



INTRODUCTION
Shellmounds represent the first evidence of human occupation on the Brazilian coast during the Holocene (Gaspar Reference Gaspar1998). Often called sambaquis, the major characteristic of these archaeological sites is the intentional accumulation of mollusk shells. Mollusk flesh was used as a food source and the shells in the construction of the mounds, as tools or in funerary rituals (Gaspar Reference Gaspar1998). In some sites, certain shell species are found disposed in patterns, associated with graves, with evidence of painting or in the form of beads or scrapers (e.g. Kneip and Machado Reference Kneip and Machado1993; Tenório Reference Tenório2007; Klokler Reference Klokler2008). In some cases consumption and ritual use were interconnected, complicating the interpretation of some attributes that are commonly used to distinguish ritual fauna (Klokler Reference Klokler2008).
Although most of the coastal shellmounds consist of marine mollusk shells, terrestrial species are also found composing the archaeological context (Figure 1) in many of these sites alongside the Brazilian coast (Table 1). Moreover, land snails are present in non-coastal sites throughout the Brazilian territory (Prous Reference Prous1986; Mello and Coelho Reference Mello and Coelho1989; Moreira et al. Reference Moreira, Araujo, Confalonieri, Ferreira and Prous1991; Magalhães et al. Reference Magalhães, Curvelo and Mello2001; Brentano et al. Reference Brentano, Rosa and Schmitz2006; Rogge Reference Rogge2006; Rosa Reference Rosa2006; Teixeira Reference Teixeira2006; Gernet and Birckolz Reference Gernet and Birckolz2011; Agudo-Padrón Reference Agudo-Padrón2012; Bandeira Reference Bandeira2013; Beltramino Reference Beltramino2013; Hadler et al. Reference Hadler, Dias and Bauermann2013). Among the species commonly found are those from the genera Megalobulimus (Miller 1878) and Thaumastus (Martens 1860) (Figure 2), both endemic to South America and characteristic of high humidity environments (Colley Reference Colley2012; Fontenelle et al. Reference Fontenelle, Cavallari and Simone2014; Macario et al. Reference Macario, Alves, Carvalho, Oliveira, Ramsey, Chivall, Souza, Simone and Cavallari2016a). Therefore, their presence is also a climate indicator, since it is related to rainy seasons or periods of increased humidity (Abbott Reference Abbott1989; Rosa Reference Rosa2006). Both genera have detritivorous-herbivorous diet, feeding from the leaves of trees and shrubs. They have nocturnal habits, hiding below the leaf coverage in soil during the day and dormancy periods (Bequaert Reference Bequaert1948; Morretes Reference Morretes1952; Miranda and Fontenelle Reference Miranda and Fontenelle2015; Cavallari pers. communication).

Figure 1 Land snails’ abundance at the Usiminas shellmound. Photographs: Luciano Rapagña.

Figure 2 Samples from the Usiminas: (A) Megalobulimus (B) Thaumastus and Saquarema shellmounds: (C) Thaumastus. Photographs: Bruna Brandão.
Table 1 Occurrence of the studied terrestrial snails’ genera reported in Brazilian archaeological sites.
