


Introduction
Somatic cell nuclear transfer (SCNT) is now used to produce transgenic and/or non-transgenic pigs in several laboratories (e.g. Schmidt et al., Reference Schmidt, Kragh, Li, Du, Lin, Liu, Bogh, Winther, Vajta and Callesen2010; Ahn et al., Reference Ahn, Kim, Kim, Lee, Heo, Kang, Kang, Lee, Nho, Hwang, Woo, Park, Park and Shim2011; Jakobsen et al., Reference Jakobsen, Li, Kragh, Moldt, Lin, Liu, Schmidt, Winther, Schyth, Holm, Vajta, Bolund, Callesen, Jørgensen, Nielsen and Mikkelsen2011; Luo et al., Reference Luo, Li, Liu, Lin, Du, Li, Yang, Vajta, Callesen, Bolund and Sørensen2011; Sommer et al., Reference Sommer, Estrada, Collins, Bedell, Alexander, Yang, Hughes, Mir, Gilger, Grob, Wei, Piedrahita, Shaw, Petters and Zhang2011). The average efficiency is, however, still low and quite varied, both in the cloning procedure and in the viability of the resulting embryos, fetuses and piglets. Therefore, studies are still needed urgently to optimize the nuclear transfer procedure.
The source of oocytes has been shown to influence the developmental competence of embryos considerably (Hyun et al., Reference Hyun, Lee, Kim, Kim, Lee, Kim, Lee, Lim, Kang, Lee and Hwang2003, Ikeda & Takahashi, Reference Ikeda and Takahashi2003; Chen et al., Reference Chen, Zhang, Jiang, Liu, Schatten, Chen and Sun2006). In addition, for SCNT in pigs, nearly all studies have utilized pre-pubertal gilts as the cytoplast donor (Lee et al., Reference Lee, Kim, Hyun, Lee, Jeon, Nam, Jeong, Kim, Kim, Han, Ahn, Kang, Lee and Hwang2005; Nottle et al., Reference Nottle, Beebe, Harrison, McIlfatrick, Ashman, O'Connell, Salvaris, Fisicaro, Pommey, Cowan and d'Apice2007; Kurome et al., Reference Kurome, Ishikawa, Tomii, Ueno, Shimada, Yazawa and Nagashima2008; Park et al., Reference Park, Choi, Hong, Han, Yoo, Jin, Seol and Park2008; Cho et al., Reference Cho, Hwang, Choi, Bui, Nguyen, Park, Kim and Kim2009), while very few studies have used sow oocytes (e.g. Schmidt et al., Reference Schmidt, Kragh, Li, Du, Lin, Liu, Bogh, Winther, Vajta and Callesen2010). However, studies have indicated a limited success using pre-pubertal oocytes as recipient cytoplast for SCNT (Kuhholzer et al., Reference Kuhholzer, Hawley, Lai, Kolber-Simonds and Prather2001; Hyun et al., Reference Hyun, Lee, Kim, Kim, Lee, Kim, Lee, Lim, Kang, Lee and Hwang2003; Ikeda & Takahashi, Reference Ikeda and Takahashi2003) or in vitro fertilization (IVF; Bagg et al., Reference Bagg, Nottle, Grupen and Armstrong2006), and a decreased rate of fertilization and viability was observed in embryos that originated from oocytes collected from pre-pubertal gilts compared with sows (Hyun et al., Reference Hyun, Lee, Kim, Kim, Lee, Kim, Lee, Lim, Kang, Lee and Hwang2003). Even though it is more convenient and relatively easy to get high numbers of gilt oocytes from the slaughterhouse, there could be biological reasons to prefer sow oocytes as they originate from sexually mature animals. Therefore, more work is needed to identify ways in which to improve the development of cloned embryos after the use of gilt oocytes.
