INTRODUCTION
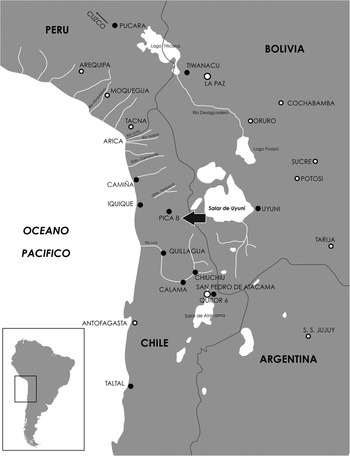
A robust chronology is essential for the interpretation of patterns found in the archaeological record and, by implication, the social processes underlying them. In this paper, we provide an example focused on the cemetery of Pica 8, one of a group of oasis sites on the edges of the Atacama Desert, northern Chile (Figure 1). The site belongs to the Pica-Tarapacá Cultural complex of the Late Intermediate Period (LIP), AD 900–1450 (Núñez Reference Núñez1984; Uribe Reference Uribe2006). This complex extends from the Camiña River to the north, the Loa River to the south, and from the Pacific coast to the west to the Andean pre-cordillera and highlands to the east (Uribe Reference Uribe2006). Even though Pica 8 is located approximately 90 km from the coast, the archaeological evidence (fish and shellfish remains, manta ray eggs, marine bird feathers) suggests connections between the oases and the seaside (Núñez Reference Núñez1984; Zlatar 1984; Moragas 1995; Briones et al. 2005; Uribe Reference Uribe2006).

Figure 1 Map of northern Chile. The cemetery of Pica 8 is indicated by an arrow. Map slightly modified from Santana-Sagredo et al. (Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a).
A total of 254 burials were excavated from the cemetery (Núñez Reference Núñez1965, Reference Núñez1984). Thus far, a total of 82 skeletons have been analyzed for sex and age estimations as well as paleopathology and violence-related trauma (Retamal et al. 2012; Pacheco and Retamal 2017). Stable isotope analysis on 30 human skeletons revealed a surprising degree of dietary variability, with some individuals interpreted as consuming significant amounts of marine foods (despite the site’s location approximately 90 km inland), while the diets of other individuals appeared entirely terrestrial (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). Moreover, the graves of individuals exhibiting these different diets appeared to be grouped spatially into different sectors of the cemetery. A question then arose regarding the extent to which the observed isotopic-spatial pattern was diachronic or “synchronic” (see below). The available chronology for the site has relied mainly on pottery typology, which lacks sufficient resolution in this region to confidently distinguish between these alternatives, although lending more support to the second scenario of considerable dietary heterogeneity within the community using Pica 8 for burial purposes. Here, we report the results of a radiocarbon dating program (n=23) focused on this question.
The isotopic results obtained for the Pica 8 human remains revealed the existence of considerable variability in collagen and apatite δ13C, and in collagen δ15N values (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). At least three dietary groupings could be identified in Pica 8. The first exhibits high δ13C values for bone collagen and apatite together with high δ15N values, suggesting a marine-C4 (maize) based diet. The second grouping of high δ13C but lower δ15N values (although still relatively high at 14–15‰) suggests a focus on maize with a lower reliance on marine resources. Finally, the third grouping suggests a terrestrial-C3 based diet with the lowest values for both carbon and nitrogen isotopes. Some of this variability can be related to the sectors—nine in total, designated A through J (Núñez Reference Núñez1984; Zlatar 1984)—in which the burials were found. In our previous study, individuals from sectors A, B, D, E, F, G, and I were analyzed (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). Sector I contains mainly individuals with very high δ15N values of more than 20‰, while most individuals in Sectors A and G exhibit lower values of around 14–15‰ (Figure 2). Other sectors, such as D, are more variable, containing individuals with both high and low δ15N values.

Figure 2 Bivariate plot showing δ13C and δ15N values from bone collagen for Pica 8, identified by sector (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). Individuals selected for radiocarbon dating are indicated by their inventory numbers.
While the cemetery can be confidently placed in the LIP, this period spans at least 500 yr, and it is possible that the observed isotopic variability is related to changing subsistence practices over time. This change could be due to increased contacts with the coast (clearly attested by finds of desiccated marine fish and shellfish, manta ray eggs, and marine bird feathers), or to intensification of farming practices based on manuring of crop staples [as suggested by experimental studies on modern crops with seabird guano by Szpak et al. (Reference Szpak, Millaire, White and Longstaffe2012a, Reference Szpak, Longstaffe, Millaire and White2012b)]. The alternative is that the community at Pica 8 comprised a cosmopolitan mix of groups of people with different origins and diets, many of whom maintained their distinct cuisines. Resolving the question of which explanation is more likely will greatly improve our understanding of the Pica-Tarapacá Complex, and could (assuming the “synchronic” view is supported) provide new avenues of research into the conditions under which such “sustained multiculturalism” arose, and how and why it was maintained over multiple generations, if not centuries.
