


INTRODUCTION
Increased collaboration between natural sciences, archeology, and humanities has been bridging conceptual and methodological gaps over the last four decades; this has led to the creation of geoarcheology (Rapp and Hill, 1998; Ghilardi and Desruelles, Reference Ghilardi and Desruelles2009). Geoarchaeology is the most effective means for revealing how past human populations dealt with the impact on ecosystems of interacting geological, climatic, and biological factors. A Paleolithic site discovered in Grosseto in Italy during the spring of 2012 illustrates the value of detailed geoarchaeological studies. The site is located at Poggetti Vecchi that is a few kilometers north of Grosseto, and is an open-air, stratified, Paleolithic site that was unexpectedly brought to light at a depth of around 2.5 m while digging for a new 160 m2 thermal swimming pool located northward of other preexisting Paleolithic sites. Emergency excavations in a 60 m2 portion of the pool led to the unearthing of human artifacts and vertebrate bones of various mammal species, mostly from the straight-tusked elephant Palaeloxodon antiquus. Researchers from the Ministry of Cultural Heritage, Activities, and Tourism conducted the stratigraphic excavation, carefully recording the position of all the finds in order to produce GIS maps of each level.
The Poggetti Vecchi assemblage of faunal remains and artifacts is radiometrically dated to the latter part of the middle Pleistocene, which indicates that the tools found were made by early Neanderthals. At transition from the middle to the late Pleistocene, the only evidence of human presence in Italy is the human skeleton from Altamura, which is dated to the Marine Isotope Stage (MIS) 6–7 boundary (Lari et al., Reference Lari, Di Vincenzo, Borsato, Ghirotto, Micheli, Balsamo and Collina2015). In fact, most of the Italian sites with evidence of human activity on bones of Paleoloxodon antiquus are assigned to the Lower Paleolithic, e.g., Notarchirico (Piperno and Tagliacozzo, Reference Piperno and Tagliacozzo2001), Castel di Guido (Boschian and Sacca, Reference Boschian and Sacca2014), Atella (Abruzzese et al., Reference Abruzzese, Aureli and Rocca2015), Ficoncella (Aureli et al., Reference Aureli, Contardi, Giaccio, Jich, Lemorini, Madonna and Magri2015), and La Polledrara (Santucci et al., Reference Santucci, Marano, Cerilli, Fiore, Lemorini, Palombo, Anzidei and Bulgarelli2015).
The Poggetti Vecchi site is particularly interesting from the geoarcheological perspective, because it provides snapshots of the local environment at a time when early Neanderthals occupied the area. The site is especially valuable because it demonstrates the considerable skills of Neanderthals to opportunistically reconfigure and vary their activities according to their circumstances and resource availability. The present study is a part of a wider, multidisciplinary research project, which aims to reconstruct the behavior and ways of life of the early Neanderthal populations in the region. In particular, this paper focuses on the environmental conditions under which middle Pleistocene humans lived in the area around Poggetti Vecchi.
GEOLOGICAL BACKGROUND AND ARCHEOLOGY OF THE SITE
Geomorphic setting
The Poggetti Vecchi site is located on the northeastern side of Grosseto’s plain in southern Tuscany, Italy (Fig. 1A). This wide, coastal lowland is surrounded by low elevations composed of Triassic–Neogene clastic and carbonate deposits. The lowland was a lake, known as Lacus Prilis, through Etruscan and Roman times, before it became a wetland reclaimed by the nineteenth century (Stea and Tenerini, Reference Stea and Tenerini1996). Its long-lasting lacustrine-palustrine system resulted in the sandy-muddy surface sediments that extend for a few hundred meters underground, forming the thick Quaternary coastal basin infill (Bravetti and Pranzini, Reference Bravetti and Pranzini1987; Censini and Costantini, Reference Censini and Costantini2002). Poggetti Vecchi stands at the foot of a hill some 11 m above the plain that has developed on Mesozoic carbonate bedrock, which is extensively exposed on the slopes leading to the town of Roselle to the east (Fig. 1B). The lowland is affected by diffuse karstification that results in catastrophic subsoil collapses; one of these sinkholes formed in January 1999 at Bottegone, about 2 km north of Poggetti Vecchi (Censini and Costantini, Reference Censini and Costantini2002). The area has also many thermal springs (Bencini et al., Reference Bencini, Duchi and Martini1977), which suggests a local geothermal anomaly. Small magmatic Neogene-Quaternary intrusions are widespread in southern Tuscany and generate a high geothermal gradient (Baldi et al., Reference Baldi, Bellani, Ceccarelli, Fiordelisi, Squarci and Taffi1995).

