, Matsubara, Kimura, Kuwayama and Monji2008). Even the co-culture length could affect the proportion of male and female embryos, as Iwata et al. (Reference Iwata, Shiono, Kon, Matsubara, Kimura, Kuwayama and Monji2008) showed in their work and concluded that a shorter duration of co-incubation results in a greater number of male embryos in bovine in vitro fertilization, and this trend partly depends on the difference in the capacitation states of the X- and Y-bearing sperm.
In the goat, the exact time required by the spermatozoa to reach the capacitation, to induce acrosome reaction, to penetrate the cumulus and the zona pelucida and to fuse with the oocyte is poorly described, neither the proportion of spermatozoa which will be able to fertilize is known. In the bovine, Parrish et al. (Reference Parrish, Susko-Parrish, Winer and First1988) showed that to induce the capacitation a minimum of 4 h of heparin exposition was required to detect the plasma membrane changes previous to the acrosome reaction. In fact, Hyttel et al. (Reference Hyttel, Xu and Greve1988) did not observe spermatozoa suffering the acrosome reaction in in vitro bovine matured and fertilized oocyte surface until 6 hpi. However, in the sheep, Crozet (Reference Crozet1988) observed, at 1.25 hpi in in vitro fertilized ovulated oocytes, different spermatozoa with reacted acrosome bound to the zona pelucida surface and some were even penetrating it. Nevertheless, we can assure that, in our conditions, at least a sperm population in contact with the oocytes has been able to suffer all these phenomena within 2 to 4 h of gamete co-culture.
Moreover, even we have not studied the state of the sperm attached to the oocyte surface, it would be worthy to note that the percentage of acrosome reaction in the free spermatozoa which were present in the IVF medium, after removing the oocytes at different post-insemination times, increased along the time until to reach the maximum value at 10 hpi and decreased later until values lower to the obtained at the onset of the gamete co-culture. In the bovine, Fukui (Reference Fukui1990) observed that the proportion of viable acrosome reacted spermatozoa increased significantly at 3 hpi of the COCs, obtaining the maximum acrosome reaction value at 6h, percentage which was decreasing in the next observations. A similar evolution of this parameter was showed during the ram sperm incubation in the absence of oocytes (Garde et al., Reference Garde, García Artiga, Gutiérrez and Vázquez1992). Also in the human, an increase in the percentage of viable acrosome reacted spermatozoa was observed during the first 6 h of incubation, showing similar values during the next 6 h (Talbot & Chacon, Reference Talbot and Chacon1981). The delay observed in the present work, related to these authors, in the observation of maximum percentage of viable acrosome reacted spermatozoa could be due, beside the differences between species, to that heparin induced capacitation is delayed by the effect of the glucose added to the medium. Actually, Parrish et al. (Reference Parrish, Susko-Parrish and First1989) showed that bovine spermatozoa need long incubation times (12 h) to reach the capacitation status in the presence of heparin and glucose. However, Fukui (Reference Fukui1990), who also used heparin in the presence of glucose, did not observe this delay, but suggested that glucose could be the responsible of the low obtained rates of acrosome reaction, which could explain also our poor percentages.
Despite it has been impossible to correlate our results of acrosome reaction incidence with the IVF penetration rates, the fact of observing the maximum penetration rate at 12 hpi and to obtain the highest incidence of acrosome reaction between 8 to 12 hpi could indicate some relationship between both phenomena. Actually, Fukui (Reference Fukui1990) observed that the significant increase in the percentage of viable acrosome reacted spermatozoa between 3 to 6 hpi coincided with the oocyte penetration by the spermatozoon. However, Plachot et al. (Reference Plachot, Mandelbaum and Junca1984) and De Jonge et al. (Reference De Jonge, Rawlins and Zaneveld1988) suggested the no existence of the correlation between the IVF results and the percentage of viable acrosome reacted spermatozoa present in the IVF medium, although these authors measured the acrosome reaction incidence at the end of, and not during, the gamete co-culture. Even so, the estimation of induced AR might be a valuable supplement to prediction of the male fertilizing potential, as Jedrzejczak et al. (Reference Jedrzejczak, Pawelczyk, Taszarek-Hauke, Kotwicka, Warchol and Kurpisz2005) suggested after show a significant positive correlation between the proportion of acrosome-reacted spermatozoa and the number of fertilized oocytes.