Central to choosing between the two alternatives set out above is a robust chronology. What prevents this from being a straightforward task is the complication of the inferred high marine contribution to the diets of those individuals with elevated δ15N values, above 18‰ (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). Moreover, the marine reservoir effect (MRE) is known to be extremely complex off the west coast of South America, being heavily influenced by deep-water upwelling, the intensity of which varies spatially and temporally, and by shifts in oceanic circulation due to influences such as the El Niño Southern Oscillation (ENSO) (Owen Reference Owen2002; Fontugne et al. Reference Fontugne, Carre, Bentaleb, Julien and Lavallee2004; Ortlieb et al. Reference Ortlieb2011; Latorre et al. Reference Latorre, De Pol-Hoz, Carter and Santoro2015).
The main aim of this study is to resolve the chronology of the Pica 8 cemetery and, in particular, whether the observed isotopic/dietary variability is diachronic or “synchronic.” The term “synchronic” in this context is not meant to imply that all the burials are necessarily exactly contemporary, but rather that the observed isotopic variation bears no relationship with chronology. The underlying inference, however, is that the rejection of the diachronic explanation implies isotopic/dietary heterogeneity within a single “living” community. To achieve this, 9 paired 14C determinations were undertaken on human bone and camelid textiles in order to address the marine reservoir offset for this region and period. We expect human collagen to be most affected by the MRE, whereas the camelid textiles should be entirely unaffected. In addition to the paired dates, 5 additional burials were selected for 14C dating. No textiles or other appropriate terrestrial dating materials could be associated with these individuals, but they were included in order to provide more comprehensive spatial coverage of the various sectors of the Pica 8 cemetery, which previous research suggested may have been a factor in the observed isotopic/dietary variability (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a).
Archaeological and Chronological Context of the Pica-Tarapaca Cultural Complex
The Late Intermediate Period (AD 900–1450) in northern Chile falls between the Tiwanaku State (preceding) and Inca (following) Empire. However, the Tarapacá region was not within the Tiwanaku sphere of influence during the Middle Horizon, AD 400–900 (Berenguer and Dauelsberg Reference Berenguer and Dauelsberg1989). Instead, the LIP here was preceded by an extended Formative Period, from approximately 1000 BC to AD 900. The LIP populations were direct heirs of the Formative Period groups that inhabited the Tarapacá lowlands. During the LIP these populations expanded their occupation to the highlands. The LIP lacked overarching sociopolitical structures, being characterized instead by segmented and autonomous communities (Uribe Reference Uribe2006). In order to provide the economic necessities, increased mobility and trade occurred between these communities, thus connecting highland, inland, and coastal zones (Schiappacasse et al. Reference Schiappacasse, Castro and Niemeyer1989). However, it is still not clear how and at what scale human mobility occurred. LIP social organization, settlement and mobility have conventionally been interpreted following models that are strongly influenced by ethnohistorical accounts involving the imposition of colonies from the highlands to the valleys and coast (Murra Reference Murra1972) or the exchange of goods controlled by llama caravans from the highlands (Núñez and Dillehay Reference Núñez and Dillehay1995). There is an ongoing debate on this matter.
Much of the current LIP chronological framework for the Pica-Tarapacá Complex is based on ceramic seriation, supported by thermoluminescence (TL) determinations (Uribe et al. Reference Uribe, Sanhueza and Bahamondes2007). The main pottery type that characterizes this cultural complex is known as Pica Charcollo (PCH) with an age range of AD 950–1470, also including variants such as Pica Chiza (PCHZ) and Pica Gris Alisado (PGA). Only six 14C dates on archaeological material have been published so far for the interior part of the Tarapacá region, specifically from the Pica 8 cemetery itself (Núñez Reference Núñez1976). However, two of these (on maize cobs) give calibrated dates post-AD 1500, too recent for what is clearly a pre-Columbian cemetery; they likely represent intrusive material. A third date on a camelid textile from tomb 6 shows a determination between cal AD 968–1270 (IVIC-173: 930±90 BP). The final three determinations are on human bone collagen, specifically individuals SDT11 (Beta 220922: 1050±40 BP: AD 900–1030), B0447 (Beta-220923: 900±40 BP: AD 1030–1230), and B0438 (Beta-220924: 810±40 BP: AD 1170–1280) (Uribe et al. Reference Uribe, Sanhueza and Bahamondes2007). All four would thus appear to fall within the LIP. However, since there is evidence of marine dietary input at Pica 8 (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a), these dates may be subject to the MRE. No δ15N measurements are associated with these dates, limiting their usefulness. Overall, given the paucity of absolute dates, our results will contribute to building a more robust chronological framework for the LIP in northern Chile.