Figure 1 (color online) (A) Location of the study area and schematic geological map of the Grosseto plain and surrounding elevations. (B) Oblique aerial view of the Poggetti Vecchi site from Bing Map (https://www.bing.com/maps/?cc=it). (C) Detailed oblique view of the excavation area, with annotated location of cores S1 and S2 and logs A and B reported in Figure 2.
Data description
This section reports the results of the stratigraphic, paleontological, and dating analyses. For the methods applied, see Supplement Item 1.
Stratigraphy of the site
The east–west trending, 20-m-long swimming pool excavation (Fig. 1C and 2) exposed vertebrate bones and stone artifacts (Fig. 3A) embedded in a few meters of largely muddy sediments of the plain. Several hydrothermal springs were found at the bottom of the excavation. The abundant discharge of the springs had to be drained with water pumps to keep the sections clear and to recover the specimens. Seven lithostratigraphic units were encountered in the excavated section; they will be described here, starting from the lowest (Figs. 2 and 3).

Figure 2 (color online) Correlation of stratigraphic data along the cross section indicated in Figure 1C. Samples analysed for mollusk and ostracod contents and for radiometric dating are shown.

Figure 3 (A) Plan view of Unit 2 during the archaeological excavation showing the framework of bones, stones (most of which artifacts), and wooden rods (indicated by red arrows). (B) Part of the section excavated along the northern side of the site where log A was measured. (C) The section excavated on the southern side (log B); note two molars and bones of Palaeloxodon antiquus on top of Unit 4. Ten centimeters divisions for scale in all photographs. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Unit 1
Unit 1 consists of about 0.5 m of massive, dark grayish mudstone (Fig. 3B). The fossils present in this unit comprise abundant mollusks, ostracods, micro-vertebrates (mammals, amphibians, fishes), and plant remains, the latter including either unaltered (seeds, carophites, pollen grains) or charred elements. The fossils were found uniformly dispersed in the sediment matrix.
Unit 2
The lower boundary of this bone bed is marked by an erosional contact. The unit is from 20 to 40 cm thick and mostly yielded remains of Palaeoloxodon antiquus mixed with bone and stone artifacts, scattered lithic production waste, but also wooden rods, which are presumably tools (Fig. 3A). Unit 2 is starved of sediment from the marginal source areas, and the skeletal remains accumulated to form a condensed layer of fauna. The top of the bone bed is an irregular surface formed by the assemblage of large bones and wooden remains.
Unit 3
Unit 3 is a lithologically composite unit, up to 20 cm thick, including two distinct divisions (Fig. 2): a lower one, indicated as 3a, made up of dark grayish mudstone with gastropod shells dispersed at the bottom; and an upper portion, called 3b, which consists of a few centimeters of laterally discontinuous, whitish, carbonate mudstone.
Unit 4
Unit 4 lays on an erosive surface and consists of millimeter- to centimeter-sized, clast-supported, coated, carbonate particles known as pisoliths, with scattered gastropod shells (Fig. 3C). Pisoliths are typical particles that grow by accretion of biochemical precipitates in stagnant thermal water; they still form in the artificial pools that were built at Poggetti Vecchi. Remains of Palaeoloxodon antiquus associated with stone tools were found at the base of this unit. Sediment samples also yielded microfossils dominated by ostracod shells. These sediments form a lenticular body up to 30 cm thick, which tapers towards the northern end of the excavation.
Unit 5
An erosional surface separates this unit from the underlying Unit 4. Unit 5 includes pebble-sized, clast-supported conglomerates with an intervening sandy-muddy matrix, arranged in a lenticular bed up to 25 cm thick (Fig. 3B). Clasts are subangular to subrounded and mostly composed of micritic grayish limestone with subordinate pisoliths and gastropod shells. Sparse vertebrate remains and stone tools were also recovered from this unit.
Unit 6
Unit 6 is up to 0.9 m thick and is a massive, dark, grayish mudstone similar to Unit 1 (Fig. 3B and C). The unit contains abundant gastropod shells and plant remains, together with occasional stone implements at the base, associated with a small amount of wooden items, possibly tools.
Unit 7
Unit 7, up to 2 m thick, starts with an erosive base. This unit consists of two layers (Fig. 2): the lower one, called 7a, is 0.4 m of massive, whitish, calcareous mudstone (Fig. 3C), and contains the skull of a juvenile Palaeoloxodon antiquus; the upper one, 7b, consisting of a set of beds made up of massive/graded calcareous sandstone and mudstone, separated from one another by erosive surfaces (Fig. 3B). An anthropogenic deposit up to 2 m thick, containing remains from the Roman Era, covers Unit 7 (Fig. 2, 3B–C).
The stratigraphic framework was constructed by correlating the stratigraphic logs recorded in the southern and northern portions of the excavation, and two shallow cores, S1 and S2, that were drilled just outside the excavated area (Fig. 2). This revealed that the site used to be in a small valley extending from the Poggetti Vecchi hill down to the plain. The seven lithological units are enclosed within a section, between cores S1 and S2, which widens a few meters north–south from the excavated area. Cores S1 and S2 were retrieved at depths up to 10 m. The sediments in the upper parts of the cores correspond to the deposits of Units 7 and 6, whereas the clastic succession beneath is dominated, from the top downwards, by massive mudstone with subordinate sandstone, which abruptly passes into reddish mudstone. The latter overlies a calcareous breccia present at the bottom of the cores. Both successions accumulated before the formation of Units 1–7. Summing up, our site consists of deposits piled up in a small valley carved at the side of a wide lacustrine area, whose water table repeatedly fluctuated, and which was ultimately affected by the geothermal heat flux.
Radiometric dating
Unit 4 contains pisoliths and vertebrate bones suitable for radiometric dating (see Supplement Item 1: Numerical dating). Pisoliths yield a U-series age of 171±3 ka, and a left lower molar of Bos primigenius provided an ESR/U-series age of 170±13 ka (Table 1). Based on these ages, the Poggetti Vecchi succession accumulated during a period spanning the latest MIS 7 interglacial and the whole MIS 6 glacial.
Table 1. (A) Activity ratios and age of the pisolite carbonatic fraction from Unit 4. All errors reported as 1σ. Age calculated using the Isoplot/EX 3.0 program proposed in Ludwig (Reference Ludwig2003). (B) ESR/U dating of Auroch’s tooth. A k-value (Da α Internal (mGy/a)) of 0.13±0.02 was used following the procedure outlined in Grün and Katzenberger-Apel (Reference Grün and Katzenberger-Apel1994). The enamel thickness (Da β (μGy/a)) removed during preparation process was used to calculate the beta contribution. Cosmic dose (Da α (γ+cosm) (μGy/a)) obtained using the formulae of Prescott and Hutton (Reference Prescott and Hutton1994). Uncertainties on the ESR/U-series datings (ESR/U-series ages (US or AU) (ka)) were calculated using Monte-Carlo approach following Shao et al. (Reference Shao, Bahain, Dolo and Falguères2014).

