


Introduction
The lean carcasses demanded by retailers can only be achieved by supplying sufficient quantities of essential amino acids to pigs possessing a high genetic potential for muscle growth. In organic agriculture, the supply of high-quality protein still proves to be difficult (Zollitsch, Reference Zollitsch2007). Among other factors, this contributes to the fact that even those European countries with the largest organic pig production only reached market shares of 1.6% (Austria) and 1.0% (Switzerland) in 2011 (Früh et al., Reference Früh, Bochicchio, Edwards, Hegelund, Leeb, Sundrum, Werne, Wiberg and Prunier2014). In conventional pork production, the dominant high-quality protein source is soybean meal, supplemented by synthetic amino acids. Both sources for amino acids are not allowed in European organic agriculture (The Council of the European Union, 2007). In organic pig farming, grain legumes, particularly faba beans (Vicia faba L.) and peas (Pisum sativum L.) are the main protein sources, apart from scarce and expensive soybean cake and dried milk products. However, in the last years, the production of these grain legumes suffered from some drawbacks and especially peas showed high yield variability. They are sensitive to drought and require long rotation breaks of up to 10 years (Liebman and Dyck, Reference Liebman and Dyck1993; Hauggaard-Nielsen and Jensen, Reference Hauggaard-Nielsen and Jensen2005). Consequently, alternative and locally produced protein sources are of great interest to organic pig producers in Europe. The currently under-utilized grain legume grass pea (Lathyrus sativus L.) was identified as one such possible feed resource for pigs.
Grass pea is a grain legume that grows well both in drought and under waterlogged conditions and produces seeds with a high crude protein content of 250–270 g kg−1 dry matter even on nutrient-poor sites (Yan et al., Reference Yan, Spencer, Li, Liang, Wang, Wang and Li2006). It resembles peas in amino acid profile and has a similarly low content of the sulfur-containing amino acids methionine and cysteine [lys:(met + cys):thr:trp of peas: 100:33:53:13; Evonik-Degussa GmbH (2006)]. Grass pea seeds are a widespread staple crop in the Mediterranean area, in India and in parts of North Africa (Padmanaban, Reference Padmanaban and Liener1980). Due to its resilience, grass pea seeds are especially important in times of severe food shortages. However, apart from trypsin inhibitors, grass pea seeds contain a neurotoxic substance, the non-proteinogenic amino acid β-N-oxalyl-L-α,β-diaminopropionic acid (ODAP), which in humans causes irreversible nerve damage after excessive and/or prolonged consumption, known as neurolathyrism (Padmanaban, Reference Padmanaban and Liener1980; Hanbury et al., Reference Hanbury, White, Mullan and Siddique2000). In an experiment with fattening pigs, pathological enlargements of liver and kidney were observed when raw grass pea seeds constituted 20% or more of the diet (Winiarska-Mieczan and Kwiecien, Reference Winiarska-Mieczan and Kwiecien2010). Because ODAP is water-soluble and susceptible to heat, both hydrothermal and thermal treatment reduce the ODAP content considerably (Padmajaprasad et al., Reference Padmajaprasad, Kaladhar and Bhat1997; Hanbury et al., Reference Hanbury, White, Mullan and Siddique2000).
In Austria, the cultivation area of grass peas has been increasing, but still remains on a relatively low level (1646 ha in 2011, Grüner Bericht, 2012), and small percentages of the seeds are used in commercial compound feed for ruminants and pigs (P. Binggl, pers. comm. April 23, 2012).
Although several feeding trials have been published on the use of grass pea seeds as protein source for fattening pigs (Castell et al., Reference Castell, Cliplef, Briggs, Campbell and Bruni1994; Trombetta et al., Reference Trombetta, Matti, Pasquini and Falaschini2006; Winiarska-Mieczan and Kwiecien, Reference Winiarska-Mieczan and Kwiecien2010), information about its use for piglets is scarce and only one experiment is known to the authors (Castell et al., Reference Castell, Cliplef, Briggs, Campbell and Bruni1994). Therefore, we saw a need for additional information on the suitability of grass pea seeds as feed for piglets, which are expected to be especially sensitive against anti-nutritive compounds of grain legumes (Dwivedi, Reference Dwivedi1989). In two feeding trials, both raw and hydrothermally treated grass pea seeds were included in diets for weaned piglets at inclusion rates of 10–30% and were compared with control diets. The aims of the experiments were to examine whether the rate of inclusion of grass pea seeds influences piglets’ performance, and whether an effect of hydrothermal treatment of the grass pea seeds can be found on performance traits. Selected blood metabolites of piglets were analyzed to detect potential metabolic effects of dietary treatment.