Marine Reservoir Effect in Northern Chile
The marine reservoir effect (MRE) is defined as the difference in the 14C content present in terrestrial and marine organisms, with the latter being depleted in 14C (Stuiver et al. Reference Stuiver, Pearson and Brazuinas1986; Stuiver and Brazuinas Reference Stuiver and Braziunas1993). In contrast to terrestrial environments and organisms, the CO2 present in the ocean is not in equilibrium with the atmosphere, being influenced by sources of “old” organic and inorganic carbon. Thus, dates obtained for modern marine organisms will be significantly older compared to those from the terrestrial biome. This average global ocean MRE difference has been estimated at around 400 yr (Stuiver et al. Reference Stuiver, Pearson and Brazuinas1986; Stuiver and Brazuinas Reference Stuiver and Braziunas1993), but local deviations from this can vary significantly depending on upwelling intensity and latitude. Inputs from freshwater systems can have their own reservoir offsets (Fernandes et al. Reference Fernandes, Rinne, Nadeau and Grootes2014; Schulting et al. Reference Schulting, Bronk Ramsey, Goriunova, Bazaliiskii and Weber2014) but this is not a factor in northern Chile, which has no significant watercourses. This additional local variation, designated ∆R, needs to be considered when correcting the radiocarbon dates for any MRE in addition to the global R average (Stuiver et al. Reference Stuiver, Pearson and Brazuinas1986; Stuiver and Brazuinas Reference Stuiver and Braziunas1993).
Northern Chile and southern Peru are characterized by a strong upwelling system (Ortlieb et al. Reference Ortlieb2011; Latorre et al. Reference Latorre, De Pol-Hoz, Carter and Santoro2015). The Humboldt Current is characterized by high productivity and biodiversity, due to the high nutrient content of upwelled waters. Upwelling also brings dissolved inorganic carbon depleted in 14C from the ocean bottom to the surface (Latorre et al. Reference Latorre, De Pol-Hoz, Carter and Santoro2015). There are relatively limited data pertaining to the region’s ∆R values: the 14CHRONO Marine Reservoir Database (http://calib.qub.ac.uk/marine/) shows seven data points for northern Chile and southern Peru, with modern local ∆R values ranging from less than 50 yr up to almost 400 yr. Most of the information available in the database comes from Ortlieb et al. (Reference Ortlieb2011), who provide the most complete study on the MRE for northern Chile and southern Peru. Using a total of 46 paired dates on marine and terrestrial samples their data show considerable variability in ∆R through time. During the early Holocene ∆R values average 511±278 yr, decreasing significantly during the middle Holocene to 226±98 yr, before increasing again in the late Holocene to 355±105 yr. The authors relate this variability to changes in upwelling and suggest a stronger influence of the ENSO during the late Holocene.
A recent study by Latorre et al. (Reference Latorre, De Pol-Hoz, Carter and Santoro2015) on archaeological shell middens from Caleta Vitor, northern Chile, shows ∆R values somewhat higher than those obtained by Ortlieb et al. (Reference Ortlieb2011) during the early and mid-Holocene, emphasizing the difficulties involved in estimating ∆R. Compounding this is the spatial as well as temporal variation. The upwelling system for northern Chile shows significant differences compared to central Chile. Carré et al. (Reference Carré, Jackson, Maldonado, Chase and Sachs2016) provide an average ∆R value of 168±69 yr for the late Holocene in central Chile, compared to 355±105 yr for northern Chile. The difference between the early Holocene of central and northern Chile is even higher, with ∆R values of 31±156 yr and 528±301 yr, respectively (Latorre et al. Reference Latorre, De Pol-Hoz, Carter and Santoro2015; Carré et al. Reference Carré, Jackson, Maldonado, Chase and Sachs2016). Caution needs to be taken when interpreting the radiocarbon dates possibly affected by the MRE, considering the high variation in the ∆R values along the Chilean coast.
MATERIALS AND METHODS
A total of 23 samples from Pica 8 were selected for 14C dating, comprising adult rib samples and one hair sample from 13 distinct individuals, 9 of which could be paired with associated textiles to address the MRE (Tables 1 and 2). Adults were chosen to avoid complications in the interpretation of δ15N values due to nursing effects. We were able to obtain a new bone sample from only one of the three individuals (B0447) previously dated by Uribe et al. (Reference Uribe, Sanhueza and Bahamondes2007).
Table 1 Carbon and nitrogen stable isotope values and radiocarbon years for human and textile paired samples.

Table 2 Carbon and nitrogen stable isotope values and 14C years for human single samples.

The burials at Pica 8 were found in a naturally mummified state with the bodies completely preserved. All the soft tissues were removed in the 1970s to facilitate osteological analysis (Aspillaga personal communication 2016). The human bone and hair samples reported here were previously analyzed for δ13Ccoll and δ15N, and δ13Cap (as appropriate). Approximately one-third are argued to show considerable consumption of marine resources, while others suggest a mixed marine and terrestrial diet, and a few others a largely terrestrial diet (Figure 2; Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a).