Archeological aspects of the site and their consequence
The open-air, stratified Paleolithic site of Poggetti Vecchi includes several anthropic layers: the oldest one is Unit 2, which yielded the richest collection of archaeological items. The excavated portion of this unit has an area of approximately 60 m2. As explained above, Unit 2 developed upon the erosional surface that formed on top of Unit 1, when the lake level dropped. Animals (mostly elephants) and groups of hunter-gatherers visited the valley bottom during the short period that this unit represents. The presence of humans is demonstrated by the occurrence of numerous lithic tools and lithic manufacturing waste, obtained from siliceous fluvial pebbles or cobbles deriving from the erosion of various Apennine mountain geological formations. The most common raw materials used at the site are different types of chert, radiolarite, and quartzite. The stone tools are only broadly indicative of an early middle Paleolithic age. About 200 lithic objects (cores, flakes, and unworked raw material) and several waste materials (debris and fragments) have been recovered. Most of the lithic implements, mostly scrapers but also notches and choppers, had been retouched. Dozens of tiny flakes have also been found at the site: these are considered to be by-products of retouching that took place on site.
In virtually all the European middle Pleistocene sites where remains of Palaeoloxodon antiquus are associated with human artifacts, hunting or scavenging is normally suggested when just one individual elephant is present (e.g., Weber Reference Weber2000; Villa et al., Reference Villa, Soto, Santonja, Perez-Gonzalez, Mora, Parcerisas and Sesé2005; Wenban-Smith et al., Reference Wenban-Smith, Allen, Bates, Parfitt, Preece, Stewart, Turner and Whittaker2006; Yravedra et al., 2010; Aureli et al., Reference Aureli, Contardi, Giaccio, Jich, Lemorini, Madonna and Magri2015; Santucci et al., Reference Santucci, Marano, Cerilli, Fiore, Lemorini, Palombo, Anzidei and Bulgarelli2015). In contrast, when many elephants are involved, they are generally assumed to have died due to natural causes (Gaudzinski, Reference Gaudzinski1996; Villa et al., Reference Villa, Soto, Santonja, Perez-Gonzalez, Mora, Parcerisas and Sesé2005; Boschian and Sacca, Reference Boschian and Sacca2014; Santucci et al., Reference Santucci, Marano, Cerilli, Fiore, Lemorini, Palombo, Anzidei and Bulgarelli2015).
Unit 2 at Poggetti Vecchi is the only European site that is dated to the latter part of the middle Pleistocene (MIS 6–7 transition) that has yielded the remains of several individuals of Palaeoloxodon antiquus of different ontogenetic ages that were found together with stone, bone, and wooden artifacts, mostly concentrated in a relatively small area at the western end of the excavation. Some of the implements found were horizontally oriented and commingled with the elephant remains (Fig. 3A), whereas others were found under the bones. The wooden artifacts present unequivocal evidence of human modification, as in, for example, the removal of small lateral branches and the removal of bark, and are therefore of particular interest, as such finds are very rarely encountered in prehistoric deposits due to the low durability of this material (Schniewind, Reference Schniewind1990; Blanchette, Reference Blanchette2003; Schoch et al., Reference Schoch, Bigga, Böhner, Richter and Terberger2015, and references therein). All the wooden remains of Poggetti Vecchi were found in a poor state of preservation: their restoration and analysis are still under way.
In Unit 4 of the same site, on the other hand, in the small area (25 m2) that was excavated, stone implements were found associated with the tusk and other remains of Palaeoloxodon antiquus. This Unit is radiometrically dated to about 171 ka, the terminus ante quem for Unit 2. Based on this dating, the human evidence from the site can be confidently attributed to the early Neanderthal period.
Unit 5 also revealed stone implements and faunal remains. Yet, only part of the lithic industry is fresh; the remainder is polished and could, therefore, possibly derive from a secondary context.