Animals, materials and methods
The feeding experiments with grass pea seeds were carried out at the Institute of Biological Agriculture and Biodiversity of Farm Animals (part of the Austrian Research and Education Center) in Wels, Austria. The first experiment took place between December 2009 and June 2010 and the second between May and November 2012. Both experiments followed the rules of EU Directive 2010/63/EU for animal experiments (The European Parliament and the Council of the European Union, 2010) and the Austrian Act on Animal Experiments (Austrian Republic, 1999). No invasive procedures were performed on the animals except the collection of blood samples, which was performed by a veterinarian.
Experimental design and animals
In both feeding trials, four adjacent pens of identical dimensions and equipment were available. Because feed allowance could not be documented for the individual piglet, but only for the group of piglets housed in one pen, a total of four experimental diets were fed simultaneously. This was repeated four times (called ‘replicate’ hereafter). The allocation of the experimental diets to the pens was changed in each replicate, so that each diet was fed once in each pen. This way, pen and replicate could be considered as blocking factors, and the experimental design as a complete 4 × 4 latin square design. Diets were fed during the rearing phase which started immediately after weaning and lasted for 33 and 29 days in experiments 1 and 2, respectively.
Piglets [crosses of (Pietrain × Duroc) × (Landrace × Large White)] weighing between 10 and 16 kg at weaning and without clinical signs of illness were selected for the feeding trial. A total of 152 piglets from 19 litters (weaning age 41 ± 1.7 days) were included in experiment 1. In experiment 2, a total of 144 piglets from 25 litters were used, and piglets were weaned at 47 ± 5.5 days. Immediately after weaning, piglets were divided into groups with the aim of achieving four equally large groups with a comparable mean body weight and mean blood haptoglobin level. In addition, we tried to combine piglets from a similar number of litters (sows) in each group and to have the same sex ratio whenever possible. The number of piglets per group was 12, 11, 8 and 7 in the four replicates of experiment 1 and 8, 10, 10 and 9 in the four replicates of experiment 2, respectively. In both experiments, the groups of piglets were housed in straw-bedded pens, equipped with a creep area, drinkers, feeder and an outdoor area. The exact measurements of the pens were slightly different: in experiment 1, pens had the size of 5.5 × 1.8 m, plus 3.4 × 1.8 m outdoor area, while in experiment 2, pen size was 5 × 1.7 m plus an outdoor area of 3 × 1.7 m.
Feeding regimen
Treatment of grass pea seeds and production of the diets were done at the feed mill Vitakorn Biofuttermittel GesmbH. Hydrothermal treatment consisted of exposing the grass pea seeds to steam and heat (98°C) for 20 min. In both experiments, the main protein-rich components of the control diet were peas and soybean cake; these were (totally or partially) substituted for by grass pea seeds in the experimental diets and barley was used to balance the dietary energy content. The nutrient contents of grass pea seeds are summarized in Table 1. For the composition and nutrient contents of the experimental diets, see Table 2.
Table 1. Nutrient contents of grass pea seeds harvested in 2009 (experiment 1) and 2011 (experiment 2), g kg−1 (as fed) unless stated otherwise.

NDF, neutral detergent fiber; ADF, acid detergent fiber; ME, metabolizable energy; n.a., not analyzed.
1 Metabolizable energy content was calculated according to GfE (1988).
Table 2. Composition and nutrient content of the diets in experiments 1 and 2, g kg−1 (as fed) unless stated otherwise. C 1, C 2 = control diets; R 10.1, R 20.1, R 20.2 = 10 and 20% raw grass pea seeds, respectively, T 20.1, T 20.2, T 30.2 = 20 and 30% hydrothermally treated grass pea seeds, respectively. For the nutrient contents of the diets, the table shows least-square estimates and P-values for the effect of diet.

RSD, residual standard deviation; NDF, neutral detergent fiber; ADF, acid detergent fiber; ME, metabolizable energy; –, not applicable; n.a., not analyzed.
1 Metabolizable energy content was calculated according to GfE (1988); means within a row with different superscripts differ (P < 0.05). P-values indicating significant differences are shown in bold font.