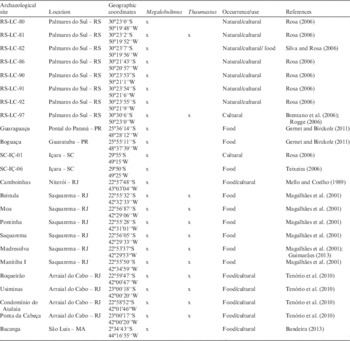
Occurring in both past and modern terrestrial environments, the presence of land snails in the archaeological record is not necessarily associated to the archaeological context (Rosa Reference Rosa2006). However, intentional fracture and other anthropic marks increase the probability of previous use as tools or in rituals (Magalhães and Mezzalira Reference Magalhães and Mezzalira1953; Leme Reference Leme1975; Solá and Jokl Reference Solá and Jokl1978; Mello and Coelho Reference Mello and Coelho1989; Simone and Mezzalira Reference Simone and Mezzalira1994; Kneip Reference Kneip2001; Magalhães et al. Reference Magalhães, Curvelo and Mello2001; Brentano et al. Reference Brentano, Rosa and Schmitz2006; Rogge Reference Rogge2006; Teixeira Reference Teixeira2006; Gernet and Birckolz Reference Gernet and Birckolz2011; Fontenelle et al. Reference Fontenelle, Cavallari and Simone2014). The depth where the shells are retrieved on the site, the abundance of specimens, the site environment, the use of shells as artifacts or cultural material, and burnt marks from the contact with archaeological hearths are criteria used for distinguishing the shells from the occupational context (e.g. Prous Reference Prous1986; Barreto Reference Barreto1988; Moreira et al. Reference Moreira, Araujo, Confalonieri, Ferreira and Prous1991; Prous Reference Prous1991; Neves and Okumura Reference Neves and Okumura2005; Rosa Reference Rosa2006; Silva and Rosa Reference Silva and Rosa2006; Tenório et al. Reference Tenório, Afonso and Pinto2010; Figuti et al. Reference Figuti, Plens and DeBlasis2013; Ramos Jr Reference Ramos2014).
The presence of terrestrial snails’ shells in Brazilian shellmounds has been reported in many works (Table 1). These data draw attention to the large exploitation of such animals in prehistoric Brazil. However, in most studies, their origin is not properly discussed and they are often observed as possibly from natural occurrence even when criteria for differentiation exist (Rosa Reference Rosa2006). Even when they are recognized as archaeological remains, the choice of materials used for radiocarbon (14C) dating Brazilian shellmounds does not traditionally include terrestrial snails, but rather only charcoal, marine shells and bones.
Terrestrial Snails as Chronological Records
Charcoal and marine shells are the most commonly used materials for dating Brazilian shellmounds. Charcoal is found either in reliable archaeological contexts in hearths or scattered over the shellmounds, but it cannot always be associated with a specific context with confidence (Théry-Parisot et al. Reference Théry-Parisot, Chabal and Chrzavzez2010). Moreover, charcoal is susceptible to the “old wood effect,” in which it reflects the age of the wood instead of the time of the burning, which is especially problematic when long-lived trees are used in archaeological fires (McFadgen Reference McFadgen1982). Marine shells, in turn, are very abundant and obviously related to the mound construction, but have the drawback that ocean waters are depleted in 14C when compared to the coeval atmosphere and that the so-called marine reservoir effect (MRE) (Stuiver et al. Reference Stuiver, Pearson and Braziunas1986) is not known for most of the regions in Brazil. Therefore, 14C calibration results in inaccurate dates, either from the misuse of reservoir offsets from different locations or from the complete lack of such information. Other marine materials, such as fish otoliths, which are reflective of prehistoric fisheries, are also good options for dating, provided that the reservoir effect is known (Alves et al. Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015; Lopes et al. Reference Lopes, Bertucci, Rapagnã, de Almeida Tubino, Monteiro-Neto, Tomas, Tenório, Lima, Souza, Carrillo-Briceño, Haimovici, Macario, Carvalho and Aguilera2016). Human and animal bones represent important records in chronological studies, but collagen is rarely preserved in many areas of Brazil, due to soil acidity (Okumura and Eggers Reference Okumura and Eggers2008; Roksandic et al. Reference Roksandic, de Souza, Eggers, Burchell and Klokler2014; Strauss Reference Strauss2016).