Many factors influence the in vitro maturation (IVM) of oocytes, including type of medium applied, duration of maturation time and co-culture with follicular cells (Hong et al., Reference Hong, Yong, Lee, Hwang, Lim and Lee2004; Holker et al., Reference Holker, Petersen, Hassel, Kues, Lemme, Lucas-Hahn and Niemann2005). Furthermore, embryo development after SCNT was affected by different IVM media or other changes used for oocyte maturation (Abeydeera et al., Reference Abeydeera, Wang, Cantley, Rieke and Day1998a; Ikeda & Takahashi, Reference Ikeda and Takahashi2001; Miyoshi et al., Reference Miyoshi, Rzucidlo, Pratt and Stice2003). Co-culture as a positive stimulus was used in several other contexts, e.g. co-culture of reconstructed embryos with follicle shells in miniature pig (Hoshino et al., Reference Hoshino, Uchida, Shimatsu, Miyake, Nagao, Minami, Yamada and Imai2005), and co-culture of oocytes with follicular shell pieces during in vitro maturation (Song & Lee, Reference Song and Lee2007), so we suggested that it might be helpful to culture gilt oocytes together with sow oocytes.
Therefore, the aim of the present study was to identify parameters to increase the cloning efficiency using oocytes from pre-pubertal (gilt) versus postpubertal (sow) pigs.
Materials and methods
In vitro maturation of oocytes
In vitro oocyte maturation was performed as described previously (Kragh et al., Reference Kragh, Vajta, Corydon, Purup, Bolund and Callesen2004; Du et al., Reference Du, Kragh, Zhang, Li, Schmidt, Bogh, Zhang, Purup, Jorgensen, Pedersen, Villemoes, Yang, Bolund and Vajta2007; Li et al., Reference Li, Villemoes, Zhang, Du, Kragh, Purup, Xue, Pedersen, Jorgensen, Jakobsen, Bolund, Yang and Vajta2009). Briefly, cumulus–oocyte complexes (COCs) were aspirated from 2–6 mm follicles of slaughterhouse-derived gilt or sow ovaries with an 18-gauge needle using vacuum suction, while the gilt ovaries were collected from pigs 4–6 months of age without a corpus luteum. COCs were selected according to their morphological characteristics, with at least three layers of compact cumulus and even cytoplasm. After washing twice in HEPES-buffered Tissue Culture Medium 199 (TCM-199, Invitrogen) plus 0.3% heparin, 5% amphotericin and 10% cattle serum (CS; Danish Veterinary Institute, DTU, Frederiksberg, Denmark), the COCs were cultured in groups of 50–60 in 4-well dishes (Nunc, Roskilde, Denmark) with bicarbonate-buffered TCM-199 supplemented with 10% (v/v) CS, 10% (v/v) gilt follicular fluid (Gilt IVM medium) or 10% (v/v) sow follicular fluid (Sow IVM medium) according to the Experimental Designs, 10 IU/ml eCG and 5 IU/ml hCG (Suigonan Vet; Intervet, Skovlunde, Denmark) at 38.5°C with 5% CO2 in air with maximum humidity in the incubator.
Somatic cell nuclear transfer
Porcine adult fibroblast monolayer cultures were established and prepared from Danish large white pigs as described previously (Kragh et al., Reference Kragh, Vajta, Corydon, Purup, Bolund and Callesen2004). On the morning of the cloning day, the trypsinized cells were transferred into an Eppendorf tube with 400 μl T2 to be centrifuged for 3 min at 3000 rpm. Until fusion, the cells were stored in T2 at room temperature (22–25°C).
For SCNT, hand-made cloning (HMC) technology was applied (Li et al., Reference Li, Du, Zhang, Kragh, Purup, Bolund, Yang, Xue and Vajta2006; Du et al., Reference Du, Kragh, Zhang, Li, Schmidt, Bogh, Zhang, Purup, Jorgensen, Pedersen, Villemoes, Yang, Bolund and Vajta2007; Kragh et al., Reference Kragh, Nielsen, Li, Du, Lin, Schmidt, Bogh, Holm, Jakobsen, Johansen, Purup, Bolund, Vajta and Jorgensen2009). Briefly, the COCs were aspirated and matured in vitro as described above. The cumulus cells of matured COCs were removed with 1 mg/ml hyaluronidase. After partial digestion of the zona pellucida with 3.3 mg/ml pronase in T33, in each batch 15–20 of oocytes were lined up in a T2 drop supplemented with 2.5 mg/ml cytochalasin B (CB). Oriented bisection, according to the polar body (PB) position, was performed manually with a microblade (AB Technology, Pullman, WA, USA) under a stereomicroscope. The halves without PB were selected as cytoplasts and transferred into a T2 drop for further fusion and activation.