Nine camelid textile and fiber samples were selected to pair with human bone. These were all directly associated with the bodies, comprising tunics and wrappings for the deceased, a textile bag, a loincloth, a camelid hair bundle, and one example of a camelid “hairball” found inside the mouth of one individual [the practice of leaving hairballs inside the mouths of the deceased has been documented at a Formative Period coastal site at Pisagua (Agüero personal communication); Figure 3].

Figure 3 Pica 8 textiles: (a) tunic (A03827) associated with individual B0414; (b) hairball found inside mummy’s mouth (A03841), associated with individual B0451; (c) bag (costal) associated with individual B0492.
Given the excellent preservation at the site due to the extremely arid conditions of the Atacama, neither the bone nor textile samples had been treated with any consolidants or conservation materials. All the analyses were carried out at the Oxford Radiocarbon Accelerator Unit (ORAU), University of Oxford. Bone collagen samples, as well has hair and textile (camelid fibers) samples were pretreated following the protocols of Brock et al. (Reference Brock, Higham, Ditchfield and Bronk Ramsey2010). The human bone collagen samples as well as the hair keratin sample had been previously prepared for stable isotope analysis (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a) at the Research Laboratory for Archaeology and the History of Art (RLAHA), University of Oxford.
Two out of the four new bone collagen samples, together with three of those previously prepared in RLAHA, presented high C/N ratios above 3.45 and were considered unacceptable for dating following the stricter ORAU standards rather than DeNiro’s (1985) widely accepted range of 2.9 to 3.6 for dietary stable isotope research. Since the samples were very well preserved, the slight elevation in their C/N ratios may have been due to the presence of traces of lipids. Therefore, two solvent washes with acetone, methanol, and chloroform were performed on these five samples. The resultant C/N ratios decreased from >3.45 to between 3.1 and 3.2, so that all samples met the quality control criteria.
The samples were combusted in a CF-IRMS system – Carlo ERBA NA 2000 coupled to a gas source IRMS, Sercon 20/20 (Brock et al. Reference Brock, Higham, Ditchfield and Bronk Ramsey2010). CO2 gas was collected and graphitized, and analyzed in the HVEE AMS system at ORAU. The resulting 14C determinations were calibrated using the SHCal atmospheric curve (Hogg et al. Reference Hogg, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013) in OxCal v 4.2.4 (Bronk Ramsey Reference Bronk Ramsey and Lee2013). Statistical analyses were undertaken in The R Project for Statistical Computing (https://www.r-project.org) and SPSS v23.
RESULTS
The results are presented in Table 1 (paired samples) and Table 2 (unpaired samples). As expected, most dates fall inside the temporal range for the LIP (AD 900–1450) spanning from cal AD 993–1414. Unexpected, however, are the very limited offsets in 14C yr observed in four of the paired dates for associated textile and bone samples (Figure 4). This includes the strongly 15N-enriched individuals (with the highest reaching δ15N values of 21–22‰), which were expected to exhibit offsets on the order of some centuries with diets dominated by marine protein. In one pairing (individual B0451) the date for the textile material—a ball of camelid hair placed inside the mouth—is actually significantly older than the bone, by about 270 14C yr. This may be a curated object, or one encountered accidentally from an earlier context, and retained as an object of some ritual significance. In any case, it is excluded from further consideration.

Figure 4 Calibrated paired radiocarbon dates of human bone collagen/hair keratin and camelids textiles/fibers from Pica 8.
Leaving aside the above outlier, the largest offset observed between the paired human bone and textile dates is 127 14C yr. Three other pairings exhibit offsets of 109–120 yr, giving an average offset of 117±9 14C yr for this group of four. Surprisingly, the δ15N values for these individuals range widely, from 10.9‰ to 21.0‰, spanning nearly the entire range of those sampled for paired dating, as well as incorporating a substantial part of the variation in δ15N seen in the previous study (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). The remaining four paired dates show no offsets, i.e., they can be successfully combined using the R_combine function in OxCal (see Ward and Wilson 1978). This group includes the individual (B0455) with the highest δ15N value of 22.0‰. Single and multiple linear regression analyses show no statistically significant correlations between any of the human stable isotope measurements (δ13C on collagen and apatite, and δ15N on collagen) and the observed offsets in 14C yr (Figure 5). Surprisingly, however, significant positive correlations were observed between the textile δ13C values (R2=0.606, p<0.05) and δ15N (R2=0.572, p<0.05) values and the associated 14C offsets (Figure 6).

Figure 5 Bivariate plots for (a) δ15N values, (b) δ13Ccoll values, and (c) δ13Cap values and the offset in 14C yr calculated from the paired bone collagen and textile samples of Pica 8.

