Weed management continues to be a high priority for organic farmers (Jerkins and Ory Reference Jerkins and Ory2016), whose fields typically have greater weed abundance and species diversity than the fields of comparable conventional farmers (Graziani et al. Reference Graziani, Onofri and Pannacci2012; Rasmussen and Ascard Reference Rasmussen and Ascard1995). Organic farmers often rely on tillage and cultivation to control weeds, which can degrade soil health and facilitate soil erosion (Peigné et al. Reference Peigné, Ball, Roger‐Estrade and David2007; Gruber and Claupein Reference Gruber and Claupein2009; Smith et al. Reference Smith, Ryan and Menalled2011). Organic vegetable farmers typically use more intensive primary tillage (rototiller vs. moldboard plow) and more frequent postplanting cultivation than organic grain farmers. This is because some vegetable crops require a fine seedbed (e.g., carrots [Daucus carota L. var. sativus Hoffm.], baby greens, other vegetable crops [e.g., onions (Allium spp.)]) and have longer critical periods for weed control than grain crops (Tursun et al. Reference Tursun, Bükün, Karacan, Ngouajio and Mennan2007), and because short-season vegetable crops enable planting and harvesting more than one crop in each growing season (Mohler and Johnson 2009). Also, rotary tillage is well suited for preparing a seedbed in small areas on a diversified vegetable farm.
Cultural management practices such as diversifying crop rotations, integrating cover crops, and using practices that increase the relative competitive ability of crops (e.g., high crop seeding-rate density, narrow row spacing, and competitive varieties) can contribute to weed suppression and reduce the need for cultivation (Bastiaans et al. 2008; Liebman et al. 2001). Such practices not only decrease competition from weeds but also reduce weed fecundity, which affects the impact of weeds on future crops. Most direct weed control practices target the weed seedling stage, but practices specifically aimed at weed seed management, such as preventing seed rain and enhancing postdispersal weed seed mortality, can reduce losses from weeds and the cost of weed management (Gallandt Reference Gallandt2006). Seeds can also be flushed from the seedbank by creating a false seedbed and then using shallow tillage to terminate weed seedlings before planting the crop. This practice can reduce competition from weeds and reduce the need for precision cultivation and hand hoeing.
Long-term cropping system research is important for understanding the cumulative effects of different management practices, especially in organic cropping systems where incomplete weed suppression in one year can lead to weed population spikes the next year. Soil, crop, and pest management practices can impact weed population and community dynamics, and their effects can vary with weather. These interactions make drawing robust conclusions from short-term field experiments difficult. Several long-term cropping system experiments have compared organic grain production practices, but far fewer have focused on vegetable production. Information from long-term vegetable experiments could inform the development of sustainable weed management practices for organic vegetable production.
Weeds in annual crops originate primarily from the weed seedbank (Cavers and Benoit Reference Cavers and Benoit1989; Fennimore and Jackson Reference Fennimore and Jackson2003), which, because of seed survival and persistence in the soil, reflects the effects of previous management on weed growth and reproduction (Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010; Smith and Gross Reference Smith and Gross2006; Swanton and Booth Reference Swanton and Booth2004). The weed seedbank can be sampled and analyzed to determine the effects of past management on weed populations and communities. Uniformity trials are another tool for evaluating the cumulative effect of management practices on weed populations and communities (Teasdale et al Reference Teasdale, Coffman and Mangum2007; Schipanski et al. 2017). A uniformity trial consists of growing the same crop using the same management over all treatments in a field experiment in order to evaluate the legacy effects on crop performance and weed communities.
The Cornell Organic Vegetable Cropping Systems Experiment (OVCS) was established in 2004 to compare four crop management systems characterized by different practices. In 2014 a uniformity trial was conducted to evaluate the legacy effects of these long-term management systems on weed abundance and community composition. An economic analysis of the first 4 yr showed that different systems maximized returns for different crops (Chan et al. Reference Chan, Caldwell, Rickard and Mohler2011). Here we report on the results of a soil weed seedbank emergence bioassay and a series of uniformity trials after 10 yr of different organic vegetable management practices. We hypothesized that weed abundance and community composition would differ among the cropping systems.