On the other hand, in spite of the importance of the sperm motility maintenance at high levels during the gamete co-culture on the success of IVF (Cheng, Reference Cheng1985), in the present study it has been impossible to find any correlation between the penetration rates and the percentage of mobile spermatozoa and neither with any of the other sperm parameters evaluated at the end of the different co-culture times. This result can be due probably to the maintenance within acceptable values of the viability as well of the acrosome integrity along the gamete co-culture without compromising the success of the fertilization, although these sperm parameters have been decreasing as the co-culture time goes. Moreover, when we were studying the evolution of the different sperm categories obtained with TST stain during the gamete co-culture, we observed that as the percentage of viable sperm with intact acrosome decreased, the percentage of dead sperm with intact acrosome increased, both significantly. On the other hand, the sperm proportion with acrosome reacted, viable as well dead, was not affected significantly. This result could be due that, in our conditions, the viable acrosome reacted spermatozoa is so little that when these spermatozoa dye quickly after suffering the acrosome reaction, the percentage of dead spermatozoa with reacted acrosome is not significantly affected. However, the fact that the dead acrosome reacted spermatozoa percentage increased to values next or even higher to 20% from 10 h of gamete co-culture, coinciding with the maximum incidence of acrosome reaction, seems to confirm that the sperm evolution cycle described by Didion et al. (Reference Didion, Dobrinsky, Giles and Graves1989), as spermatozoa not capacited–capacited–acrosome reaction–cellular death, also exists.
In this work, the presence of COCs increases the incidence of acrosome reaction in the spermatozoa present in the medium, besides not showing a significant effect. It has been reported that cumulus and the hyaluronic acid included in the extracellular-matrix of cumulus cells induced capacitation in sperm (Suzuki et al., Reference Suzuki, Asano, Eriksson, Niwa, Nagai and Rodriguez-Martinez2002; Hong et al., Reference Hong, Chiu, Lee, Tse, Ho and Yeung2004) improving the fertilization ability of sperm cells (Iwata et al., Reference Iwata, Shiono, Kon, Matsubara, Kimura, Kuwayama and Monji2008) in bovine oocytes. Also Fukui (Reference Fukui1990) indicated that the cumulus cells are necessary for maximize the incidence of acrosome reaction in bovine frozen spermatozoa, as they compare the results obtained from denuded oocytes. In human, De Jonge et al. (Reference De Jonge, Rawlins and Zaneveld1988) observed higher percentages of viable acrosome reacted spermatozoa when the spermatozoa were incubated in presence of COCs than in its absence, while Plachot et al. (Reference Plachot, Mandelbaum and Junca1984) did not find differences. Nevertheless, we have to consider again that both research groups observed the acrosome reaction incidence at the end of the gamete co-culture (at 16–17 hpi).
In conclusion, in our conditions, the optimum sperm concentration is 4 × 106 sperm/ml, as this sperm:oocyte ratio (approximately 28,000) allowed us to obtain the highest penetration rate, without increasing polyspermy incidence. Likewise, we obtained the maximum penetration rate at 12 h of co-incubation, suggesting that a gamete co-culture of 12–14 h would be sufficient to obtain the highest rate of penetration, reducing the exposure of oocytes and embryos to high levels of reactive oxygen species produced by spermatozoa, especially when a high sperm concentration is used to increase the IVF outcome. Even there is not a significant correlation between the penetration and the incidence of acrosome reaction, it would be worthy to note that the highest percentage of viable acrosome-reacted spermatozoa was observed between 8–12 h of gamete co-culture, coinciding with the highest penetration rate. Finally, the presence of COCs seems to favour the acrosome reaction of free spermatozoa on IVF medium.
Acknowledgments
This study was supported by Generalitat de Catalunya and CICYT.