In the case of Unit 6, the lithic industry found at the base of the unit was found scattered with faunal and wood remains. This suggests that the shores of the lake were visited at a time when the water levels were starting to rise. The recurrent association of vertebrate remains and human implements in different layers (Units 2-4-5-6) of the Poggetti Vecchi stratigraphic succession indicates that humans visited the area on a regular basis, and were hunting and exploiting large mammals, especially Palaeloxodon antiquus.
The microenvironment generated by the hot thermal springs at Poggetti Vecchi at a time when the climate was cooling on a global scale probably attracted both the local fauna and their human pursuers, and no doubt these circumstances helped create Poggetti Vecchi’s unusual and interesting co-occurrence of faunal remains and human implements. The site is undoubtedly important because it dates to the middle-late Pleistocene transition, which is very poorly studied in Europe owing to the lack of evidence found thus far. In Italy, following the recently revised dating of the remains from Saccopastore to around 250 ka (Marra et al., Reference Marra, Ceruleo, Jicha and Salari2015), the only prehistoric evidence from the time of the MIS 6–7 transition is the human skeleton from Altamura (Lari et al., Reference Lari, Di Vincenzo, Borsato, Ghirotto, Micheli, Balsamo and Collina2015). Sites from this period are also rare in other parts of Europe, probably because of the concurrent climate cooling (Sanchez Yustos and Diez Martin, Reference Sanchez Yustos and Diez Martin2015). In this context, therefore, the Poggetti Vecchi site offers a noteworthy chance of gaining new insights into early Neanderthal behavioral plasticity and food procurement strategies (cf., Binford, Reference Binford1987; Weber, Reference Weber2000; Surovell and Waguespack, Reference Surovell and Waguespack2008; Rabinovich et al., Reference Rabinovich, Ackermann, Aladjem, Barkai, Biton, Milevski, Solodenko and Marder2012; Yravedra et al., Reference Yravedra, Panera, Rubio-Jara, Manzano, Expósito, Pérez-González, Soto and López-Recio2014; Scott et al., Reference Scott, Bates, Bates, Conneller, Pope, Shaw and Smith2014).
Paleontological analyses
Mollusks
Overall, the samples analyzed for this study come from Units 1, 3, and 6. They have yielded rich and well-diversified assemblages of non-marine mollusks in a good state of preservation. A total of 29 freshwater and terrestrial gastropod species (17 aquatic prosobranchs and pulmonates plus 12 land pulmonates) and one bivalve have been identified. Moreover, a few undetermined species of land Hygromiidae were also recorded. In many cases, the species found at Poggetti Vecchi are still present today in Europe, Italy included. The complete list of species, together with the abundance matrix, is reported in Table 2. The aquatic gastropods dominate both in number of species (varying from 8 to 13) and specimens (up to 2781 in sample PV4), whereas the land gastropods are generally represented by lower species diversity (varying from 0 to 9) and especially by a lower number of specimens (up to 47 in sample PV9; Table 2). There is, therefore, a much higher number of aquatic than of terrestrial species in the excavated interval. The details of the molluscan assemblage from Units 1, 3, and 6 are given below. Starting from Unit 1 (PV1-2), the total number of shells ranges from 2317 to 2436, and species richness from 18 to 19. Pseudamnicola moussonii dominates among the aquatic species (ca. 83%), followed by Bithynia tentaculata (7–8%), and Bithynia sp. (3–4.5%; Fig. 4), whereas Melanopsis etrusca (1.5–2%), Bithynia leachii (1–1.5%), Belgrandia thermalis (<1%), Valvata piscinalis (<1%), Galba truncatula (<1%), Radix peregra (<1%), Stagnicola palustris (<1%), Planorbis moquini (<1%), P. planorbis (<1%), and Acroloxus lacustris (<1%) are poorly represented. Freshwater bivalves include a single shell of Pisidium sp., and terrestrial gastropods consist of occasional hygrophilous taxa (Oxyloma elegans and Vertigo antivertigo), and very rare woodland species (Vitrea subrimata and Daudebardia rufa), and openland species (Vertigo pygmaea and Vallonia pulchella).