Given the drawbacks of these materials, terrestrial snails’ shells are desirable as an alternative source of 14C-datable material. As long as terrestrial snails are in isotopic equilibrium with the atmosphere and their shells behave as closed systems, they can yield reliable dates (Pigati Reference Pigati2015). Studies in other regions of the world have shown that some terrestrial snails have presented apparent ages due to the incorporation of old carbon from limestone (e.g. Goodfriend and Stipp Reference Goodfriend and Stipp1983). The amount of old carbon incorporated varies between taxa and between populations of a single taxa living in different habitats (Goodfriend et al. Reference Goodfriend, Ellis and Toolin1999; Pigati et al. Reference Pigati, Rech and Nekola2010; Pigati Reference Pigati2015). In southern Italy, Quarta et al. (Reference Quarta, Romaniello, D’Elia, Mastronuzzi and Calcagnile2007) observed apparent ages of circa 1000 yr for two species of land snails, while 78% of the 46 species studied by Pigati et al. (Reference Pigati, Rech and Nekola2010) from carbonate areas did not contain old carbon. Many studies were developed with the aim of discussing the difficulties underlying this approach (e.g. Tamers Reference Tamers1970; Yates Reference Yates1986; Goodfriend et al. Reference Goodfriend, Ellis and Toolin1999; Zhou et al. Reference Zhou, Head, Wang, Donahue and Jull1999; Pigati et al. Reference Pigati, Rech and Nekola2010; Pigati Reference Pigati2015), and Pigati (Reference Pigati2015) highlights that for terrestrial carbonates to be used for 14C dating, they should be repeatedly evaluated in different depositional settings, habitats and climate regimes.
Figuti et al. (Reference Figuti, Plens and DeBlasis2013) reported shell dates for riverine shellmounds at the Ribeira Valley in southeast Brazil, where most of the shells are from land snails, but they do not specify which species were dated or which of the shells are from aquatic or terrestrial origin. Mascarenhas (Reference Mascarenhas2008) dated Megalobulimus shells from natural context in caves of the Peruaçu Valley in Minas Gerais to study river flood episodes. In Argentina, Fontana (Reference Fontana2007) dated Heleobia parchappii (d’Orbigny 1835) land snails to study a palaeolagoon in Buenos Aires. In southern Peru, Unkel et al. (Reference Unkel, Kadereit, Mächtle, Eitel, Kromer, Wagner and Wacker2007) dated Scutalus sp. (Broderip 1832) snails that indicated appreciable geomorphic activity from 1390 to 1714 cal AD and Mächtle et al. (Reference Mächtle, Unkel, Eitel, Kromer and Schiegl2010) dated both Scutalus and Pupoides sp. (Pfeiffer 1854) as evidence for a humid period late Pleistocene and early Holocene.
The first 14C dates of specific terrestrial snails from Brazilian coastal archaeological sites were reported by Carvalho et al. (Reference Carvalho, Macario, Oliveira, Oliveira, Chanca, Alves, Souza, Aguilera and Douka2015), who compared archaeological charcoal, land snails, and marine mollusks from the same context with the aim of calculating corrections for the marine reservoir effect for the region. The study has shown that the terrestrial mollusk Thaumastus achilles (Pfeiffer 1852) provides reliable atmospheric dates for the Manitiba shellmound, in Saquarema, RJ. Macario et al. (Reference Macario, Alves, Carvalho, Oliveira, Ramsey, Chivall, Souza, Simone and Cavallari2016a) evaluated the use of the genera Thaumastus and Megalobulimus as representatives of the atmospheric carbon reservoir. Shells that were mostly collected while still alive and have known collection dates (1948–2004 AD, covering the nuclear testing period), were analyzed and the results agreed within less than 15 years with the bomb-peak curve for zone 1–2 (Hua et al. Reference Hua, Barbetti and Rakowski2013; Macario et al. Reference Macario, Alves, Carvalho, Oliveira, Ramsey, Chivall, Souza, Simone and Cavallari2016a). Archaeological shells from Megalobulimus and Thaumastus from the Usiminas shellmound, on Cabo Frio Island, were compared with marine species in Macario et al. (Reference Macario, Alves, Chanca, Oliveira, Carvalho, Souza, Aguilera, Tenório, Rapagnã, Douka and Silva2016b) as a means of studying the local marine reservoir effect in the region. However, in that work, no comparison was made between charcoal and terrestrial snails. In this paper, we report the results for charcoal samples from the Usiminas shellmound and terrestrial snails shell samples from the Saquarema shellmound, comparing the data with corresponding atmospheric representatives from literature (Alves et al. Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015; Macario et al. Reference Macario, Alves, Chanca, Oliveira, Carvalho, Souza, Aguilera, Tenório, Rapagnã, Douka and Silva2016b). Despite the abundance of Thaumastus and Megalobulimus terrestrial snails, this is the first work in South America aiming to establish shellmounds chronology and test its validity based on shells from such genera.