A two-step fusion procedure was performed with the second fusion including the initiation of activation (Kragh et al., Reference Kragh, Vajta, Corydon, Purup, Bolund and Callesen2004; Li et al., Reference Li, Du, Zhang, Kragh, Purup, Bolund, Yang, Xue and Vajta2006). For the first fusion step, approximately 200–400 somatic cells were placed into a T2 drop. By using a finely drawn glass pipette, each cytoplast was transferred to 1 mg/ml of phytohemagglutinin (PHA) for 2–3 s, and then dropped over a single somatic cell. After attachment, the equilibrated cytoplast–fibroblast pairs in the fusion medium (0.3 M mannitol, 0.1 mM MgSO4 and 0.01% (w/v) polyvinyl alcohol (PVA)) were aligned to one wire of a fusion chamber (BTX microslide 0.5 fusion chamber, model 450, BTX, San Diego, CA, USA) by using an alternating current (AC) of 0.06 kV/cm and with the fibroblast farther away from the wire. Pairs were then fused in the fusion medium with a direct current (DC) impulse of 2.0 kV/cm for 9 μs. After 1 h incubation in a T10 drop, fused pairs were selected and equilibrated in the activation medium (0.3 M mannitol, 0.1 mM MgSO4, 0.1 mM CaCl2 and 0.01% PVA). By using a single DC pulse of 0.86 kV/cm for 80 μs, each pair was fused with another cytoplast and activated in activation medium. Fusion was checked again after 15–20 min in the T10 drop.
For post-activation, reconstructed embryos were transferred into a Nunc 4-well dish with 400 μl porcine zygote medium 3 (PZM-3; Yoshioka et al., Reference Yoshioka, Suzuki, Tanaka, Anas and Iwamura2002) supplemented with 5 μg/ml CB, 10 μg/ml cycloheximide (CX) and covered with 400 μl mineral oil. The embryos were chemically activated for 4 h at 38.5°C in 5% CO2, 5% O2 and 90% N2 with maximum humidity.
After chemical post-activation and after twice washing in PZM-3, all activated embryos were further incubated individually in the Well-of-the-Well (WOW) system (Vajta et al., Reference Vajta, Peura, Holm, Paldi, Greve, Trounson and Callesen2000) made in Nunc 4-well dishes filled with 400 μl PZM-3 medium covered with 400 μl mineral oil at 38.5°C in 5% CO2, 5% O2 and 90% N2 with maximum humidity in either a standard incubator (Heraeus Cell 240) or in an EmbryoScope with time-lapse facilities (Unisense A/S, Skejby, Denmark).
Experimental design
The present work was carried out over a period of several months, but both before and during this period our laboratory performed SCNT (Kragh et al., Reference Kragh, Nielsen, Li, Du, Lin, Schmidt, Bogh, Holm, Jakobsen, Johansen, Purup, Bolund, Vajta and Jorgensen2009, Li et al., Reference Li, Villemoes, Zhang, Du, Kragh, Purup, Xue, Pedersen, Jorgensen, Jakobsen, Bolund, Yang and Vajta2009; Schmidt et al., Reference Schmidt, Kragh, Li, Du, Lin, Liu, Bogh, Winther, Vajta and Callesen2010) based on sow oocytes almost every week. Because of this regular use and the relative stability of the results obtained, this system served as the overall control for the results from the experimental work.
In brief, our control was the HMC system described above, i.e. in vitro maturation of sow oocytes, maturation medium supplemented with 10% sow follicular fluid for 42 h, matured oocytes with first PB directly used for HMC with bisection and fusion of the halves to the same fibroblast cells as used in the experimental work.
In all experiments, the developmental ability of the reconstructed embryos was evaluated on day 2 and day 6 after day of cloning (day 0) based on cleavage and blastocyst rates, respectively.
Experiment 1: Gilt oocytes – different cytoplasmic volume of cloned embryos
Gilt and sow oocytes were aspirated (Fig. 1) and standard matured (42 h in Sow IVM medium), and those with PB were selected for HMC. After the hand-held bisection, it is always found that the resulting cytoplasts are of different sizes, so the selected putative cytoplasts were sorted under stereomicroscope into two groups according to their size (large, small). These two cytoplasmic sizes were then used in three different combinations during the first and second fusion procedures of HMC to create cloned embryos with different cytoplasmic volume: large + large (L + L), large + small (L + S) or small + small (S + S) (Fig. 2), resulting in reconstructed embryos with a cytoplasmic volume being larger than, same as or smaller than the cytoplasmic volume of the normal oocyte, respectively.