Materials and Methods
Study Site and Experimental Design
The OVCS was established in 2004 at the Homer C. Thompson Vegetable Research Farm near Freeville, NY (42.52°N, 76.33°W). The experiment used a split-plot randomized complete block design with four replications. The main plots were cropping systems, representing four management strategies: (1) intensive, (2) intermediate, (3) bio-extensive, and (4) ridge tillage. Subplots (n=32) consisted of two entry points into the crop rotation so that the same crop in the sequence was grown in two successive years in adjacent subplots. Multiple entry points into the crop rotation is common in cropping system studies (Davis et al. Reference Davis, Hill, Chase, Johanns and Liebman2012; Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010; Smith et al. Reference Smith, Gross and Robertson2008) and allowed evaluation of the effects of weather on crop production and ecological processes. The subplots were 19.8-m long and 6.0-m wide with a 1.5-m buffer between plots. The soil was a Howard gravelly loam (loamy-skeletal, mixed, active, mesic Glossic Hapludalfs) and a Phelps gravelly silt loam (Fine-loamy over sandy or sandy-skeletal, mixed, active, mesic Glossoboric Hapludalfs). The experimental site was located in USDA Plant Hardiness Zone 5B with a 30-yr average annual precipitation of 979 mm. During the uniformity trial period from April 28 to September 24, 2014, rainfall was 517 mm and growing degree day accumulation was 1,098 (base 10 C). Additional details of the experiment can be found in Chan et al. (Reference Chan, Caldwell, Rickard and Mohler2011).
Cropping Systems
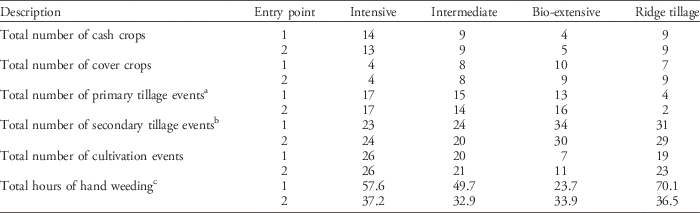
The four cropping systems varied in cash crop frequency, weed management strategy, cover crop frequency, fertilizer inputs, and tillage methods (Table 1). Cultivation and hand weeding were used in all systems. Hand hoeing in the row area was done with a stirrup hoe when weeds were small (<8 cm). Hoeing was standardized across systems by hoeing until weeds were removed. Large weeds (>20 cm) were hand rogued in all systems, and additional hand roguing was done in the bio-extensive system to prevent seed production. Labor associated with hand weeding in each system was recorded (Table 2).
Table 1 Summary of management differences across the four cropping systems.

The intensive cropping system simulated a farm with limited land that focused on maximizing crop production and income per hectare. Over a 4-yr rotation period, six cash crops were grown in this system. The weed management strategy was to minimize weed–crop competition, but no additional effort was made to limit weed seed production. Weeds were controlled primarily by cultivation and hand hoeing. Compost with a test of 1.9-1.4-0.7 (N- P2O5- K2O) was the primary nutrient source, typically applied at an average rate of 6.25 Mg ha−1 year−1.
The intermediate cropping system also replicated a land-limited farm but endeavored to obtain most of its nitrogen from legume cover crops. One cash crop was grown annually. The weed management strategy was similar to the intensive system, with additional weed management when time allowed. About half the nitrogen for crop production was supplied by legume cover crops, and compost was applied at an average rate of 3.13 Mg ha−1.