Figure 6 Bivariate plot showing δ13C and δ15N values for keratin from textiles and fibers from Pica 8, showing the positive relationship between 14C offset and stable isotope values.
Four of the five results for the remaining unpaired dates on human bone are largely consistent with the Late Intermediate Period (Figure 7). However, one individual (B0483) can be placed at the end of the Formative Period for the Tarapacá region. The results for two other individuals (B0424 and B0431) exhibit probability distributions crossing the boundary between the Late Formative Period and the LIP.

Figure 7 Calibrated dates for paired bone collagen/textile samples and unpaired bone collagen samples from Pica 8.
DISCUSSION
Paired 14C Dates and the MRE
The results indicate the presence of two groups, one with a relatively small but significant offset of 117±9 14C yr, and the other showing no offset at all (excluding the outlier, B0451). This is a surprising outcome, given the previously assumed importance of marine foods in the diets of some individuals at Pica 8, combined with the strong upwelling regime characterizing northern Chile’s coast. The estimated ∆R for the late Holocene along this coast is 355±105 yr (Ortlieb et al. Reference Ortlieb2011; Latorre et al. Reference Latorre, De Pol-Hoz, Carter and Santoro2015). When added to the global ocean MRE value of 400 yr, the combined offset should be on the order of 650–860 yr for an individual obtaining most or all of their protein from the sea [see Fernandes et al. Reference Fernandes, Rinne, Nadeau and Grootes(2014) on the use of 14C offsets as a dietary proxy]. Using this range, the observed average offset of 117±9 yr for four individuals implies a contribution of marine foods on the order of between 12.6% and 19.4%, i.e., the observed offset in the four paired dates as a proportion of the expected total marine reservoir offset in a 100% marine consumer, taking into account the errors associated with these estimates (Table 3).
 $$\eqalignno{ & {\rm Minimum}\;\,\%\,{\rm marine}{\equals}^{{14}} {{{\rm C}_{{{\rm minoffset}}} } \mathord{\left/ {\vphantom {{{\rm C}_{{{\rm minoffset}}} } {{\rm MRO}_{{{\rm max}}} }}} \right. \kern-\nulldelimiterspace} {{\rm MRO}_{{{\rm max}}} }} \cr & {\rm Maximum}\;\,\%\,{\rm marine}{\equals}^{{14}} {{{\rm C}_{{{\rm maxoffset}}} } \mathord{\left/ {\vphantom {{{\rm C}_{{{\rm maxoffset}}} } {{\rm MRO}_{{{\rm min}}} }}} \right. \kern-\nulldelimiterspace} {{\rm MRO}_{{{\rm min}}} }} $$
$$\eqalignno{ & {\rm Minimum}\;\,\%\,{\rm marine}{\equals}^{{14}} {{{\rm C}_{{{\rm minoffset}}} } \mathord{\left/ {\vphantom {{{\rm C}_{{{\rm minoffset}}} } {{\rm MRO}_{{{\rm max}}} }}} \right. \kern-\nulldelimiterspace} {{\rm MRO}_{{{\rm max}}} }} \cr & {\rm Maximum}\;\,\%\,{\rm marine}{\equals}^{{14}} {{{\rm C}_{{{\rm maxoffset}}} } \mathord{\left/ {\vphantom {{{\rm C}_{{{\rm maxoffset}}} } {{\rm MRO}_{{{\rm min}}} }}} \right. \kern-\nulldelimiterspace} {{\rm MRO}_{{{\rm min}}} }} $$
Table 3 Estimation of marine contribution to Pica 8 human diet based on the expected ∆R values for the Late Holocene (Ortlieb et al. Reference Ortlieb2011) and the average offset in 14C yr obtained for the paired bone-textile dates.

This result is substantially lower than what would have been predicted on the basis of the stable carbon and nitrogen isotope values (Figure 2). Moreover, the lack of any significant correlation between the stable isotope measurements and the observed 14C offsets is difficult to explain (see below). Certainly, earlier studies have found significant predictive relationships in other contexts (Price et al. Reference Price, Ambrose, Bennike, Heinemeier, Noe-Nygaard, Brinch Petersen, Vang Petersen and Richards2007; Schulting et al. Reference Schulting, Bronk Ramsey, Goriunova, Bazaliiskii and Weber2014).