Figure 4 Malacological diagram (percentages of species) from the Poggetti Vecchi archeological site.
Table 2. List of mollusk species and abundance matrix of samples collected from the different layers of the Poggetti Vecchi archaeological site. Ecological classes (EC): AF, Aquatic freshwater; TH, terrestrial hygrophilous; TW, terrestrial woodland; TO, terrestrial openland; TC, terrestrial catholic; TU, terrestrial unassigned.

The total number of shells and species richness (1052 and 15, respectively) plummeted in Subunit 3a (PV3). P. moussonii still dominates, but at a lower percentage (61%) than in the previous samples. B. thermalis becomes proportionally more abundant (25%) than other aquatic species. Land gastropods include only occasional specimens of Carychium tridentatum, V. subrimata, and V. pulchella.
In Subunit 3b, the number of shells ranges from up to 2797 (PV5 sample) to 698–866 (PV7-9 samples), whereas species richness varies from 10 to 13. P. moussonii strongly dominates with a high percentage in PV4 (89%), just as in the PV1-2 samples. In contrast, its frequency declines abruptly in samples PV5–6 (47%–53%). Among the other aquatic taxa, Planorbidae are the best represented (7.5%), with the subordinate occurrence of B. thermalis and A. lacustris. Land gastropods are very poorly represented occurring with only occasional specimens of V. antivertigo and Hygromiidae indet.
In Unit 6 (PV7-9 samples), the total number of shells ranges from 1357 to 2316, with species richness ranging from 15 to 20. P. moussonii continues to dominate, with very high percentages (82–89%), followed by P. moquini, which, in turn, shows an increasing upward trend within the unit (2.5–10.5%). The other aquatic taxa are found in much lower frequencies. The spring water species Islamia sp. was recorded (with only one specimen) for the first time in the succession in this unit. Terrestrial species show a higher shell abundance (25–47) and species richness (6–9) than in the other units. In particular, the open-landscape Pupilla muscorum, the catholic taxa Punctum pygmaeum and Euconulus fulvus, and the ecologically unassigned species, Discus sp. and Monacha sp., first appear in the succession here.
Summing up, the molluscan fauna recovered from the deposits exposed by the excavation, but also from cores S1-2, is characterized by the dominance of freshwater taxa (Fig. 5). The most common and abundant freshwater species, P. moussonii (from 30% to nearly 90% of abundance), colonizes not only spring water, or stream and river waters not far from the spring, but also rocky and sandy substrata (Giusti and Pezzoli, Reference Giusti and Pezzoli1980; Giusti et al., Reference Giusti, Manganelli and Schembri1995; Cianfanelli, Reference Cianfanelli2009). Members of Bithyniidae (B. leachii and B. tentaculata, and Bithynia sp.), Lymnaeidae (R. peregra and S. palustris), Planorbidae (P. moquini and P. planorbis), and Acroloxidae (A. lacustris), which are almost constantly present in the samples (Fig. 4), require slow moving or still, clear, shallow waters found in well-vegetated lakes and ponds (Ložek, Reference Ložek1964; Girod et al., Reference Girod, Bianchi and Mariani1980). Furthermore, two species typical of thermal spring waters and slow flowing waters, in this case, M. etrusca (Fig. 5) and B. thermalis, are frequent, with variable percentages in Units 1–4 and 6, as well as in core S2. These indicate the presence of thermal springs either nearby (Units 1 and 6) or at the site itself (Units 3 and 4). M. etrusca requires running water fed by thermal springs with a high mineral content, temperatures between 18° and 35°C, and an average pH of 6.8–7.2 (Bartolini et al., Reference Bartolini, Aquiloni, Lori and Cianfanelli2010). Population densities decrease moving away from the spring and with faster flowing water (Cianfanelli et al., 1991; Manganelli et al., Reference Manganelli, Bodon, Cianfanelli, Favilli and Giusti2000; Bartolini et al., Reference Bartolini, Aquiloni, Lori and Cianfanelli2010). M. etrusca, which is often associated with T. fluviatilis, B. tentaculata, P. moussonii, B. thermalis, S. palustris, P. planorbis, and A. lacustris (Giusti and Pezzoli, Reference Giusti and Pezzoli1980; Cianfanelli et al., Reference Cianfanelli, Talenti and Calcagno1991, Reference Cianfanelli, Bodon, Giusti and Manganelli2010; Manganelli et al., Reference Manganelli, Bodon, Cianfanelli, Favilli and Giusti2000; Bartolini et al., Reference Bartolini, Aquiloni, Lori and Cianfanelli2010), is endemic to Tuscany. Abundant populations of the species are still present today in 7 sites with thermal waters between Livorno and Grosseto. One of them is the Poggetti Vecchi spring. M. etrusca is currently an endangered species due to the overexploitation of thermal waters (Manganelli et al., Reference Manganelli, Bodon, Cianfanelli, Favilli and Giusti2000; Cianfanelli, Reference Cianfanelli2010). In addition, a single individual of the spring water genus Islamia (Islamia sp.) was recorded in PV7-9. The generally rare occurrence of land hygrophilous taxa, such as C. tridentatum, V. antivertigo, and the presence of one species of Succineidae (O. elegans), indicates restricted marshland surroundings. Instead, P. muscorum (Fig. 5) and V. pulchella in the PV7-9 samples denote an open landscapes such as dry grassland and exposed areas (Ložek, Reference Ložek1964; Kerney, Reference Kerney1999) in the later stages of the site.