Experiment 1. The grass pea seeds for experiment 1 were bought from an organic farmer in Lower Austria, a province of Austria. Before weaning, piglets were offered a commercial organic piglet starter feed consisting of wheat, barley, triticale, maize, oats, peas, soybean cake, rapeseed cake, toasted full-fat soybeans, potato protein (from a conventional source), wheat bran and spelts bran. The calculated nutrient and energy content of the all-mash starter feed was 199 g crude protein, 46 g ether extracts, 55 g crude fiber, 61 g ash and 11 g total lysine kg−1, and the estimated energy content was 13.1 MJ metabolizable energy (ME) kg−1 (as fed basis; ME calculated according to GfE (2008)). For 1 week prior to weaning, piglets were offered a 1:1 mixture of the all-mash starter feed and the pelleted control diet (C 1) to get used to pelleted feed. In the feeding trial, the control diet was compared with three diets containing grass pea seeds: Two diets with raw grass pea seeds, 10% (R 10.1) and 20% (R 20.1), respectively, and one diet with 20% hydrothermally treated grass pea seeds (T 20.1, as fed basis). Diets were formulated to be iso-energetic and contained the same amounts of lysine and methionine. In the experimental diets, peas were partially or fully replaced by grass pea seeds. The mineral mix included in all the diets consisted of calcium carbonate, monocalcium phosphate and sodium chloride and contained 22.6% calcium, 9.2% phosphor and 4.7% sodium. Piglets had ad libitum access to the pelleted diets by mean of automatic dry feeders.
Experiment 2. In experiment 2, grass pea seeds were purchased from an organic farmer in the Austrian province of Burgenland. Before weaning, the piglets received a commercial organic piglet starter feed consisting of wheat, barley, oats, peas, soybean cake, skimmed milk powder and pumpkin seed cake. The calculated nutrient and energy content of the all-mash starter feed was 207 g crude protein, 35 g ether extracts, 42 g crude fiber, 61 g ash and 9.6 g total lysine kg−1, and the estimated energy content was 13.6 MJ ME kg−1 (as fed basis; ME calculated according to GfE (2008)). In the feeding trial, the control diet (C 2) was compared with three diets containing grass pea seeds: one diet with 20% raw grass pea seeds (R 20.2) and two diets with 20 and 30% hydrothermally treated grass pea seeds, respectively (as fed basis). The diets were formulated to be iso-energetic and had the same content of lysine. In diets R 20.2 and T 20.2, peas were completely replaced by grass pea seeds and the proportion of soybean cake was reduced. In diet T 30.2, the percentage of soybean cake was further reduced. The composition of the mineral mix was as described above for experiment 1. The granulated diets were distributed using an automated feeding system, programmed to supply feed at five times per day, in amounts slightly increasing every day. Feed allowance was based on recommendations by LfL (Bavarian State Research Institute for Agriculture, 2011), but to prevent diarrhea, feed allowance was restricted and adjusted daily to insure complete intake of feed until the next meal. Feed supply therefore differed between the groups of piglets depending on their individual feed intake.
Data collection and analytical procedures
Experiment 1. Feed intake was recorded for each pen by weighing feed input into the automatic feeder and weighing back feed refusals weekly. Bodyweight of piglets was documented on days 1, 4, 12, 19, 26 and 33. Feed efficiency was defined as body weight gain (weekly group averages) divided by feed intake. Blood samples of piglets were taken on days 1 and 33 by puncturing the vena jugularis externa, using vacutainer tubes. To conserve the samples until analysis at the institute's laboratory, blood was centrifuged at 3500 rpm for 10 min. Within 1 h after taking the samples, serum was stored at −21°C. Commercial kits were used for photometric determination of the concentrations of serum cholesterol, albumin, urea and total protein (cobas® c 111 system, Roche Diagnostics GmbH, Mannheim, Germany; 2009, 2011, 2012 and 2012, respectively). A commercial kit (BioRépair GmbH, Sinsheim, Germany) was used for photometric determination of serum haptoglobin concentration.
Prior to the feeding trial, one sample of grass pea seeds was taken for analysis of nutrients and amino acids. A total of four samples were taken of each diet throughout the experiment, comprising one feed sample per diet per replicate. Analysis of nutrient contents was done at a commercial laboratory according to VDLUFA (2007), and the method numbers are as follows: Method 3.1 was used to determine the dry matter content of feed by oven-drying at 105 °C. Methods 8.1, 5.1.1, 6.1.1, 7.1.1 and 7.2.1 were used to measure ash, ether extracts, crude fiber, sugar and starch contents, respectively. The content of crude protein was analyzed by Dumas combustion, following method 4.1.2. Contents of ME were calculated according to GfE (2008). Piglets were examined for lameness once a week and their gait was scored according to the following scheme: 0 = no impairment of posture and gait; 1 = weakness in the hind legs, visible as unsteady gait with front legs wider than usual and head bowed low to keep the balance; 2 = lameness of the hind legs, piglet can either only stand on the front legs or not stand up at all. Due to a high prevalence of symptoms of post-weaning diarrhea, a metaphylactic antibiotic treatment (5 mg Colistinsulfat and 8 mg Tylosin-Tartrat kg−1 body weight d−1) was administered to all piglets for the first 9 days of the experiment to avoid distortions from impaired gut health of individual piglets.