Figure 1 Sow and gilt ovaries collected from slaughter house. (A) Sow ovaries. (B) Gilt ovaries. Scale bar indicates 1 cm.
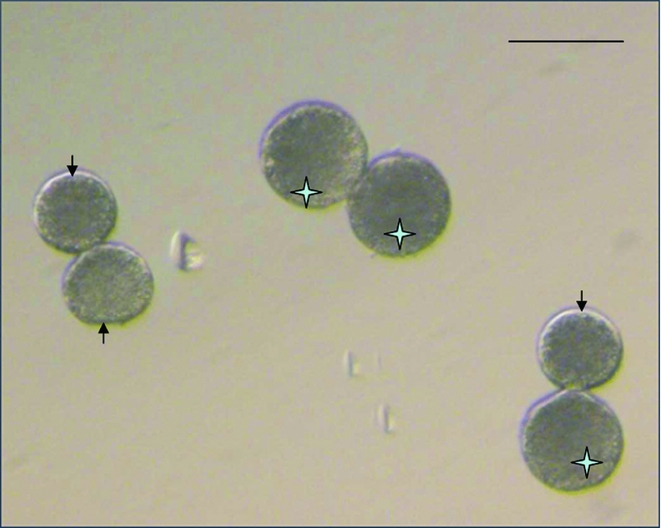
Figure 2 Two cytoplasmic sizes used in three different combinations during the hand-made cloning (HMC) procedure to create cloned embryos with different cytoplasmic volume. The star indicates a large cytoplast; an arrow indicates a small cytoplast; and the scale bar indicates 100 μm.
Experiment 2: Gilt oocytes – extended IVM time
Gilt oocytes were standard matured in Sow IVM medium, but for an extended time of 48 h. This longer interval should provide additional time for completion of cytoplasmic maturation of the oocyte. The matured oocytes with PB were then selected for HMC.
Experiment 3: Gilt and sow oocytes – co-culture (during IVM) in different proportions
Gilt and sow oocytes were standard matured (42 h, Sow IVM medium). Three groups were made with 50 oocytes in different gilt:sow proportions: 20% gilt (10 gilt + 40 sow oocytes), 50% gilt (25 gilt + 25 sow oocytes) and 80% gilt (40 gilt + 10 sow oocytes). In each group, all matured oocytes with PB were selected to be used for HMC.
Experiment 4: Gilt oocytes – in vitro maturation in gilt medium and cloning with oocytes of different sizes
Gilt oocytes were standard matured (42 h), but in Gilt IVM medium. After maturation, just after COCs removal and before bisection, the matured oocytes with PB were divided into three groups according to their sizes determined under stereomicroscope: large oocytes, small oocytes or non-sorted oocytes as control. The diameter of the oocyte was determined by the EmbryoScope's software, measuring an average distance of two measurements from the ooplasmic membrane under the zona pellucida passing through the centre of the oocyte. From the three groups, all matured oocytes with PB were selected to be used for HMC.
Experiment 5: Gilt oocytes – transfer of cloned embryos
Gilt oocytes were standard matured (42 h) in Gilt IVM medium. The matured oocytes with PB were sorted as described in Experiment 4, so only large oocytes were used for HMC. On day 0, a team of 4–5 people in our laboratory performed the cloning work to collect sufficient numbers of cloned embryos for further in vitro culture and subsequent transfer. On days 5 or 6, a mixture of blastocysts and morulae with good morphology (Fig. 3) were transferred surgically to both cornua uteri of Danish Landrace sows 5 days after weaning (Schmidt et al., Reference Schmidt, Kragh, Li, Du, Lin, Liu, Bogh, Winther, Vajta and Callesen2010). Pregnancies were diagnosed by ultrasonography on day 28, and at term piglets were delivered naturally.

Figure 3 Blastocysts and morula with good morphology used for transferring into the sow recipient. (A) Blastocyst. (B) Morula. Scale bar indicates 100 μm.
Data analysis
The data were analysed by one-way analysis of variance (ANOVA) analysis, and a probability of P < 0.05 was considered to be statistically significant.
Results
In Table 1, the same overall control results are described at the bottom line for each of the five experiments. These results were made from 10 replicates performed during the same weeks of the five experiments.