MATERIAL AND METHODS
The samples dated in the present work are from the Usiminas shellmound (Figure 1), on the Cabo Frio Island (23º01′S, 42°00′W) and from the Saquarema shellmound in Saquarema city (22°55′S, 42°30′W), north of Guanabara Bay, in the state of Rio de Janeiro (Figure 3). The former is situated on a flat area on the top of a dune at 53 m asl (above sea level) and has a total area of 1920 m² (Tenório et al. Reference Tenório, Afonso and Pinto2010) whereas the latter, located between the Saquarema Lagoon and the sea (Kneip Reference Kneip1998), is at 2.17 m asl, with 4 m height and perimeter of 180 m (Guimarães Reference Guimarães2007). This sandy environment between lagoon and sea is also the location of a third site, the Manitiba I shellmound, which is the largest shellmound in the Saquarema region, with its base 3.36 m asl and total area of 6000 m² (Kneip Reference Kneip2001; Carvalho et al. Reference Carvalho, Macario, Oliveira, Oliveira, Chanca, Alves, Souza, Aguilera and Douka2015). This settlement was the object of study by Carvalho et al. (Reference Carvalho, Macario, Oliveira, Oliveira, Chanca, Alves, Souza, Aguilera and Douka2015), whose data are also discussed here. A notable difference among the three sites (Figure 3) is that while the Saquarema region offers strategic proximity to food resources, Cabo Frio Island offers great visibility and proximity to rock sources for tool fabrication instead (Kneip Reference Kneip2001; Tenório et al. Reference Tenório, Afonso and Pinto2010).
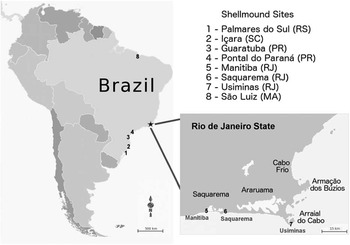
Figure 3 Map of South America with the locations of studied sites in Rio de Janeiro (5–7) and other regions where land snails were recorded in shellmounds (1–4, 8). Saquarema (6) and Usiminas (7) are the sites dated in this study and Manitiba (5) is used for comparison.
The studied sites were chosen for the availability of terrestrial snails in association with charcoal from the same archaeological context within the collection of the National Museum of Rio de Janeiro Federal University. Although sample collection directly in the field would reduce variability sources, this is no longer possible for some shellmounds in the state of Rio de Janeiro, such as the Saquarema shellmound, which has been destroyed. For the Usiminas and the Manitiba shellmounds, new excavations are possible and will be supported by the success of the present work.
Six archaeological layers in the Usiminas site were identified in two excavations performed by Tenório et al. (Reference Tenório, Afonso and Pinto2010), who describe the archaeological material recovered from each of them in detail. Particularly, the fifth layer had abundant malacological remains, with intense presence of the land snails Megalobulimus terrestris (Spix 1827) and Thaumastus achilles (Pfeiffer 1752), followed by marine mollusks associated with urchin spines, fish bones and archaeological artifacts (Macario et al. Reference Macario, Alves, Chanca, Oliveira, Carvalho, Souza, Aguilera, Tenório, Rapagnã, Douka and Silva2016b; Lopes et al. Reference Lopes, Bertucci, Rapagnã, de Almeida Tubino, Monteiro-Neto, Tomas, Tenório, Lima, Souza, Carrillo-Briceño, Haimovici, Macario, Carvalho and Aguilera2016). The samples analyzed in the present work are from sector 1C at 30–50 cm depth (Layer V), and from sector 1D at 80–90 cm depth (Layer VI).
Excavations undertaken in 1999 and subsequently in 2000 revealed seven archaeological layers in the Manitiba I settlement. Only two funerary structures were recovered, in contrast to other shellmounds in the same area, with at least 19 funerary structures each. The spatial distribution of artifacts and other archaeological remains characterize this shellmound as a living place resulting from intense domestic use and ceremonial activities (Kneip Reference Kneip2001). The characteristics of each layer are reported in Kneip (Reference Kneip2001). The results presented here are from Carvalho et al. (Reference Carvalho, Macario, Oliveira, Oliveira, Chanca, Alves, Souza, Aguilera and Douka2015) and the analyzed samples belong to Layer IV.