The bio-extensive cropping system was modeled after Eric and Anne Nordell’s farm, which uses cover crops and tilled fallow periods in alternate years to reduce the weed seedbank and build soil health (Gallandt Reference Gallandt2006). Two cash crops were grown over a 4-yr time period. The weed management strategy was to deplete the weed seedbank by preventing seed production with added hand labor and tillage and flushing seeds out of the seedbank with tilled fallows. Fertilizer inputs were the same as the intermediate system, except that in years without cash crops, no compost was applied. Tillage was reduced in this system through shallow plowing rotary tillage, and harrowing.
The ridge-tillage cropping system used ridge tillage and controlled traffic to reduce soil disturbance and compaction. One cash crop was grown annually. The weed management strategy and fertilizer inputs were similar to the intermediate system. Ridges in fixed locations were built up after planting cover crops in the fall and scraped prior to planting cash crops to remove weeds and cover crops from the planting area. Interrows were cultivated. Ridge bases were undisturbed, except for shallow disturbance during potato digging.
Winter squash, cabbage, lettuce, and potato were grown in all systems, except that cabbage and potato were replaced by tilled fallow and cover crops in the bio-extensive system. The intensive system also had snap pea (Pisum sativum var. macrocarpon L.) before cabbage and spinach (Spinacia oleracea L.) after lettuce to increase the cropping intensity. This crop rotation was followed throughout the experiment, except that the first crop was sweet corn (Zea mays L.) instead of winter squash. The last crops grown in 2013 were cabbage in entry point 1 and winter squash in entry point 2. Over the duration of the experiment, cropping systems differed in the total number of cash crops, cover crops, tillage events, and cultivation events, and the amount of hand hoeing (Table 2).
Table 2 The total number of cash crops, cover crops, tillage events, cultivation events, and hours spent hoeing and hand weeding in each cropping system and entry point from 2004 through 2013.

a Primary tillage events include moldboard plowing, chisel plowing, rotary tillage, and potato harvesting.
b Secondary tillage events include disking, harrowing, ridge scraping, ridge hilling, potato hilling, and use of a field cultivator but not use of a roller harrow.
c Hours of hand weeding is measured in hours per 0.04-ha plot.
In 2014, after the cropping systems had been in place for 10 yr, a series of uniformity trials were conducted. On April 28, 2014, the entire field was rotary tilled to a depth of 10 cm. Oats were then drilled in all of the plots the same day at a rate of 134 kg ha−1. The oat crop was mowed, and the residue was incorporated by moldboard plowing to a depth of 18 cm on July 18, 2014. A mix of sorghum-sudangrass (33.6 kg ha−1) and Japanese millet (16.8 kg ha−1) was then planted across all plots. As weed species differ in their emergence timing, the two uniformity trials, conducted in early spring and midsummer, provided a more thorough assessment of the weed community than a single uniformity trial conducted either in early spring or midsummer.
Data Collection
Soil Seedbank
Soil samples were collected on April 10 and April 14, 2014, prior to tillage. Twelve cores (7-cm diameter by 18-cm depth) were collected in each of the 32 subplots. Samples were taken at the turns of two parallel zigzag patterns down the length of the plot. Sampling avoided areas within 1.8 m of a plot edge.
Soil samples from each plot were bulked and then screened through 1.25-cm hardware cloth onto a 1.8-m by 1.8-m plastic sheet. Rocks were discarded. The sample for each plot was then thoroughly mixed and weighed and a 2.5-kg subsample was taken. These subsamples were used for the identification and quantification of the readily germinable (nondormant to partially dormant) seedbank using an emergence bioassay in the greenhouse (Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010). The subsamples were mixed with an equal volume of Cornell’s standard potting mix and spread in two plastic flats (28 cm by 56 cm). Flats were placed in a climate-controlled greenhouse maintained at 27/21 C (day/night). Weed seedlings were permitted to grow until a positive identification could be made and were then counted and removed. After about 10 wk, the soil samples were allowed to dry out. Three months after initial placement of the soil in flats, the dry soil was remixed. It was placed back in the flats and watered, and then new flushes of weeds were counted and removed until 6 mo after initial preparation. The soil was not stratified prior to the emergence bioassay, and thus weed species that require a period of cold temperatures to break dormancy were likely underrepresented. Weeds were identified to species level when possible, although a few taxa could only be identified to genus. Although multiple weed seedling emergence flushes were recorded in the emergence bioassay, probably some dormant seeds did not germinate and other seeds germinated but failed to emerge and thus were not observed.