Experiment 2. Throughout the experiment, feed allowance was automatically recorded for each pen. Feed refusals were weighed and subtracted from feed supply, but were rarely observed due to the restricted feeding regimen. In the absence of feed refusals, feed supply was interpreted as feed intake. Piglets were weighed once a week, on days 1, 8, 15, 22 and 29. Feed efficiency was calculated, grass pea, feed and blood samples were collected and gait was scored as described above for experiment 1. Neutral detergent fiber (6.5.1) was determined using a heat stable amylase. Both neutral detergent fiber (NDF) and acid detergent fiber (ADF; 6.5.2) are expressed including residual ash. In replicates 1 and 3, an additional feed sample was taken per diet for analysis of the amino acid profile at the Department of Chemistry of BOKU-University of Natural Resources and Life Sciences Vienna. Amino acid concentrations were determined with a high-performance liquid chromatography system of Hewlett-Packard (1050 Series) and a Shimadzu RF 535 variable wavelength fluorescence detector, using samples that had been hydrolyzed in 6 M HCl for 20 h. Tryptophan was stabilized with alkaline Ba(OH)2, and to avoid methionine and cysteine loss, samples were oxidized (ALVA, 1983; Altmann, Reference Altmann1992; The European Parliament and the Council of the European Union, 2000). Whenever symptoms of diarrhea were observed, all piglets were given tea of Cortex quercus, dry peat and an electrolyte solution. When symptoms of diarrhea persisted, individual piglets were treated with an antibiotic (Baytril).
Statistical analysis
Experiments 1 and 2 were analyzed separately, using the program package SAS (SAS 2002). For multiple comparison of means, the Tukey test was performed, and when the P-value was <0.05, the statistical difference was interpreted as significant. The nutrient contents of the experimental diets were analyzed using proc GLM, with a model including the fixed effect of diet only. Proc GLM was also used for the animal-related parameters feed intake (g d−1 piglet−1) and feed efficiency (kg weight gain kg−1 feed intake), which were measured for groups of piglets. Those parameters which were measured for individual piglets (body weight, daily weight gain and blood metabolites) were analyzed using proc MIXED, which enables the inclusion of the random effect of individual piglet. The models were as follows:
Feed intake and feed efficiency
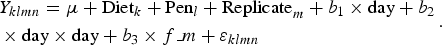 $$\eqalign{&Y_{klmn} = \mu + {\rm Diet}_k + {\rm Pen}_l + {\rm Replicate}_m + {\rm} b_1 \times {\rm day} + {\rm} b_2 \cr & \times {\rm day} \times {\rm day} + {\rm} b_3 \times f\_m + \varepsilon _{klmn}}. $$
$$\eqalign{&Y_{klmn} = \mu + {\rm Diet}_k + {\rm Pen}_l + {\rm Replicate}_m + {\rm} b_1 \times {\rm day} + {\rm} b_2 \cr & \times {\rm day} \times {\rm day} + {\rm} b_3 \times f\_m + \varepsilon _{klmn}}. $$
In experiment 2, feed efficiency was not analyzed for the first week after weaning because a considerable number of piglets lost weight during this period. Analysis of feed efficiency over the whole rearing phase was done separately and included the data from the first week.