Figure 5 Selected examples of mollusc species from the sediment samples collected at Poggetti Vecchi archaeological site. Aquatic species: (A) Melanopsis etrusca Brot; (B) Pseudamnicola moussonii (Calcara); (C) Heleobia stagnorum (Gmelin); (D) Planorbis planorbis (Linnaeus). Land species: (E) Pupilla muscorum (Linnaeus). Scale bar=1 mm.
Ostracods
All the ostracod valves are well preserved and, generally, each species includes both adults and juveniles. A population age structure of this type indicates an autochthonous thanatocoenosis typical of an environment with moderate energy (Boomer et al., Reference Boomer, Horne and Slipper2003). Thirteen species of ostracods from eleven genera have been identified in the nine analyzed samples (Fig. 6, Table 3). These include: Darwinula stevensoni (Brady and Robertson), Vestalenula cylindrica (Straub), Candona (Candona) improvisa Ostermeyer, Candona (Neglecandona) neglecta Sars, Pseudocandona marchica (Hartwig), Cypria ophtalmica (Jurine), Cyclocypris ovum (Jurine), Ilyocypris monstrifica (Norman), Ilyocypris getica Masi, Heterocypris salina (Brady), Scottia pseudobrowniana Kempf, Paralimnocythere messanai Martens, and Cyprideis torosa (Jones).

Figure 6 Scanning electron micrographs of fossil ostracod valves recovered in the sediments of Poggetti Vecchi archaeological site. (A) Candona (Candona) improvisa Ostermeyer. (B) Candona (Neglecandona) neglecta Sars. (C) Cyprideis torosa (Jones). (D) Darwinula stevensoni (Brady and Robertson). (E) Cypria ophtalmica (Jurine). (F) Cyclocypris ovum (Jurine). (G) Vestalenula cylindrica (Straub). (H) Ilyocypris getica Masi. (I) Ilyocypris monstrifica (Norman). (J) Pseudocandona marchica (Hartwig). (K) Scottia pseudobrowniana Kempf. (L) Paralimnocythere messanai Martens. (M) Heterocypris salina (Brady). White bar=0.1 mm.
Table 3 Percentage frequencies of the ostracod species recovered in the different units of the Poggetti Vecchi archaeological site.

Almost all samples (except PV3) are dominated by H. salina, which is accompanied by P. marchica (particularly abundant in the upper part of the succession), with subordinated amounts of C. torosa and C. (N.) neglecta. All the other species show very low frequencies and are found scattered throughout the succession, except for C. (C.) improvisa, which is the dominant species (77%) in the low diversity assemblage recovered in sample PV3.
Cluster analysis was used to produce the dendrogram as illustrated in Figure 7, with a correlation coefficient of 0.98. Two clusters, A and B, emerge at the very low similarity value of about 0.07. Cluster A includes all samples from the archeological excavation site, except one from Unit 3a; these samples are mainly dominated by H. salina and include species typical of permanent shallow waters of ponds and lakes (Meisch, Reference Meisch2000; Furhmann, 2012). The presence of C. torosa with both noded and smooth shells, combined with the predominance of H. salina, indicate slightly brackish waters, with salinity values below the osmoregulation threshold of C. torosa (8 psu; Keyser, Reference Keyser2005; Frenzel et al., Reference Frenzel, Schulze and Pint2012). The co-occurrence of P. marchica restricts the likely salinity range to a maximum of 4 psu (Griffiths and Holmes, Reference Griffiths and Holmes2000). Such a low salinity is testified also by the abundance of Chara gyrogonites. Cluster B, on the other hand, includes only sample PV3: it is dominated by C. (C.) improvisa, a species typical of seasonal pools (Meisch, Reference Meisch2000).

Figure 7 Dendrogram resulting from cluster analysis in Q-mode using the UPGMA method and the Morisita–Horn distance applied to the samples from the Poggetti Vecchi archeological site.
The Mutual Ostracod Temperature Range (MOTR) method was applied to the assemblages of cluster A in Figure 1 to estimate the paleotemperature ranges at the time of the deposition of the sediment. Horne (Reference Horne2007) and Horne et al. (Reference Horne, Curry and Mesquita-Joanes2012) created this method for determining paleotemperature ranges by combining the Nonmarine Ostracod Distribution in Europe Database (NODE; Horne et al., Reference Horne, Baltanas and Paris1998) and WorldClim (version 1.3; Hijmans et al., Reference Hijmans, Cameron, Parra, Jones and Jarvis2005) datasets. The inferred paleotemperatures were then compared with today’s mean July and January temperatures at Poggetti Vecchi. The results of the MOTR analysis are reported in Figure 8, and they show that July and January paleotemperatures of 15–22°C for July and and −1–4°C for January were both significantly below the present day mean values, which are 25°C and 7.6°C, respectively (these being the average values for the period 2005–2014; data retrieved from the hydrological station Grosseto -Servizio Idrologico Regionale Toscana).