Emerged Weed and Crop Biomass in Oat and Sorghum-Sudangrass Uniformity Trials
Weeds density and biomass and crop biomass were sampled simultaneously in two randomly placed 0.25-m2 quadrats per plot. Samples were collected at the oat soft dough stage on June 30, 2014, in uniformity trial 1 (oat) and on September 24, 2014, in uniformity trial 2 (sorghum-sudangrass plus Japanese millet). Weeds were counted and clipped at ground level, with each species placed in a separate paper bag. Cover crop biomass was similarly sampled. Biomass was dried for 3 d at 60 C and then weighed.
Data Analysis
We used ANOVA to test for differences in total weed seedbank density and species richness of the seedlings that emerged in the greenhouse emergence bioassay. ANOVA was also used to test for differences in (1) weed density in the oat, (2) weed biomass in the oat, (3) oat biomass, (4) weed density in the sorghum-sudangrass plus Japanese millet, (5) weed biomass in the sorghum-sudangrass plus Japanese millet, and (6) the sorghum-sudangrass plus Japanese millet biomass. ANOVA was conducted using JMP Pro 12 (SAS Institute, Cary, NC). Data were transformed as ln(x + 1) for the ANOVAs, and back-transformed means are presented. Pairwise mean comparisons were made by using the Tukey-Kramer method, and significance was declared for P≤0.05. A split-plot fixed-effects model was used with cropping system (main plot), rotation entry point (subplot), and the interaction between the cropping system and the rotation entry point as fixed effects, and the replicate (block) and the cropping system by replicate interaction as random effects.
The legacy effect of cropping system and entry point on the weed community was tested with five data sets: (1) the weed seedbank before establishing oat, (2) the weed density and (3) weed biomass in the oat, and (4) the weed density and (5) weed biomass in the sorghum-sudangrass plus Japanese millet. Weed seed or plant density and biomass data were transformed as ln(x+1) and then subjected to principal coordinates analysis (PCoA) with Bray-Curtis distance metric implemented in the ‘Vegan’ package (Oksanen et al. Reference Oksanen, Blanchet, Kindt, Legendre, O’Hara, Simpson, Solymos, Stevens and Wagner2010) in R software (R Development Core Team 2015). The significance of cropping system, entry point, and their interaction was tested with a permutation-based multivariate ANOVA (PerMANOVA; Anderson Reference Anderson2001) using the Adonis function of the ‘FactoMineR’ package (Le et al. Reference Le, Josse and Husson2008). We used PC-ORD 6 (MjM Software Design, Gleneden Beach, OR) to conduct an indicator species analysis (ISA; Dufrene and Legendre Reference Dufrene and Legendre1997) to test for associations between weed species in the weed seedbank and the four cropping systems. Indicator values for each species were calculated by multiplying the relative abundance by the relative frequency within a cropping system. Indicator values range from 0 (no association) to 100 (exclusive association with one system). Significance of indicator values was assessed using a Monte Carlo procedure (1,000 runs). We used Mantel tests (vegdist function in the ‘Vegan’ package) to test for correlations between weed communities in each of the five data sets using the Bray-Curtis dissimilarity index (Mantel Reference Mantel1967). Correlation significance was assessed using permutation tests with 1,000 permutations.