These results challenge the previous interpretation of the high δ15N values (up to 24.4‰) at Pica 8 as indicating a substantial contribution of marine protein in the diets of these individuals. This was a reasonable assumption, considering the average δ15N values of between 19.2±2.1‰ and 18.5±1.7‰ for modern northern Chilean marine fish and sea mammals, respectively (Tieszen and Chapman 1992). In addition, there is the aforementioned archaeozoological evidence for marine resources in the cemetery at Pica 8 (Núñez Reference Núñez1984). Other factors, however, could affect the nitrogen isotopic composition of the humans. First, it is well known that dry desert conditions can significantly affect the nitrogen isotopic composition of soils, plants and animals (Evans and Ehleringer Reference Evans and Ehleringer1994; López et al. Reference López, Cartajena and Núñez2013; Díaz et al. Reference Díaz, Frugone, Gutiérrez and Latorre2016). It has been observed that modern and archaeological Atacama Desert plants can reach δ15N values up to 12‰ (Evans and Ehleringer Reference Evans and Ehleringer1994; Díaz et al. Reference Díaz, Frugone, Gutiérrez and Latorre2016). A similar situation has been observed for archaeological camelids at Tulan, south of the Atacama Salt Lake, also exhibiting relatively high δ15N values above 10‰ (López et al. Reference López, Cartajena and Núñez2013). Thus, human values of approximately 15‰ could be explained by the consumption of plant or animal resources with high δ15N values, and not necessarily a marine diet. However, there remains a substantial gap between this and the highest observed values of >20‰ that cannot be accounted for solely by aridity effects, so that another explanation is required.
Agriculture was fully developed and consolidated in the south-central Andes by the LIP. Early European chronicles document the use of fertilizers for agricultural activities in the Atacama Desert oases after the Conquest [Frezier Reference Frezier1717; Cieza de León Reference Cieza de León1922 (1553); Diez de San Miguel 1964 (1567)]. Fertilizers included llama dung, seabird guano and anchovy heads (Parsons and Psutty 1975; Julien 1985). Thus, it might be expected that the Pica 8 community fertilized their crops, particularly given the otherwise impoverished soils of the Atacama. The use of any of these manures would have undoubtedly elevated soil 15N composition, in turn reflected in crops and hence in the humans and animals consuming them. In particular, the impact of the use of seabird guano on crops has been clearly demonstrated by the experimental studies carried out by Szpak et al. (Reference Szpak, Millaire, White and Longstaffe2012a, Reference Szpak, Longstaffe, Millaire and White2012b; Reference Szpak, Longstaffe, Millaire and White2014). Thus, the consumption of crops fertilized with seabird guano and/or fish would increase human δ15N values significantly. Since the crops would only take up nitrogen from the manure, seabird guano would leave plant 13C and 14C unaffected, effectively uncoupling the two isotopic systems.
Thus, the consumption of crops fertilized with seabird guano presents a viable alternative to marine foods in the explanation of high δ15N values above 20‰. The interpretation of elevated 13C in contexts with access to both C4 plants (maize) and marine foods is always problematic, and so taking this isotope into consideration does not appear to offer any solutions, except in the case of individuals who clearly partook of neither food. This problem, together with the enormous variability in 15N introduced by the combination of marine foods and manuring with seabird guano, could explain the lack of any predictive relationships between stable isotope measurements and 14C offsets.
The positive correlation (R2 >0.5) observed for the textile isotope measurements and the offsets in radiocarbon years could be explained by the impact of fertilized crop consumption by these camelids. As mentioned above, the archaeological evidence from the sectors at Pica 8 suggested that some individuals/social units had access to and/or connections with the coast, which could have included the importation of seabird guano for crops. It follows that the camelids owned by these groups would have access to the fertilized, 15N-enriched crops, specifically maize, thus leading to the relatively high δ15N and δ13C values observed for some of the textiles. Humans would not show this correlation since their marine δ13C and δ15N values would overlap with those obtained through consuming the fertilized crops, along with some marine foods that were sufficient to impart a moderate offset in 14C years.
The results obtained here contrast with a case reported by Cases et al. (Reference Cases, Rees, Pimentel, Labarca and Leiva2008) for northern Chile, in which a considerable offset was observed between an ancient human body and its textile. This individual was found isolated near the María Elena Nitrate Mine in the Atacama Desert. 14C dating of the main textile and a repair patch gave very similar dates (1890±40 BP and 1870±40 BP, respectively). However, the muscle tissue from the individual yielded a 14C determination of 2390±70 BP, 640 yr older than the textiles. This individual also had high δ13Ccoll and δ15N values, –13.7‰ and 21.6‰ respectively, comparable to the highest values at Pica 8. In this case the marine diet suggested by the individual’s δ13C and δ15N composition, and by the direct association with dry fish and fishhooks (Cases et al. Reference Cases, Rees, Pimentel, Labarca and Leiva2008), is confirmed by the observed large 14C offset.