The Saquarema shellmound, in turn, has less documentation of its excavations. Discovered in 1932, it has been exploited and partially destroyed due to the extraction of material for building purposes. An excavation was conducted in this site in 1993, when three layers of archaeological settlement were identified. However, the most well recorded intervention in this site is a larger scale excavation performed in 1994 (Kneip Reference Kneip1995). Kneip (Reference Kneip1995) observed that the limits of human occupation were difficult to distinguish. In the work of Alves et al. (Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015), where different species of marine mollusks, fish otolith, and charcoal samples from sector B were measured, it was shown that, within a same sector, there was no correlation between age and depth, with the set of marine mollusk shells showing larger dispersion of results than charcoal and fish otolith samples. In the present work, we compare six charcoal samples from 30–100 cm depth in sector B with four Thaumastus shells from 50–90 cm in sector E.
Calcite and aragonite contents were measured for powdered, raw shell samples from Saquarema and Usiminas at the X-Ray Diffraction Laboratory of the Fluminense Federal University. To confirm the efficiency of the chemical treatment for 14C dating, for one of the samples (the one with largest calcite content), we repeated the measurement after HCl etching to remove 50% of mass. We employed a Bruker AXS D8 Advance (Cu Kα radiation, 40 kV, 40 mA) operation in a Bragg–Brentano θ/θ configuration with the diffraction patterns being collected in a flat geometry with steps of 0.02 degrees and accumulation time of 2.0 s per step using a PSD detector (Bruker AXS LynexEye model). The XRPD data were refined following the Rietveld method with the TOPAS 5.0 software.
Sample preparation and analysis were performed at the Radiocarbon Laboratory of the Fluminense Federal University (LAC-UFF) in Brazil (Macario et al. Reference Macario, Gomes, Anjos, Carvalho, Linares, Alves, Oliveira, Castro, Chanca, Silveira, Pessenda, Moraes, Campos and Cherkinsky2013). Charcoal samples were chemically pre-treated at 90°C using acid-base-acid (1.0M HCl – 1.0M NaOH – 1.0M HCl) followed by combustion in independently sealed quartz tubes with CuO and Ag wire. Mollusk shells were cleaned and ultrasonicated in ultrapure water (18 MΩ), and a 40mg fragment was cut with a razor blade. Carbonate samples were etched (aiming to remove 50% in mass) with 0.1M HCl and hydrolyzed in H3PO4. The gas obtained from this reaction was purified by means of cryogenic traps and the carbon dioxide was transferred to Pyrex tubes containing Zn and TiH2 and an inner tube with Fe. Graphitization took place in a muffle oven at 520°C for 7 h (Macario et al. Reference Macario, Oliveira, Carvalho, Santos, Xu, Chanca, Eduardo, Jou, Oliveira, Pereira, Moreira, Muniz, Linares, Gomes, Anjos, Castro, Anjos, Marques and Rodrigues2015). Measurements were performed in a 250 kV Single Stage AMS System from NEC.
Calibration was performed with the software OxCal v 4.2.4 (Bronk Ramsey Reference Bronk Ramsey2009a) using the Southern Hemisphere atmospheric curve SHCal13 (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013). In order to statistically test the similarity of results for charcoal/mollusk groups, a general outlier model with a Student t-distribution (Bronk Ramsey Reference Bronk Ramsey2009b) was used while constraining each group of samples (same context) to within a single phase.
RESULTS AND DISCUSSION
Radiocarbon dating results for both terrestrial mollusk shells and charcoal are shown in Table 2. We present two new charcoal and one new terrestrial snail dates for the Usiminas shellmound and four new terrestrial snail dates for the Saquarema shellmound, plus the results from Carvalho et al. (Reference Carvalho, Macario, Oliveira, Oliveira, Chanca, Alves, Souza, Aguilera and Douka2015) for the Manitiba I shellmound.
Table 2 Archaeological sites, type of material, position within the sites, LAC-UFF reference codes, aragonite content, and 14C results of studied samples.