Body weight and daily weight gain
 $$\eqalign{&Y_{klmnopq} = \mu + {\rm Diet}_k + {\rm Pen}_l + {\rm Replicate}_m \cr & + {\rm Sow}_o \left( {{\rm Replicate}_m} \right)+ {\rm Day}_n + {\rm} b_4 \, \times \,bw\_{\rm weaning} \cr &+ {\rm Piglet}_p \left( {{\rm Diet}_k} \right) + {\rm Diet}_k \, \times \,{\rm Day}_n + \varepsilon _{klmnop}. }$$
$$\eqalign{&Y_{klmnopq} = \mu + {\rm Diet}_k + {\rm Pen}_l + {\rm Replicate}_m \cr & + {\rm Sow}_o \left( {{\rm Replicate}_m} \right)+ {\rm Day}_n + {\rm} b_4 \, \times \,bw\_{\rm weaning} \cr &+ {\rm Piglet}_p \left( {{\rm Diet}_k} \right) + {\rm Diet}_k \, \times \,{\rm Day}_n + \varepsilon _{klmnop}. }$$
Because of a lack of significance, the initially included effect of sex was removed from the model. The following eight covariance structures were identified as being potentially suitable: unstructured, autoregressive, heterogenous autoregressive, autoregressive moving average, compound symmetry, heterogenous compound symmetry, Toeplitz and heterogenous Toeplitz. Toeplitz and compound symmetry were chosen for body weight and daily weight gain, respectively, because their Bayesian information criterion indicated the best fit of the model to the data (Wang and Goonewardene, Reference Wang and Goonewardene2004; Littell et al., Reference Littell, Milliken, Stroup, Wolfinger and Schabenberger2006). The interaction diet × day was only significant in experiment 2; therefore it was dropped from the model for analysis of data from experiment 1.
Blood metabolites
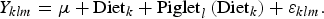 $$Y_{klm} = \mu + {\rm Diet}_k + {\rm Piglet}_l \left( {{\rm Diet}_k} \right) + {\rm \varepsilon} _{klm}. $$
$$Y_{klm} = \mu + {\rm Diet}_k + {\rm Piglet}_l \left( {{\rm Diet}_k} \right) + {\rm \varepsilon} _{klm}. $$
Statistical analysis of blood metabolites was done separately for the first and last day of the experimental period, using the covariance structure ‘variance components’.
Legend:
Y = variable studied; μ = overall mean; Diet = fixed effect of diet (k = C1, C2, R10.1, R20.1, R20.2, T20.1, T20.2, T30.2); Pen = fixed effect of pen (l = 1, 2, 3, 4); Replicate = fixed effect of replicate (m = 1, 2, 3, 4); Sow(Replicate) = fixed effect of sow within replicate (o = ear tag number of sow); Day = fixed effect of day in the feeding trial (experiment 1: n = 1, 12, 19, 26, 33 and experiment 2: n = 1, 8, 15, 22, 29); b 1, b 2 = regression coefficients of day and day × day; day, day × day = continuous linear and quadratic effect of day in the feeding trial; b 3 = regression coefficient of f_m; f_m = continuous effect of ratio between female and male piglets in the pen; b 4 = regression coefficient of bw_weaning; bw_weaning = continuous effect of body weight at weaning; Piglet l (Diet k ) = random effect of piglet within diet (l = ear tag number of piglet); Diet k × Day n = interaction between diet and day; ε = random error.
Results and discussion
Feed intake
In experiment 1, including 20% treated grass pea seeds in the diet resulted in a significantly higher intake of feed, energy and crude protein as compared with the treatment with 20% raw grass pea seeds (Table 3). This seems to allow the conclusion that hydrothermal treatment of grass pea seeds improves feed intake, which is in accordance with results published by Castell et al. (Reference Castell, Cliplef, Briggs, Campbell and Bruni1994) and Rotter et al. (Reference Rotter, Marquardt and Campbell1991), who observed a reduction in feed intake of pigs and chicken when increasing the dietary percentage of raw grass pea seeds. Feed intake of piglets receiving the control diet and diet R 10.1 was numerically lower than for treatment T 20.1 and higher than for R 20.1, but the differences were not statistically significant.
Table 3. Performance of piglets fed organic diets with or without grass pea seeds in experiment 1. C 1 = control diet; R 10.1 and R 20.1 = 10 and 20% raw grass pea seeds, respectively; T 20.1 = 20% hydrothermally treated grass pea seeds. The table shows least-square estimates and P-values of the effect of diet for the whole experimental period. In addition, least-square estimates for individual time periods are given for further illustration.

RSD, residual standard deviation; CP, crude protein; means within a row with different superscripts differ (P < 0.05). P-values indicating significant differences are shown in bold font.
Diets R 20.1 and T 20.1 in experiment 1 were not identical to diets R 20.2 and T 20.2 in experiment 2, but similar enough to give reason to expect a similar response in performance. Nevertheless, no significant difference in feed intake could be found in experiment 2 (Table 4), although feed intake was numerically smallest when diet R 20.2 was fed. A likely explanation for the observed differences might be that the grass pea seeds used in the two experiments differed with regard to their content of ODAP and other anti-nutritive components such as trypsin inhibitors, lectins and tannins (Sharma et al., Reference Sharma, Kalia and Malhotra2003; Ramachandran et al., Reference Ramachandran, Bairagi and Ray2005). But because the anti-nutritive factors in the grass pea seeds were not analyzed, this cannot be explored further.