Results and Discussion
Weed Seedbank
A total of 41 taxa were recorded in the weed seedbank. The most frequently occurring species (occurrence>50%), in decreasing order, were common chickweed [Stellaria media (L.) Vill.], pigweed (Amaranthus spp. mostly Powell amaranth [A. powellii S. Wats.] and redroot pigweed [A. retroflexus L.]), hairy galinsoga (Galinsoga quadriradiata Cav.), common lambsquarters (Chenopodium album L.), oakleaf goosefoot (Chenopodium glaucum L.), large crabgrass [Digitaria sanguinalis (L.) Scop.], common purslane (Portulaca oleracea L.), and shepherd’s purse [Capsella bursa-pastoris (L.) Medik.]. Thirteen rare species were observed in only one plot, whereas common chickweed was the only species observed in all plots. Weed seedbank density ranged from an average of 3,260 seeds m−2 in the bio-extensive system in entry point 1 to 35,200 seeds m−2 in the ridge-tillage system in entry point 2.
Systems and entry points differed significantly in seedbank density, and the interaction between the system and entry point was not significant (Figure 1; Table 3). The bio-extensive system had lower weed seedbank densities than the other systems. Weed seedbank density was greater in entry point 2 than entry point 1 (Figure 1). The low weed seedbank densities in the bio-extensive system indicate that tilled fallow periods combined with hand roguing of weeds before they go to seed can reduce weed seedbanks. To comply with the goal of the bio-extensive system of zero tolerance for weed reproduction, some cover crops that might have allowed escape of mature weeds were terminated early and replanted in the early years of the experiment.

Figure 1 Mean weed seedbank density by cropping system and crop rotation entry point. Bars with the same letters are not significantly different (Tukey-Kramer, P>0.05) among cropping systems (uppercase letters) and entry points (lowercase letters).
Table 3 Significance levels from the ANOVAs performed on weed seedbank density and aboveground weed density, weed biomass, and crop biomass in the oat and the sorghum-sudangrass plus Japanese millet uniformity trials.

The differences observed between the two crop rotation entry points in the seedbank are likely due to the last crop grown in the entry points. Cabbage was the last crop in entry point 1, and winter squash was the last in entry point 2. Midseason planting of cabbage resulted in fewer weeds emerging and a shorter period for weeds to grow. Thus, the timing of planting, in addition to hand hoeing, resulted in few weed seeds being produced in cabbage. In contrast, winter squash was planted earlier, and, although the squash was also hand hoed, weeds that emerged after the vines had spread through the plots had plenty of time to grow and produced many seeds. These results are congruent with Gallandt (Reference Gallandt2006), who compared seed rain in winter squash and broccoli (Brassica oleracea L. var. botrytis) and found a large crop effect on the weed seedbank due to abundant seed rain in winter squash. Seedbank density data from elutriated soil samples that were collected in 2012 and 2013 in the OVCS suggest the difference between crop rotation entry points is likely due to management during the previous crop (unpublished data). However, total hand-hoeing labor over the duration of the experiment was also greater in entry point 1 than in entry point 2 in the intensive (55%), intermediate (51%), and ridge-tillage (92%) systems, and also could have contributed to the crop rotation entry point effect (Table 2).
Results from the PerMANOVA on the weed seedbank community data showed a significant interaction between cropping system and crop rotation entry point (Table 4). The two first axes of the PCoA accounted for 35.3% of variability in the seedbank weed community and showed a large amount of overlap in the seedbank communities in the intensive system and the intermediate system, whereas the communities of the bio-extensive and ridge-tillage system were each more distinct from all other systems (Figure 2). At the individual species level, results from the ISA (Table 5) showed that oakleaf goosefoot and fall panicum (Panicum dichotomiflorum Michx.) were associated with the intensive system, and common purslane, large crabgrass, purple deadnettle (Lamium purpureum L.), pigweed species, and common chickweed were associated with the ridge-tillage system. Purple deadnettle and common chickweed are winter annual species that often emerge in late summer. The soil disturbance used to create the ridges after cover crop seeding in the fall and the subsequent scraping of the ridges in the spring were ineffective at controlling these species, which likely set seed before interrow cultivation in the summer.