A graphical representation of radiocarbon dating results from Table 2 is shown in Figure 4, where the boxes represent the 14C age values obtained for each material: shells of Thaumastus sp. and Megalobulimus sp. mollusks and charcoal for each studied site. The space inside the box indicates the degree of dispersion in the data, the error bars represent the minimum and maximum of all data within the group and the open squares and the horizontal line inside the boxes represent the mean and the median, respectively. There are no significant differences between Thaumastus sp. and charcoal samples’ results from Manitiba I site (Layer IV). Although the available samples were rather limited for this site, the integrity of this shellmound is preserved and future excavations are possible. This site has the presence of sterile layers between the occupational layers, which assures no disturbance of a layer by the one overlying it. Unfortunately, this important feature is not present in the Usiminas or Saquarema shellmounds.

Figure 4 BoxPlot of the radiocarbon ages of charcoal and land snails, Thaumastus and Megalobulimus, from the Manitiba I, Saquarema and two layers of the Usiminas shellmounds. The size of the box indicates the degree of dispersion in the data, the error bars represent the minimum and maximum of all data within the group and the open squares and the horizontal line inside each box represent the mean and the median, respectively.
Concerning the contemporaneity among samples from the same context, variability in results may reflect the long occupational periods of the native populations, but the shellmounds archaeostratigraphy does not always allow more detailed inferences (Alves et al. Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015; Macario et al. Reference Macario, Alves, Chanca, Oliveira, Carvalho, Souza, Aguilera, Tenório, Rapagnã, Douka and Silva2016b). Shallow sites often present mixing of occupational layers due to human activities, such as cleaning of hearths, leveling of terrain or digging of graves (Macario et al. Reference Macario, Oliveira, Carvalho, Santos, Xu, Chanca, Eduardo, Jou, Oliveira, Pereira, Moreira, Muniz, Linares, Gomes, Anjos, Castro, Anjos, Marques and Rodrigues2015). Although the relationship between the processes of mound construction and funerary activities cannot be extended to all sites (Peixoto Reference Peixoto2008; Scheel-Ybert et al. Reference Scheel-Ybert, Afonso, Barbosa-Guimarães, Gaspar and Ybert2009; Klokler et al. Reference Klokler, Villagrán, Giannini, Peixoto and Deblasis2010), funerary rituals have probably structured the way of life of fishers and gatherers, the builders of shellmounds (Gaspar et al. Reference Gaspar, DeBlasis, Fish and Fish2008, Reference Gaspar, Klokler and Bianchini2013). Ceremonial activities such as food and artifacts offerings, treatment of bodies with pigments and replacement of specific bones were frequent (Gaspar et al. Reference Gaspar, DeBlasis, Fish and Fish2008). Secondary burials and especially multiple burials (e.g. adult and child), common in shellmound culture, can interfere with archaeostratigraphy (Gaspar et al. Reference Gaspar, DeBlasis, Fish and Fish2008). In the case of primary burials, it is possible to observe individuals with articulated bones so that their position can be inferred and so is that of the materials disposed in ritual association. It can be observed in archaeostratigraphy if the bodies were buried in graves or simply disposed on the ground.
In Saquarema, charcoal fragments from several depths in sector B have statistically similar results. For this site, no correlation was observed with depth for either otolith or marine shells analyzed by Alves et al. (Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015), who have shown the difficulty in defining the occupational layers. For these reasons, any attempt to constrain the results based on depth would be pointless. For the purpose of this study, even though the available samples are from two distinct sectors, it was possible to observe that the whole extension of sector B (up to 110 cm deep) was from a single occupational period, contemporaneous with 80–90cm-deep land snails samples from sector E. At 30–50 cm depth, samples are about two centuries younger and probably represent a different occupation.
In the Usiminas shellmound, mixing of different time ranges in superficial layers (up to 20 cm) was verified by Macario et al. (Reference Macario, Alves, Chanca, Oliveira, Carvalho, Souza, Aguilera, Tenório, Rapagnã, Douka and Silva2016b) with dates from 1900±45 to 1305±35 BP for terrestrial snails. In that work, the deepest samples from sector F3_1C (main excavation) were consistent with those from sector NT (secondary excavation) with dates ranging from 1675±35 to 1505±30 BP. In the present work, for the group of samples from sector F3_1C 30-50 cm (layer V) both terrestrial mollusks species results agree with each other and with that of charcoal. New results from even deeper samples (>80 cm) in sector F3_1D have disclosed a much older occupational layer, represented by one individual of terrestrial mollusks Megalobulimus sp. and a charcoal fragment (layer VI).