Table 4. Performance of piglets fed organic diets with or without grass pea seeds in experiment 2. C 2 = control diet; R 20.2 = 20% raw grass pea seeds; T 20.2 and T 30.2 = 20 and 30% hydrothermally treated grass pea seeds, respectively. The table shows least-square estimates and P-values of the effect of diet for the whole experimental period, unless stated otherwise. In addition, least-square estimates for individual time periods are given for further illustration.

RSD, residual standard deviation; fat intake, intake of ether extracts; CP, crude protein; means within a row with different superscripts differ (P < 0.05). P-values indicating significant differences are shown in bold font.
1 For body weight and daily weight gain, least-square estimates for interaction diet × day are given, except for overall daily weight gain, for which the least-square estimate for diet is shown.
Because the crude protein and lysine content of grass pea seeds (lysine content 17.9–19.5 g kg−1) is only slightly higher than that of peas (lysine content 15.2 g kg−1, Evonik-Degussa GmbH, 2006), only a slight effect of including grass pea seeds in the diet on the crude protein content of the diets was found: in experiment 1, there was no effect at all, and in experiment 2 only a tendency (P = 0.052) towards a lower crude protein content in diet T 30.2 compared with the control was observed. The above-mentioned significant differences in feed intake found in experiment 1 resulted in equally significant differences in crude protein intake. In experiment 2, the tendencies in dietary crude protein content and numerical differences in feed intake led to a significantly lower intake of crude protein in piglets fed diet R 20.2 as compared with the control diet. As a consequence, lysine intake in experiment 2 was lowest when diet R 20.2 was fed. The relationship of the amino acids lys:(met+cys):thr:trp of the grass pea seeds fed in the experiments was 100:38:53:13, which is quite similar to that of peas (100:33:53:13, Evonik-Degussa GmbH, 2006) and is therefore as deficient in methionine and cysteine, when compared with the ideal relationship for piglets of 5–20 kg bodyweight (100:60:65:18, Blair, Reference Blair2007). Consequently, methionine intake was significantly lower in all diets of experiment 2 which included grass pea seeds as compared with the control diet.
Growth performance
In experiment 1, no significant differences in body weight were found between the diets, despite differing feed intake (Table 3). At weaning, piglets weighed on average 12.3 kg, and at the end of the experimental period they had reached an average weight of 25.2 kg.
In experiment 2, the statistical analysis of body weight and daily weight gain revealed a significant interaction between diet and day, indicating that the effect of including grass pea seeds in the diet changed over time: 1 week after weaning, no difference between diets could be observed, but 2 weeks after weaning, the body weight of piglets fed diet R 20.2 was already significantly lower than the control, but not statistically different from T 20.2 and T30.2 (Table 4). At weaning, piglets weighed on average 12.9 kg. At the end of the 29 days rearing phase, piglets fed the control diet on average weighed 24.3 kg, while feeding diet R 20.2 resulted in an average body weight of only 21.6 kg. During the experimental period, the inclusion of treated grass pea seeds did not affect body weight, but on day 29 piglets receiving diet T 30.2 weighed significantly less than piglets fed diet C 2.