The contrasts observed between this particular case and the Pica 8 paired dates could be explained by temporal and dietary differences. Since this individual dates to the Formative Period, a significant dietary contribution of maize is unlikely as agriculture was still experimental and crops did not form a dietary staple at this time (Santana et al. Reference Santana, Herrera and Uribe2012; García et al. 2014; Santana-Sagredo et al. Reference Santana-Sagredo, Uribe, Herrera, Retamal and Flores2015b). Therefore, the use of seabird guano as fertilizer would not yet have featured, so that the high δ15N value seen in the María Elena individual was likely to be exclusively the result of a marine diet. It is also important to consider that ∆R values can change dramatically through time (Ascough et al. Reference Ascough, Cook, Dugmore, Barber, Higney and Scott2004, Reference Ascough, Cook, Church, Dunbar, Einarsson, McGovern, Dugmore, Perdikaris, Hastie, Frioriksson and Gestsdottir2010; Dewar et al. Reference Dewar, Reimer, Sealy and Woodborne2012; Hutchinson et al. Reference Hutchinson, James, Reimer, Bornhold and Clague2004; Ortlieb et al. Reference Ortlieb2011; Latorre et al. Reference Latorre, De Pol-Hoz, Carter and Santoro2015). For instance, the range of ∆R values reported for northern Chile during the Holocene goes from 48±36 yr to 1052±47 yr (Ortlieb et al. Reference Ortlieb2011). Thus, it is possible that ∆R values in the Formative Period were considerably higher compared to the LIP, which could also have an impact on the differences observed.
Diet Diveristy in Pica 8: Diachronic or Synchronic?
The paired dates obtained here strongly suggest that the isotopic variability observed at Pica 8 is not related to diachronic change. On the contrary, the limited range of dates for all but one individual, of between AD 990 and 1270 yr, suggests that considerable heterogeneity existed within a broadly contemporaneous community. This is further emphasized by the absence of any chronological trends between the stable isotope results and the calibrated dates. However, the lack of a predictive relationship between the stable isotope values and the small marine reservoir offset that was identified for a subset of individuals requires a re-consideration of the source of the isotopic variability at Pica 8. Rather than marine foods as previously proposed (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a), it may relate to differential access to seabird guano for manuring of crops, particularly maize, which by the LIP had become an important staple (Santana-Sagredo et al. Reference Santana-Sagredo, Uribe, Herrera, Retamal and Flores2015b). Thus, a link with the coast is still indicated, and no doubt included the transportation of some marine foods, which after all were present in the cemetery and also indicated in the diets of some individuals by the modest 14C offsets. But more importantly for explaining the isotopic variability—most notably the highly enriched 15N—is the access to seabird guano that these exchange links may have facilitated. What is most interesting, then, is that the resulting fertilized crops appear not to have been made available to the entire community at Pica 8. This reinforces the impression obtained from the differing isotopic results in the sectors, that sections of the community maintained distinct and separate diets and lifeways (perhaps kin-based?).
While significant temporal trends can be effectively ruled out by the results of the dating program, it is still possible that there were small-scale fluctuations in access to coastal resources (including seabird guano) over a period of some decades—too brief a duration to be detected by 14C. In this scenario, when guano was available it was used, significantly raising the δ15N levels for that generation. But it is hard to see how such fluctuations in access should come about, as the 90-km distance to the coast could hardly be construed as a barrier, particularly given the presence of pack animals and of long-established coast-inland trade connections. So, while acknowledging this possibility, we do not consider it likely.
The textiles at Pica 8 also hint at different origins and connections. For instance, some individuals (B0414, B0415, B0444, and B0460) were interred with textile styles typically associated with the local Pica-Tarapacá Complex (Agüero Reference Agüero2015). However, in this case they cross-cut the different cemetery sectors (B, D, and F). The hair bundle associated with individual B0447 is a common trait found on the Tarapacá coast. This individual was buried in sector I where most of the marine evidence was found, providing a δ15N value of 21‰, and an offset of 120 14C yr in the paired dating. Yet, since other individuals with equally high δ15N values show no offset, this in itself cannot be interpreted as indicating access to marine foods.
One male individual (B0492) from sector D was associated with a “non-local” bag of the type used for camelids to carry and store objects and food (known as costal) (Cases et al. Reference Cases, Lemp and Campos2007). The textile style has also been observed by one of the authors (CA) in the San Pedro Oases, specifically in Quitor and Solcor (Agüero Reference Agüero2004). However, it is not possible to associate it directly with the San Pedro de Atacama groups, since it could also have an origin in the highlands or elsewhere. Strikingly, this individual shows carbon and oxygen isotopic values that are clearly non-local, and are not inconsistent with highland origins, marking him as a clear outlier in Pica 8 (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a).
The most peculiar case is individual B0451 with the hairball found inside the mouth. This was an ancestral practice dating to the Formative Period in the Tarapacá coast and indeed the date date for the hairball (AD 796–969) is consistent with the Formative Period. One possible explanation is that it was removed from a burial in the coast and brought inland to the Pica oases. An alternative would be long-term curation of the hairball and re-use during the LIP. In any case, in Tarapacá it is associated with a typical coastal practice.