Concerning the validity of the presented results, a few issues should be discussed other than the contemporaneity among each group of samples from the same archaeological layer, such as the longevity of tree species from which charcoal was originated, recrystallization of shell carbonates or the incorporation of dead carbon from limestone.
The very essence of comparing charcoal with other materials relies on the validity of charcoal dating. The old wood effect to which charcoal is subject to may be responsible for age shifts of decades or even centuries depending on the plant species (McFadgen Reference McFadgen1982). Anthracological analysis is desirable to avoid errors caused by the old wood effect. For the studied charcoal samples, those from the Saquarema shellmound had been anthracologically analyzed (Alves et al. Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015). The plant families identified are shown in Table 2 and represent sandbank vegetation, which is not usually composed of long-lived species (Alves et al. Reference Alves, Macario, Souza, Aguilera, Goulart, Scheel-Ybert, Bachelet, Carvalho, Oliveira and Douka2015).
Like marine carbonate, terrestrial shells can be subject to recrystallization. It has been observed that for old samples, aragonite can transform into calcite, allowing exogenous carbon atoms to be incorporated (Douka et al. Reference Douka, Hedges and Higham2010). In this case, monitoring with x-ray diffraction, for example, can give information regarding the aragonite/calcite ratios. Samples from the Usiminas and Saquarema shellmounds were analyzed and the results were between 90.4 and 96.2% aragonite in raw material (before acid etching). Since sample preparation for 14C measurements included a 50% etching, we adopted the same procedure and re-analyzed sample LACUFF 140552 (which had the largest calcite content). The result was a decrease in calcite content from 9.6% to 0.9%, indicating that the chemical treatment was successful. Given the radiocarbon age of this sample, even if all the re-crystalized carbon was modern, the difference in age would be less than the measurement uncertainty.
Another potential problem is the incorporation of dead carbon from limestone, but the study region does not have carbonate rocks (Jansen et al. Reference Jansen, Cavalcanti and Lamblém2012). Moreover, Macario et al. (Reference Macario, Alves, Carvalho, Oliveira, Ramsey, Chivall, Souza, Simone and Cavallari2016a) have shown that a set of terrestrial mollusks from the studied genera were not affected by the dead carbon effect even in samples collected from karstic, high limestone content regions, such as São Domingos in the state of Goiás and from carbonate rock exploitation areas, such as Aracruz and Baixo Gandú in the state of Espírito Santo. Although Pigati et al. (Reference Pigati, Rech and Nekola2010) claim that larger snails would require to scrape limestone to fulfill their need for calcium, none of the large snails from the Megalobulimus genera (e.g. up to 105 mm in length for the Megalobulimus paranaguensis reported by Miranda and Fontenelle Reference Miranda and Fontenelle2015) from Macario et al. (Reference Macario, Alves, Carvalho, Oliveira, Ramsey, Chivall, Souza, Simone and Cavallari2016a) work has shown signs of dead carbon incorporation.
Other than the discussed factors, evaluating the contemporaneity of 14C results should also take into account the influence of calibration curves. Despite the fact that both charcoal and land snails’ results should be calibrated with the atmospheric curve, in our case, the Southern Hemisphere SHCal13 curve (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013) wiggles and plateaus will interfere with the probability distribution of calibrated results.