Figure 2 Principal coordinates analysis (PCoA) with Bray-Curtis distance of the weed communities of the seedbank. Dots represent plots; cropping system and entry point labels are the centroids of all species belonging to the cropping system or entry point; and ellipses show the 95% confidence intervals. Weed species (left) are named with their Bayer codes. Cropping systems (center) are: S1, intensive; S2, intermediate; S3, bio-extensive; and S4, ridge tillage. Crop rotation entry points (right) are: EP1, entry point 1; and EP2, entry point 2.
Table 4 Results of permutation-based multivariate ANOVAs assessing the effects of cropping system and crop rotation entry point on the weed community.

Table 5 Results of indicator species analysis showing weeds observed in the weed seedbank emergence bioassay that were associated with a cropping systems (P<0.1).

The PCoA showed little overlap between the two crop rotation entry points (Figure 2), which supports the results from the PerMANOVA (Table 4). Differences between the seedbank communities of the two entry points can be attributed to the last cash crop grown. Winter squash favors long-season weeds like common lambsquarters and pigweeds and warm-season species like hairy galinsoga and large crabgrass. Accordingly, these species have high loadings in the PCoA, as does entry point 2 (Figure 2). Differences between the entry points may also be due to differences in crops planted in preceding years, since crop rotation history can have a large influence on weed community composition (Leeson et al. Reference Leeson, Sheard and Thomas2000).
Oat Uniformity Trial
A total of 37 species were recorded in uniformity trial 1 (oat). Species occurring in more than 50% of the 32 plots were, in decreasing order of frequency, common chickweed, pigweed, large crabgrass, hairy galinsoga, Persian speedwell (Veronica persica Poir.), common lambsquarters, large crabgrass, and shepherd’s purse. Eight weed species were observed in only one plot, whereas common chickweed, pigweed, and large crabgrass were observed in all plots.
Weed Density in Oat
The aboveground weed density data collected during the oat uniformity trial showed significant cropping system and entry point effects (Figure 3; Table 3). The bio-extensive system had a lower weed density compared with the other systems. The relatively lower density of emerged weeds in the bio-extensive system presumably results from the lower seedbank density in that system and results from the special weed management practices in that system, as discussed earlier. Similarly, the greater weed density in entry point 2 than entry point 1 in the oat uniformity trial was due to higher seedbank density in entry point 1. As explained in the previous section, the difference in weed density between the entry points was likely due to a difference in seed production during the last crop grown.

Figure 3 Mean weed density (top), mean weed biomass (middle), and mean biomass of oat (bottom) by cropping system and crop rotation entry point in uniformity trial 1. Bars with the same letters are not significantly different (Tukey-Kramer, P>0.05) among cropping systems (uppercase letters) and entry points (lowercase letters).
The aboveground weed community composition in oat was affected by the cropping system, the entry point, and their interaction (Table 4). Not surprisingly, the differences in the emerged weed community reflected differences in the seedbank, with the intensive and intermediate systems similar to each other, and the bio-extensive and ridge-tillage systems more distinctive (unpublished data). Previous research has reported correlations between weed seedbank and aboveground weed community structure (Davis et al. Reference Davis, Renner and Gross2005; Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010; Teasdale et al. Reference Teasdale, Mangum, Radhakrishnan and Cavigelli2004).
Weed Biomass in Oat
Results for the weed biomass in oat followed the same pattern as the results for weed density (Figure 3; Table 3). Weed community composition based on the weed biomass in oat was significantly affected by the cropping system and entry point but not by their interaction (Table 4). The bio-extensive system had a significantly different weed community composition compared with the other three systems, which again can be attributed to its distinctive management practices affecting the seedbank.
Oat Biomass
Oat biomass ranged from 421 g m−2 in the intensive system in entry point 2 to 569 g m−2 in the bio-extensive system in entry point 1. Crop rotation entry point was significant, but systems did not differ (Table 3). Entry point 1 had higher oat biomass than entry point 2. Possibly, the greater weed density and biomass in entry point 2 inhibited oat growth (Figure 3).