Individual B0420 has a relatively low δ15N value of 10.9‰ and is also an outlier for carbon and oxygen isotopes (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). At cal AD 1271–1383, this is the most “recent” of the dated individuals. However, the camelid textile associated with this individual has an offset of 110 yr (cal AD 1301–1414) compared to the human bone collagen, suggesting the consumption of approximately 15% marine protein by the human, but then this seems incommensurate with the δ15N value. Unfortunately, this textile (possibly a bag) in particular was in a poor state of preservation and it not possible to identify its type, style, and cultural tradition.
The only dates that could suggest temporal differences in the cemetery are individuals B0483, B0424, and B0431 for which we have no paired textile dates. Individual B0483 (cal AD 682–876) clearly falls in the Late Formative Period for the Tarapacá region, while individuals B0424 (cal AD 778–981) and B0431 (cal AD 788–987) present dates with probability distributions straddling the Late Formative Period to LIP boundary. It is possible that both are affected to a small extent by the MRE. A high marine diet was previously suggested for individual B0424 given the high δ13Ccoll and δ15N values of –14.1‰ and 24.4‰, respectively, together with a δ13Capatite value of –10.5‰ sufficiently low to suggest minimal maize in the diet (Santana-Sagredo et al. Reference Santana-Sagredo, Lee-Thorp, Schulting and Uribe2015a). However, given the results of the paired dating program, the inference of a high marine diet is now potentially problematic. Nevertheless, the fact that half of the individuals in the paired dating exercise exhibited offsets of approximately 100 14C yr makes a moderate contribution of marine foods a relatively uncontroversial claim.
As can be seen, the cultural variability of Pica 8 is reflected in the dietary composition of the population together with the textile styles associated with the burials. The dates clearly demonstrate that the isotopic and dietary heterogeneity is not diachronic in origin, supporting instead the presence of a multicultural community at Pica 8 during the LIP, maintaining distinct practices over some centuries at least. Yet these individuals, who consumed different diets and were interred with diverse textile types and styles, were buried in the same cemetery. Thus, identity and ethnicity factors appear to have played a major role in Pica during the LIP, leading to the coexistence (and presumably cohabitation) of different cultural groups.
CONCLUSIONS
Paired human–textile 14C dating from 9 graves at Pica 8 were undertaken as a first step to understanding the chronology of the Pica-Tarapacá Cultural Complex and the potential impact of the MRE on the dating of human remains from this region and period. The results show two clear groups: one with an approximately 100 14C-yr offset and the other with no offset. The former is consistent with a contribution of approximately 12–19% of marine protein, not insignificant but considerably less than was expected. Moreover, no correlations were observed between the high δ15N values—or indeed with δ13C measurements on collagen and apatite—and the 14C offsets between the paired dates. These results question the use of stable nitrogen isotopes in order to evaluate the MRE on the 14C dates for the LIP in northern Chile, and potentially more widely (e.g. southern Peru) and for other periods (e.g. Inca). Use of marine fertilizers and the possible effects of aridity could strongly influence the nitrogen isotopic composition of crops, animals and ultimately humans. Since fertilizers would not significantly affect the carbon composition of the crops, no impact of the MRE would be observed on them nor on the individuals consuming them. This, then, is a possible explanation for those individuals with high δ15N values and no 14C offset. The MRE for northern Chile and southern Peru remains poorly known and this situation greatly complicates the dating and interpretation of samples associated with the coast. We predict that the effect found in this paper will be largely restricted to the LIP and later periods when maize agriculture had become important and intensive, but of limited importance in the preceding Formative period.
The main question posed in the paper was whether the previously noted high isotopic/dietary diversity was related to temporal change, or whether it reflected variation within a living community. The results appear to favor the latter explanation, with no relationship being found between the stable isotope measurements and radiocarbon determinations. This in turn suggests the importance of sociocultural influences such as identity, ethnicity, and inequality in structuring foodways, and in determining differential access to marine resources, both in the form of food but also in the availability of seabird guano to fertilize the crops that were intensively cultivated at the Pica oasis. The alternative explanation that access to seabird guano was intermittent on short timescales not amenable to 14C dating and therefore undetectable seems improbable, since the advantages of the system once established should have been readily apparent. It does, however, raise the question of why not everyone in the community had access to fertilized crops. This situation requires us to rethink social complexity in the different Andean territories before the Inca expansion. Thus, while answering one question, we have raised others—not an unusual outcome in archaeology or indeed any science.
Acknowledgments
We thank the NERC Radiocarbon Facility for funding support (Grant number: NR/ 2015/1/7). We are also thankful to the FONDECYT project 1130279 and Anillo SOC1405. We are grateful to Tom Higham for commenting on the manuscript and to David Chivall for his help in the laboratory. We also thank Lorena Sanhueza and Nicole Barreaux for providing access to the textile collections at the Universidad de Chile. We are grateful to Sebastián Santana Sagredo and Marcelo Figueroa for editing some of the figures in the paper. We would like to thank Viviana Rivas for supplying the textile photographs in Figure 3.