Table 1 Developmental ability of gilt and sow embryos reconstructed with somatic cell nuclear transfer

N indicates the number of replicates. n indicates the number of embryos being observed.
At each experiment rows, different superscripts in the same column show significant differences. a,b,c (P < 0.05); * indicates the difference when compared with the result (L + S) from Experiment 1. Mean ± SEM (standard error of mean). See ‘Experimental design’ for further details.
Experiment 1: Gilt oocytes – different cytoplasmic volume of cloned embryos (Table 1)
With gilt oocytes, the cleavage rates were only reduced compared with the control sow oocytes when the L + L combination of gilt cytoplasts was used (96.0 versus 80.3%, respectively). However, all blastocyst rates were lower when using gilt compared with sow oocytes, and within the experimental groups the S + S combination was almost half the blastocyst rate compared with the other two combinations (14.8% versus 25.8% (L + S) and 31.4% (L + L)).
Experiment 2: Gilt oocytes – extended IVM time (Table 1)
No difference was found in cleavage rates when using gilt oocytes matured for 48 h versus both gilt oocytes matured for 42 h (Experiment 1) and control sow oocytes. Also as found in Experiment 1, the blastocyst rate with gilt oocytes matured for 48 h was approximately half the rate for control sow oocytes.
Experiment 3: Gilt and sow oocytes – co-culture in different proportions (Table 1)
No difference was found in cleavage rates when using co-culture of gilt and sow oocytes in all three proportions or using only sow oocytes (control) or gilt oocytes (Experiment 1). Also as found in Experiment 1, the blastocyst rates with co-culture of gilt and sow oocytes in all three proportions were reduced compared with the rate for control sow oocytes, but it was increased compared with the rates for gilt cytoplasts (Experiment 1).
Experiment 4: Gilt oocytes – in vitro maturation in gilt medium and cloning with oocytes of different sizes (Table 1)
The diameter of oocytes in the three categories were: (a) large oocytes: long diameter 110.6 ± 1.6 μm, short diameter 106.6 ± 1.6 μm (n = 12); (b) small oocytes: long diameter 103.8 ± 0.8 μm, short diameter 97.9 ± 1.0 μm (n = 12); and (c) non-sorted oocytes (control): long diameter 106.4 ± 1.1 μm, short diameter 101.3 ± 1.3 μm (n = 12) (Fig. 4). Within all matured oocytes, the overall proportion of large oocytes and small oocytes was around 60–70% and 30–40%, respectively.

Figure 4 Measurement of the diameter of oocytes by EmbryoScope with time-lapse facilities. (A) Large oocyte with a diameter of 110 μm. (B) Small oocyte with a diameter of 102 μm.
No differences were found in cleavage rates compared with the control sows or the relative result in Experiment 1. However, blastocyst rates were the highest for large oocytes compared with small and non-sorted cells (42% versus 30% and 26%, respectively). This highest result was however still reduced compared with the control sows.
Experiment 5: Gilt oocytes – transfer of cloned embryos (Tables 2, 3 and 4)
Table 2 Production of cloned pig embryos for transfer

D6 efficiency indicates no. of both blastocysts and morula on day 6/no. of cloned embryos.
D5 efficiency indicates no. of both blastocysts and morula on day 5/no. of cloned embryos.
Table 3 Development of the cloned embryos after transfer into sow recipients

Table 4 Piglet's birth weight (kg) and health status at delivery

*Indicates that the piglets were dead during delivery or were killed because of weakness immediately after delivery.
As shown in Table 2, of the 820–1040 oocytes used for each cloning day, 248–335 reconstructed embryos in total were produced per day. After culture in vitro, there was a higher variation in embryos selected on day 5 (25–42%) than on day 6 (36–40%).
As shown in Table 3, in total 503 mixed day 5 and day 6 blastocysts and morulae developed from gilt oocytes were transferred to four sow recipients, while two sow recipients had received, in total, 190 mixed day 5 and day 6 blastocysts and morula developed from sow oocytes. All six recipients became pregnant (100%) and gave birth to piglets with a litter size from three to 12, with a higher variation in the birth weights in litters from gilt than from sows (Table 4). No abnormalities were observed on physical examination of the live piglets (Fig. 5).