Sorghum-Sudangrass plus Japanese Millet Uniformity Trial
A total of 29 weed species were recorded in uniformity trial 2 (sorghum-sudangrass plus Japanese millet). Species occurring in more than 50% of the 32 plots were, in decreasing order of frequency, common chickweed, pigweed, hairy galinsoga, large crabgrass, volunteer oat, and Persian speedwell. Eight species were observed in only one plot, whereas common chickweed and pigweed were observed in all plots.
Weed Density and Biomass in Sorghum-Sudangrass plus Japanese Millet
The aboveground weed density in the sorghum-sudangrass plus Japanese millet uniformity trial had significant system effects (Figure 4; Table 3). The bio-extensive system had a significantly lower weed density than in the other three systems, which illustrates that legacy effects of crop management systems can persist through at least one crop. However, the entry point effect on weed density was not significant. Moreover, weed biomass did not differ significantly among systems and entry points (Table 3). Weed community composition based on either weed density or biomass did not vary either (Table 4).

Figure 4 Mean weed density (top), mean weed biomass (middle), and mean biomass of sorghum-sudangrass and Japanese millet (bottom) by cropping system and crop rotation entry point in uniformity trial 2. Bars with the same letters are not significantly different (Tukey-Kramer, P>0.05) among cropping systems (uppercase letters) and entry points (lowercase letters).
The weaker legacy effect in the sorghum-sudangrass plus Japanese millet uniformity trial relative to the oat uniformity trial has two possible causes. First, the germinable seedbank may have been sufficiently depleted by the oat trial to dampen differences among systems and entry points in the sorghum-sudangrass plus Japanese millet trial. Second, a large percentage of the seedbank may have been dormant in the latter trial. A substantial portion of the seeds of many spring-germinating species like common lambsquarters and giant foxtail (Setaria faberi Herrm.) enter secondary dormancy in the warm, summer conditions that prevailed after the sorghum-sudangrass and Japanese millet were planted (Dekker Reference Dekker2003; Karssen Reference Karssen1980). In addition, relatively few cool-season species like common chickweed, shepherd’s purse, and Persian speedwell germinate in midsummer. If a large portion of the seedbank was dormant during the sorghum-sudangrass plus Japanese millet trial, that would also dampen differences between systems and entry points. Overall, the information generated from the uniformity trials and the seedbank emergence bioassay shows that the assessment method and timing (early spring in uniformity trial 1 vs. midsummer in uniformity trial 2) can affect the results and that a more complete understanding of the effects of cropping systems on weed community structure can be obtained by using multiple methods.
Sorghum-Sudangrass plus Japanese Millet Biomass
Mean biomass of sorghum-sudangrass plus Japanese millet ranged from 525 g m−2 in the intensive system in entry point 2 to 699 g m−2 in the ridge-tillage system in entry point 1. No significant differences in crop biomass were observed between the systems or the entry points (Table 3; Figure 4).
Correlation between Soil Weed Seedbank and the Emerged Weed Community
We used Mantel tests to evaluate the correlation between weed communities measured in the seedbank and uniformity trials (Figure 5). As expected, the weed density and weed biomass in each uniformity trial were correlated with one another, since the more weeds that emerge the more weed biomass will be produced with consistent species composition. Although the legacy effect apparently weakened after the oat uniformity trial, deep tillage (>10 cm) in preparation for planting a subsequent summer crop could bring dormant seeds near the soil surface where conditions favor germination and emergence. System and entry point differences might then again be expressed. The extent to which legacy effects persist for several years depends on the initial abundance and survival rates of the various species in the seedbank. Schweizer and Zimdahl (Reference Schweizer and Zimdahl1984) showed that after 3 yr of seedbank depletion, populations of redroot pigweed and common lambsquarters rebounded quickly when control was relaxed, whereas kochia [Kochia scoparia (L.) Schrad.] and wild buckwheat (Polygonum convolvulus L.) populations did not. These responses demonstrate both the legacy of the pre-experiment management that allowed build-up of redroot pigweed and common lambsquarters populations but also the legacy of the 3 yr of strict management that drove kochia and wild buckwheat populations to densities below observable levels for the next 3 yr. Légere et al. (Reference Légere, Stevenson and Benoit2011) reported that 12 yr after the termination of a field experiment, a residual weed management effect on total seed density was observed in a no-tillage system but not in other tillage systems. The no-tillage system in monoculture also had more pronounced weed management legacy effects than the no-tillage system with a crop rotation. These observations illustrate the potential longevity of weed management legacy effects.