Figure 8 (color online) Estimated average air palaeotemperatures during the deposition of the sediments in the Poggetti Vecchi archaeological site based on the recovered fossil ostracods. Black lines indicate the tolerance temperature ranges for each species. The pink and light blue fields indicate the estimated July and January average temperature ranges in which the recoverd ostracod species could cohexist. The red arrows indicate the present July and January average temperatures for the Poggetti Vecchi area. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Vertebrates
Unit 2 was particularly rich in vertebrate remains, whereas several isolated bones were found in Units 4 and 6. A total of 634 specimens were retrieved from Unit 2: 149 (23.5%) of them are undetermined. Table 4 and Figure 9 report the list of the species, with their relative frequencies and abundances expressed in Number of Identified Specimens (NISP) counts.
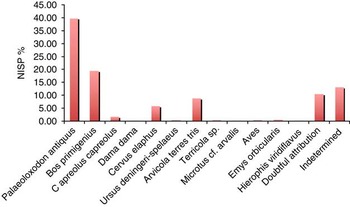
Figure 9 (color online) Relative frequencies of the vertebrate taxa of Poggetti Vecchi, expressed in Number of Identified Specimens (NISP) counts (data reported in Table 4).
Table 4. Vertebrate taxa from Poggetti Vecchi, with relative frequencies expressed in Number of Identified Specimens (NISP) counts.

The remains of very large-sized grazers, i.e., the straight-tusked elephant (Palaeloxodon antiquus) and the aurochs (Bos primigenius), predominate in the excavation. Based on NISP 67% and 32.6% were found for the two species, respectively. Browsing herbivores, such as the red deer (Cervus elaphus), roe deer (Capreolus capreolus), and fallow deer (Dama dama) are much rarer. Cervus elaphus, for example, is the only middle-sized species present in the Unit 2 assemblage; whereas the remains of roe deer and fallow deer total 1.4% and 1.2% of the bones of the small-sized species, respectively. A very advanced Ursus deningeri (transitional to U. spelaeus; Auguste, Reference Auguste1992; García et al., Reference García, Arsuaga and Torres1997; Pacher and Stuart, Reference Pacher and Stuart2008) is attested to by a right upper canine and a left m2. The mammal remains were accompanied by some scanty remains of the European pond turtle (Emys orbicularis), together with a vertebra of the European whip snake (Hierophis viridiflavus), and two fragmental bones of birds.
Only three specimens, in this case the lateral portion of diaphysis of a left humerus of Bos primigenius and the basal portion of a shed right antler and a fragmental diaphysis of a tibia of Cervus elaphus, show evidence of human modification. No such evidence has been observed on the elephant remains.
The terms of micromammals in Unit 2 (Table 4), these are dominated in NISP counts by the watervole Arvicola amphibius. This species, which is characterized by individuals with undifferentiated dental enamel (sensu Heinrich, Reference Heinrich1978), is typical of waterside habitats. This water vole totals approximately 95% of the small mammal counts. The genus Microtus, on the other hand, is represented in Unit 2 by only three specimens: a fragmental left m1, attributed to Microtus cf. arvalis, a left m1 of Microtus (Terricola) ex gr. savii, and a right M1 ascribed to Microtus sp. The first lower molars found show a relatively short and wide anteroconid as well as a wide anterior cap, which are features suggesting a Microtus (Terricola) species of the savii group.
A cross examination of Units 4 and Unit 6 shows that they yielded fossil assemblages very similar to that found in Unit 2. Indeed, the relative frequencies in Unit 4 are virtually the same as those observed in Unit 2. In Unit 6, however, the frequency of Arvicola amphibius drops to 88%. Microtus spp. also grow progressively less abundant from Unit 6 to Unit 2. Although the cheek teeth (M3 and m1) diagnostic of Microtus cf. arvalis are missing in the arvicolid assemblages from Units 6 and 4, we cannot exclude its undetected presence in either unit. Worth noting is the occurrence of a fragmental mandible of Crocidura cf. suaveolens and a fragmental maxillary bone of Oryctolagus sp. in Unit 6.
In terms of large mammals, based on the evidence of the skull, mandible, molar, and tusk remains from Unit 2, Palaeoloxodon antiquus is represented by four juveniles in the range of 1–8 years of age, together with a 14–15 year-old subadult, and two 25–30 year-old adults. Cervus elaphus is also represented by at least four individuals, ranging from 3 to 12/13 years of age. Particularly striking is the presence of at least nine left and three right shed antlers of this species. Also, remains of a single adult individual of Capreolus capreolus were found, alongside those of a fetus, a juvenile, and an adult of Bos primigenius. All the other taxa include one individual each.
Pollen
All the samples contained a fairly rich amount of well-preserved pollen grains, yielding a total of more than 100 taxa (Fig. 10). In all of the samples, herbaceous plants (NAP) dominate, whereas arboreal, shrubby and climbing plants (AP) never exceed 22%, indicating the persistence of an open environment throughout the whole time period under examination.