In analogy to body weight, daily weight gain was not affected by diet in experiment 1. In experiment 2, daily weight gain did not vary during the first 8 days after weaning. But from day 9 onwards, daily weight gain was significantly lowest when diet R 20.2 was fed. Including treated grass pea seeds in the diet resulted in slightly lower daily weight gains than in the control, but the difference was only significant for days 16–22. On average, piglets gained 422 g d−1 in experiment 1 and 355 g d−1 in experiment 2, with the growth depression for all piglets in the first week after weaning in experiment 2 being the main reason for this difference. The general level of body weight gain was lower than that reported by Castell et al. (Reference Castell, Cliplef, Briggs, Campbell and Bruni1994), who fed diets containing 10–40% raw grass pea seeds to conventionally reared piglets and found a progressive reduction of feed intake and daily weight gain with increasing percentage of grass pea seeds. For fattening pigs, Winiarska-Mieczan and Kwiecien (Reference Winiarska-Mieczan and Kwiecien2010) recommended that raw grass pea seeds should not make up more than half of the protein-rich feed components in a diet, which in experiments 1 and 2 would have been 17.5 and 19.5%, respectively. Confirmation for this recommendation can be found in reports by Martelli et al. (Reference Martelli, Rizzi, Boccuzzi, Paganelli, Sardi, Alberto and Costa2009), who conducted a digestibility trial with fattening pigs fed diets containing 19% raw grass pea seeds and did not record any health problems in pigs. Knowing that young piglets are more sensitive to ODAP than older pigs (Dwivedi, Reference Dwivedi1989), the growth depression observed when 20% raw grass pea seeds were included in the diet (diets R 20.1 and R 20.2) agrees well with the recommendation of Winiarska-Mieczan and Kwiecien (Reference Winiarska-Mieczan and Kwiecien2010). Trombetta et al. (Reference Trombetta, Matti, Pasquini and Falaschini2006) found a comparable growth performance and digestibility of diets when partially substituting soybean meal in diets for fattening pigs by 10 and 20% extruded grass pea seeds, respectively. This is in accordance with the results from both experiments 1 and 2, in which piglets’ growth was not affected by replacing and partially substituting peas and soybean cake by treated grass pea seeds. However, despite feed intake being significantly higher when only 10% raw grass pea seeds were included, daily weight gain did not differ between piglets fed diets R 10.1 and R 20.1. Neither ODAP content nor trypsin inhibitors or other anti-nutritive substances in grass pea seeds were analyzed in the two experiments, therefore it is not possible to ascribe the negative effect of raw grass pea seeds on piglets’ feed intake (experiment 1) and growth (experiment 2) directly to ODAP. Hydrothermal treatment reduces trypsin inhibitors and other anti-nutritive compounds as well as ODAP, consequently the negative effects of raw grass pea seeds are likely the result of the sum of negative effects of the present anti-nutritive compounds. Castell et al. (Reference Castell, Cliplef, Briggs, Campbell and Bruni1994) explained the performance depression they observed in piglets fed raw grass pea cultivars with low ODAP contents to the presence of a trypsin inhibitor, which prevailed in seeds despite low ODAP levels. The same authors also fed fattening pigs with diets containing up to 30% raw grass pea seeds and failed to find ODAP residues when analyzing loin samples (Castell et al., Reference Castell, Cliplef, Briggs, Campbell and Bruni1994). Symptoms of lathyrism in humans do not appear immediately after intake, but rather develop with time after a prolonged consumption (Padmanaban, Reference Padmanaban and Liener1980). Therefore, the potential risk of ODAP residues in meat should be taken seriously and this issue requires further research. The prevention of potential health risks for pork consumers is another reason for issuing a strong suggestion to feed only heat treated grass pea seeds to fattening animals.
In experiment 1, feed efficiency was not affected by dietary treatment. Non-significant numerical differences between diets resulted from feed intake being significantly lower when diet R 20.1 was fed as compared with diet T 20.1, while body weight gain did not differ. In experiment 2, the significant negative effect of including raw grass pea seeds in diet R 20.2 on body weight gain also had an impact on feed efficiency, which was significantly lower when diet R 20.2 was fed as compared with all other diets.
Blood metabolites
Neither in the first nor in the second experiment, significant differences between dietary treatments could be found for blood metabolites at weaning, indicating that piglets were evenly distributed to treatments. In experiment 1, the main effect of diet on blood haptoglobin level was significant at the end of the experimental period, but pairwise comparison of means only showed a strong tendency (P = 0.052) towards a difference between diets R 20.1 and T 20.1 (Table 3). In experiment 2, statistically significant differences at the end of the experimental period could be observed for albumin, cholesterol and urea (Table 4).
Blood haptoglobin is an acute phase protein and as such a component of the body's response to inflammations or infections, hence an elevated blood haptoglobin level may be seen as a non-specific sign of sub-clinical health problems (Petersen et al., Reference Petersen, Nielsen and Heegaard2004). The general blood haptoglobin level observed in experiment 1 was higher than in experiment 2, which can be interpreted as a sign that efforts to improve the general health status of the piglets on the experimental station were successful. Nevertheless, all values in both experiments were within the reference range for healthy piglets (Petersen et al., Reference Petersen, Nielsen and Heegaard2004; Piñeiro et al., Reference Piñeiro, Piñeiro, Morales, Andrés, Lorenzo, del Pozo, Alava and Lampreave2009). The observation that blood haptoglobin levels increased with age is in accordance with those authors.