Figure 5 Piglets at 6 days after birth from the cloned embryos reconstructed with gilt oocytes.
Discussion
The general finding of this work was a confirmation that the use of gilt oocytes for nuclear transfer results in reduced blastocyst rates compared well with the use of sow oocytes. However, our study showed results that were strongly optimistic for the future use of gilt oocytes in this work. One quite significant finding was the achievement of 100% pregnancy rates after transfers and birth of piglets, based on use of gilt oocytes. A second finding was that size of oocytes after in vitro maturation and of cytoplasts after enucleation resulted in significantly improved blastocyst rates. The third finding was that there was a positive effect after use of gilt oocytes for nuclear transfer if these oocytes were cultured together with different proportions of sow oocytes.
As the recipient cytoplasm exerts a key role of chromatin remodelling, the effect of the cytoplasmic volume after SCNT cannot be neglected. The removal of 25–50% of the ooplasm during enucleation compromised embryo development and embryo quality (Peura et al., Reference Peura, Lewis and Trounson1998; Koo et al., Reference Koo, Kang, Choi, Park, Kim, Oh, Son, Park, Lee and Han2002). We constructed SCNT embryos with different cytoplasmic volumes using gilt oocytes, but no improvement in development was observed compared with sow oocytes. However, we did observe a negative effect if the total cytoplasmic volume was too small. These findings might indicate that the compromised developmental ability of SCNT embryos originating from gilt oocytes is caused by cytoplasmic immaturity rather than the volume of the reconstructed embryos.
The maturation period of recipient oocytes may have significant effects on the developmental capacity of pig cloned embryos (Holker et al., Reference Holker, Petersen, Hassel, Kues, Lemme, Lucas-Hahn and Niemann2005). It is common to mature pig oocytes in vitro for 40–44 h before manipulation (Tao et al., Reference Tao, Machaty, Boquest, day and Prather1999; Holker et al., Reference Holker, Petersen, Hassel, Kues, Lemme, Lucas-Hahn and Niemann2005; Song & Lee, Reference Song and Lee2007). Most of the 42 h matured oocytes were found to have been aged at metaphase II (MII) stage for at least 12 h (Miyoshi et al., Reference Miyoshi, Rzucidlo, Pratt and Stice2002) and the completion of the oocyte's cytoplasmic maturation needed additional culture time after reaching the MII stage (Ikeda & Takahashi, Reference Ikeda and Takahashi2003). In pigs, immature oocytes can be fully matured in vitro after 38 h, but oocytes expel their first PB over a wide range of maturation times (Ikeda & Takahashi, Reference Ikeda and Takahashi2001; Miyoshi et al., Reference Miyoshi, Rzucidlo, Pratt and Stice2002). On the other hand, the activity of maturation/M-phase promoting factor (MPF) was affected by the age of the oocyte, and the MPF was increased during the process of oocyte maturation and remained at a high level during meiotic arrest. However, inactivation of MPF and prematuration of oocytes resulted in a decreased developmental competence of SCNT embryos after 42 h of pig oocytes maturation. Therefore, we extended the IVM duration time to 48 h, but no significant difference was observed. The reason might be that MPF activity in aged oocytes gradually decreased, and that the fragmentation frequency gradually increased (Kikuchi et al., Reference Kikuchi, Nagai, Ding, Yamauchi, Noguchi and Izaike1999; Reference Kikuchi, Naito, Noguchi, Kaneko and Tojo2002).
As demonstrated previously the rates of cell fusion, cleavage and blastocyst formation in SCNT embryos were influenced significantly by the IVM period, such as co-culture with follicular cells during IVM (Liu et al., Reference Liu, Dai and Moor1997; Kidson et al., Reference Kidson, Schoevers, Langendijk, Verheijden, Colenbrander and Bevers2003; Hoshino et al., Reference Hoshino, Uchida, Shimatsu, Miyake, Nagao, Minami, Yamada and Imai2005) or with follicular shell pieces (Abeydeera et al., Reference Abeydeera, Wang, Cantley, Rieke and Day1998a; Hoshino et al., Reference Hoshino, Uchida, Shimatsu, Miyake, Nagao, Minami, Yamada and Imai2005; Song & Lee, Reference Song and Lee2007). This situation might be because the follicular cells surrounding oocytes secrete specific proteins required for cytoplasmic maturation (Liu et al., Reference Liu, Dai and Moor1997). Our results showed that when different proportions of gilt and sow oocytes were co-cultured during IVM, the developmental capacity of the cloned embryos with these mixed gilt and sow oocytes could be improved – no matter what was the ratio between gilt and sow oocytes. The reasons for this outcome are so far unknown, but we postulate at least two points. First, the improved embryo development after co-culture with sow oocytes may be caused by promoting substances contained in sow oocytes during maturation. Secondly, during HMC, the two half parts of the cytoplasts are used to fuse with one donor cell, and these half parts might come from two oocyte types, such that the promoting factors in the sow cytoplasm could be helpful for reprogramming the donor cells and furthermore the development of the cloned embryos.