Figure 5 shows calibrated ages for each of the studied sites. Both terrestrial mollusk taxa present similar results to their respective charcoal samples. From the work of Carvalho et al. (Reference Carvalho, Macario, Oliveira, Oliveira, Chanca, Alves, Souza, Aguilera and Douka2015) on the Manitiba I shellmound, the results from charcoal agreed with those from terrestrial snail shells, showing that terrestrial gastropods can be used to yield reliable 14C dates instead of charcoal. From the calibrated results, the occupation of the Manitiba shellmound (Layer IV) has happened within the range from 4.2 to 3.7 ka cal BP (95.4%), contemporary to the Saquarema shellmound in the same municipality, occupied within the period from 4.3 to 3.6 ka cal BP (95.4%). For the Usiminas shellmound, the terrestrial shells have demonstrated results that are as reliable as charcoal samples for distinguishing between different occupational layers. Samples from sector 1C at 30–50 cm depth are from 1.6 to 1.3 ka cal BP (95.4%), while samples from sector 1D at 80–90 cm depth are from 2.3 to 2.1 ka cal BP (95.4%). A phase model constraining each group of samples to within a single phase has a general agreement of 97%, with only two outliers out of 22 dates, yielding minimum individual agreement of 74% and 7% posterior outlier probability for Saquarema shells.

Figure 5 Probability distributions for the calibrated radiocarbon dates of charcoal (black) and land snails (grey) (see Table 2). LACUFF reference codes, conventional dates, agreement within the model and outlier probability are given for each sample. Calibration was performed with the software OxCal v 4.2.4 (Bronk Ramsey Reference Bronk Ramsey2009a) using the Southern Hemisphere atmospheric curve SHCal13 (Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013).
FUTURE PERSPECTIVES
The study of shellmounds on the Brazilian coast remains an important subject for both archaeological and environmental studies. Not only do shellmounds allow for studying social behavior of humans, they also give insights into paleoenvironmental aspects, as they contain sets of both botanical and zoological remains representative of the existing fauna and flora at the time they were built (Fürsich Reference Fürsich1995; Scheel-Ybert et al. Reference Scheel-Ybert, Klökler, Gaspar and Figuti2006; Lindbladh et al. Reference Lindbladh, Brunet, Hannon, Niklasson, Eliasson, Eriksson and Ekstrand2007; Froyd and Willis Reference Froyd and Willis2008). In the case of coastal sites, it is always important to increase statistics as much as possible so that outliers, interspecies variations and bioturbation may be identified, due to the many challenges that these studies may present (e.g. 14C age variability due to anthropic interference and post-depositional effects, longevity or feeding habits of different species, statistical fluctuations in measurements, calibration curve plateaus, and lack of knowledge of local reservoir effect).
As discussed before, the samples from the present study came from museum collections and were therefore associated with a labeled position from the site, which could introduce a source of variability. Ideally, for comparative works, shells and charcoal should be carefully selected close together in the field. However, as a preliminary work, our aim with this study was to show the potential use of the studied mollusk genera in archaeological studies, especially shellmounds, where so many difficulties must be overcome to establish an accurate chronology. The greatest advantages of using land snails in future shellmound studies include their high abundance in some Brazilian archaeological sites, which adds to the robustness of statistical models, and their independence from marine reservoir effects, a traditional obstacle in studies of coastal sites.
Successful work that demonstrated that terrestrial mollusks are representative of the atmospheric 14C reservoir enabled this study to compare 14C dates from archaeological charcoals and terrestrial mollusk shells in three shellmounds on the southeast coast of Brazil. These results show that the snails were not only contemporary to the archaeological context, but also represent an important alternative for chronological and multiproxy studies. Agreement in 14C dates of charcoals and terrestrial shells was obtained for all the studied sites. The results are encouraging enough to consider using terrestrial mollusk shells as a source of 14C dates for Brazilian shellmounds. Even more importantly, these results suggest that in sites with convenient archaeostratigraphy (e.g. with gaps between occupational layers), abundant terrestrial snails could be used for establishing chronological sequences; ongoing research in Manitiba I shellmound is being performed with this aim.
ACKNOWLEDGMENTS
The authors would like to thank Brazilian financial agencies CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, 305079/2014-0) and FAPERJ (Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro, E-26/111.278/2014, E-26/110.138/2014) for their support. We are grateful to Marcelo Muniz for the help with sample preparation. We also thank Luciano Rapagña and Bruna Brandão for the photographs, Daniel Cavallari, José Fontenelle, and Luiz Simone for information on land snails’ ecology, and Katy Sparrow for revising the text. We acknowledge the reviewers and Associate Editor Cristine Hatté for their very useful comments and suggestions, which helped to improve this work.