In both experiments, the level of the major plasma protein albumin was in accordance with previous reports (Fürll and Röhl, Reference Fürll, Röhl, Schubert, Flachowsky, Jahreis and Bitsch2003). At the end of experiment 2, blood albumin levels were lowest when diet R 20.2 was fed, and highest when diets T 20.2 and T 30.2 were fed, but despite being statistically significant, these differences were numerically small and do not seem to be relevant for piglets’ health.
Blood cholesterol level is largely determined by dietary fat and cholesterol intake, also high intakes of crude protein have been found to downregulate blood cholesterol (Theodorou et al., Reference Theodorou, Papadomichelakis, Tsiplakou, Lampidonis, Chadio, Zervas and Politis2015). The general level of crude protein intake was higher in experiment 1 than in experiment 2, corresponding to lower blood cholesterol levels in experiment 1 as compared with experiment 2. At the end of experiment 2, blood cholesterol levels were highest in piglets fed the control diet, paralleled by intake of ether extracts being significantly highest when the control diet was fed. Blood cholesterol levels were significantly lower when diet T 20.2 was fed as compared with the control, and significantly lowest when diet T 30.2 was fed, while intake of ether extracts did not differ between diets R 20.2, T 20.2 and T 30.2.
Blood urea is an indicator of protein metabolism, namely the amount of metabolized protein and how well the amino acid pattern of the diet matches the animal's demand. Increased blood urea levels are the result of surplus amino acids being degraded to urea before they are excreted. However, despite a significantly lower crude protein intake of piglets fed diet R 20.2, their blood urea levels were significantly higher as compared with those fed diets C 2 and T 30.2. Most likely, the reduced growth rate of piglets fed raw grass pea seeds resulted in a more positive protein balance and therefore higher amino acid breakdown and urea excretion. Nevertheless, all analyzed blood urea values were still within the reference range (Kraft and Dürr, Reference Kraft and Dürr2005). To summarize, the levels of all blood metabolites analyzed throughout both experiments were within their respective reference ranges, and no signs of metabolic changes related to dietary treatment could be found.
Animal health
Throughout experiment 1, five piglets died of various reasons, one of them in the first, two in the third and two in the fourth replicate. Two of the lost piglets had been fed the control diet, and of the remaining three piglets each had been fed a different experimental diet. Because of the quite even distribution of the mortalities, no causal link between treatment and deaths was suspected. In experiment 2, one piglet was excluded during the second replicate due to persisting diarrhea, and three piglets were excluded during the third replicate. Two of these piglets had been fed diet R 20.2 and the other two had been fed diet T 20.2. After the initial metaphylactic antibiotic treatment in experiment 1, no further treatments against diarrhea were necessary. In experiment 2, antibiotic treatment was only necessary for individual piglets. Five piglets fed the control diet, four piglets fed diet R 20.2, six piglets fed diet T 20.2 and two piglets fed diet T 30.2 were treated, which corresponds to 14, 11, 16 and 5% of the piglets, respectively. A recent study on health and welfare of organic pigs in six European countries found a highly variable prevalence of diarrhea with values between 0 and 100%, but the median prevalence was 0% because groups were scored rather than piglets (Dippel et al., Reference Dippel, Leeb, Bochicchio, Bonde, Dietze, Gunnarsson, Lindgren, Sundrum, Wiberg, Winckler and Prunier2013). Weekly gait scoring in experiment 1 and at the end of each replicate in experiment 2 did not reveal any signs of lameness which might have indicated nerve disorders caused by ODAP.
Conclusions
Including hydrothermally treated grass pea seeds in proportions of 20–30% in diets for weaned piglets resulted in feed intake and daily weight gain similar to that for the control diets. Including 20% raw grass pea seeds led to a significantly lower feed intake in experiment 1 and significantly lower daily weight gain in experiment 2. Due to these negative effects of feeding raw grass pea seeds, hydrothermal treatment of grass pea seeds prior to feeding to piglets is strongly recommended.
Acknowledgements
Experiment 2 was part of the ERA-NET CORE Organic II Research Project ICOPP (improved contribution of local feed to support 100% organic feed supply to pigs and poultry, see www.organicresearchcentre.com/icopp). Funding was provided by the Austrian Federal Ministry of Agriculture, Forestry, Environment and Water Management and is gratefully acknowledged. Furthermore, the authors want to thank Franz Traudtner (Bio Austria) for his valuable inputs during planning and preparation of this study. We also acknowledge the valuable comments received from two anonymous reviewers on an earlier version of this paper.