Follicular fluid contains both stimulatory and inhibitory factors that affect oocyte development (Wiesak et al., Reference Wiesak, Hunter and Foxcroft1990; Dostal & Pavlok, Reference Dostal and Pavlok1996; Choi et al., Reference Choi, Takagi, Kamishita, Wijayagunawardane, Acosta, Miyazawa and Sato1998). Oocytes are matured in vivo through mutual interaction of oocytes, follicular cells and their surrounding environment including the follicular fluid. Various factors such as growth factors and hormones (Byskov et al., Reference Byskov, Yding, Hossaini and Guoliang1997; Makarevich et al., Reference Makarevich, Sirotkin and Genieser2004) are expected to influence oocyte maturation and embryos development after IVF (Abeydeera et al., Reference Abeydeera, Wang, Cantley, Rieke, Prather and Day1998b) or SCNT (Hoshino et al., Reference Hoshino, Uchida, Shimatsu, Miyake, Nagao, Minami, Yamada and Imai2005). In addition, even from the same aged pig, the developmental competence of oocytes could be affected by different batches of ovaries (Song & Lee, Reference Song and Lee2007). Therefore, we postulate that different factors are present in gilt and sow follicular fluid. The maturation of the gilt oocytes might benefit from supplementation with gilt follicular fluid. However, when we matured gilt oocytes in IVM medium supplemented with gilt instead of sow follicular fluid, we observed no difference in developmental ability of the SCNT embryos. This finding might indicate that gilt follicular fluid has a similar function for the ooplasmic maturation as the sow follicular fluid.
Of much more importance is, however, the high frequency of the smaller size of matured gilt oocytes (around 30–40%) compared with matured sow oocytes. When we sorted the matured gilt oocytes into large and small oocytes and used the large matured gilt oocytes as cytoplasmic recipients, the development capacity of the cloned embryos was improved. This situation might indicate that the large matured gilt oocytes were more matured than the small oocytes. One reason for the low efficiency when gilt oocytes were used for SCNT could be the high proportion of the small and non-matured gilt oocytes. Therefore, a possible way to use pre-pubertal gilt oocytes to produce cloned embryos is to select the matured large oocytes according to their morphology. Furthermore, in this study, naturally cycling, mature sows were used as recipients, as used routinely in our group (Schmidt et al., Reference Schmidt, Kragh, Li, Du, Lin, Liu, Bogh, Winther, Vajta and Callesen2010). The four sow recipients that received cloned embryos from gilt oocytes gave birth to 26 piglets in total with an average birth weight of 1.34 kg, while the two recipients that received cloned embryos from sow oocytes gave birth to six piglets in total with an average birth weight of 1.81 kg.
This study demonstrates that cloned embryos that originate from matured gilt oocytes have a compromised development compared with their counterparts that originate from matured sow oocytes. However, according to a selection based on their morphology and an improved maturation regime, the efficiency to produce piglets by in vitro matured gilt oocytes could be increased. In conclusion, the efficiency of the production of piglets using in vitro matured gilt oocytes can be as high as that of cloned piglets when using sow oocytes.
Acknowledgements
The authors thank A. Pedersen, K. Villemoes, R. Kristensen and A.K. Nielsen for invaluable technical assistance.
The study was supported by grants from the ‘Pigs & Health’ project (Danish Advanced Technology Foundation no. 013–2006–2), the ‘DAGMAR’ project (the Danish National Research Infrastructures Programme no. 09–065333) and the project support by the Fundamental Research Funds for the Central Universities (The Ministry of Finance and the Ministry of Education of China, no. KYZ 201115).