Figure 5 Results from Mantel tests showing correlations between weed communities occurring in the soil weed seedbank, in oat (uniformity trial 1), and in sorghum-sudangrass plus Japanese millet (uniformity trial 2). Dashed lines and “n.s.” indicate correlation is not significant, whereas solid lines, bold text, and asterisks indicate correlation is significant (*, P≤0.05; ***, P≤0.001).
We report results from a soil weed seedbank analysis and series of uniformity trials after 10 yr of organic vegetable production and show that weed abundance and community composition differed among the four cropping systems tested. We found that the ridge-tillage system resulted in a weed community that differed from the other cropping systems. Although weed communities are known to shift from being dominated by annual species to being dominated by perennial species in response to reductions in tillage (Buhler et al. Reference Buhler, Stoltenberg, Becker and Gunsolus1994), none of the indicator species associated with the ridge-tillage system were perennial. Two species were associated with the intensive system, indicating that nutrient inputs might have acted as a community filter (Ryan et al. Reference Ryan, Smith, Mirsky, Mortensen and Seidel2010). The uniformity trials also provided insights into the persistence of legacy effects from management practices. Specifically, the uniformity trials showed that rapid weed seedbank depletion from recruitment without seed additions, reduced germination from midsummer planting, and competitive crops reduced the expression of legacy effects in uniformity trial 2.
Results from this research can contribute to the refinement of management practices in organic vegetable production and help farmers reduce weed populations and associated production challenges. We found that tilled fallows and preventing weed seed production, which were used in the bio-extensive system, effectively reduced weed abundance. Using data from 2005 to 2009, Chan et al. (Reference Chan, Caldwell, Rickard and Mohler2011) reported that net returns were 3.2, 2.2, and 2.0 times greater in the intensive, intermediate, and ridge-tillage systems compared with the bio-extensive system, respectively. Despite this apparent trade-off between weed suppression and profitability, many small-scale diversified vegetable farmers in the Northeast region are labor rather than land limited (Jabbour et al. Reference Jabbour, Zwickle, Gallandt, McPhee and Wilson2014; Lockeretz Reference Lockeretz1997). Thus taking a portion of land out of production to accommodate tilled fallows and other weed-suppressive practices (e.g., mowed perennials) is feasible. In other regions with greater competition for land and higher land prices, such as the Central Valley in California, taking land out of production to accommodate weed-suppressive practices might be more difficult to justify.
Acknowledgments
Multiple people contributed to this work, including Caroline Marschner, Steve McKay, Rick Randolph, Sarah Zipfel, Nicole Scott, Yerae Seo, Heather Scott, Marissa Weiss, Louis Widom, Chris Pelzer, Alex Eacker, Lauren Gabuzzi, and Jason Chari. This work was supported by grants from the U.S. Department of Agriculture (USDA) Organic Research and Extension Initiative (Project numbers 2004-51300-02230 and 2009-51300-05586), a grant from the New York Farm Viability Institute (Project number ARP 08 007), and a grant from the USDA Organic Transitions Program (2014-03385). Funds were also provided by the Cornell Agricultural Experiment Station (Multi-State Hatch Projects NE-1000 [Project number 125450], NE-1026 [Project number 3120006036 NYC-145843], and NE-1047 [Project number 231327 NYC-125800]), and French ANR COSAC (ANR-14-CE18-0007), the Burgundy Region in France (FABER project Couv’Herbi).