Figure 10 Pollen diagram of samples collected on the wall excavation of log A: selected taxa. Exaggeration 15×.
Noteworthy, among the arboreal taxa is the presence of Tertiary elements in the lowermost samples of Unit 1, such as Tsuga, cf. Keteleeria, and Carya. Two different morphotypes (i.e., with different exine thickness) of Ulmaceae suggest the possible presence of Zelkova in addition to Ulmus. These morphotypes occur throughout the sequence but always in low amounts. Buxus is always present in low concentrations throughout the succession. Its presence is unusual because Buxus grains are easily degraded.
Poaceae, on the other hand, are dominant and represented by different morphotypes, and this indicates the presence of extensive grasslands on the plain. The most frequent morphotype (A) is quite small, ranging from 22 to 28 µm, with a thick annulus measuring about 5 µm in diameter.
A noteworthy variety of pollen grains of hygrophytes and hydrophytes were recorded, indicating habitats with high biodiversity. They include Poaceae t. Glyceria type, Cyperaceae, Juncaceae, Alismataceae, Lythrum, cf Butomus, Callitriche, Potamogeton, cf. Sparganium t., Sagittaria, Typha spp., cf Lemna, Nymphaea, and Nuphar. Their amounts change throughout the stratigraphic sequence.
Wood remains
An isolated fragment of Ulmus sp. (elm) was recovered from Unit 1, at the base of the stratigraphic succession. Unit 2 contained several horizontally oriented wood fragments interspersed with the elephant remains. Most of the wood items were concentrated in a relatively small area of the excavation. A few wood fragments were also found at the base of Unit 6.
Buxus sempervirens L. (boxwood) is dominant throughout the succession, whereas decidous Quercus sp. (deciduous oak), Fraxinus sp. (ash), Juniperus sp. (juniper), and Populus/Salix (poplar/willow) are barely represented. The relative number of fragments for each of these trees is summarized in Table 5.
Table 5. Taxa identification of the wood fragments found in the succession of Poggetti Vecchi.

The Boxwood remains above all consist of straight branches, most of whose surfaces of most of these branches are partially or entirely blackened. Buxus is a small, slow-growing evergreen tree or shrub. It usually lives in forests of larger trees, associated with Juniperus. It can also be found next to xero-termophilus taxa, such as Quercus, in shrubs or thickets on well-drained calcareous substrates on plains to lower slopes. The high concentration of straight boxwood branches found in the Units 2 and 6 may indicate that they were intentionally selected by humans to make wooden implements. The study of these purported wooden tools is presently under way.
DISCUSSION
Paleoenvironmental reconstruction at the archeological site of Poggetti Vecchi
The integration of the multiproxy paleobiological data with the stratigraphy at the site has allowed for a detailed reconstruction of the landscape and an understanding of its evolution during the deposition of Units 1–6. The stratigraphic data together with the information provided by the fossil mollusk and ostracod assemblages indicate four different flooding events separated by regressions. During the flooding episodes, organic muds were deposited in a protected embayment of a shallow lake with rather homogeneous physicochemical characteristics. In contrast, concentrations of vertebrate remains and human artifacts accumulated on the erosive surfaces that formed during the regressions. The succession of the events is as follows.
A narrow valley was carved in the oldest deposits found in cores S1–S2, along the edge of a lake. A very shallow bay formed in this valley during the first episode of lake-level rise, and subsequently lacustrine sediments started to accumulate. The deposits of Unit 1 record the infrequent sedimentation of lacustrine mud on a poorly oxygenated bottom that favored the preservation of organic matter. Abundant and diversified mollusk and ostracod assemblages, dominated by P. moussonii and H. salina, respectively, inhabited the bottom and this indicates fresh or slightly saline waters (<4 psu). When the bay dried up an erosive surface formed, separating Unit 1 from Unit 2. As the lake receded, elephant carcasses and various human artifacts accumulated on its now-exposed shores.
Unit 3 records a second lacustrine phase. A new rise in the water table initially created a shallow, mud-dominated pool in the small embayment (Unit 3a); the predominance of the ostracod C. (C.) improvisa in this particular horizon may reflect seasonal fluctuations of the water level. This event was then followed by the deposition of carbonate mud (3b), which is the first significant evidence of thermal activity at the site. The pisoliths of Unit 4 document the formation, during the late middle Pleistocene (MIS 6), of a thermal pool similar to the man-made ones that were built in modern times. This MIS 6 thermal pool was visited by both humans and large mammals, which included Palaeoloxodon. Although the invertebrate assemblages were still dominated by P. moussonii and H. salina at this point, B. thermalis and M. etrusca reach their highest frequency in Unit 3, confirming the thermal influence on the waterbody.
The second lacustrine phase ended with a new lake-level fall, and the older deposits were dissected by fluvial downcutting (Unit 5). In fact, a short creek flowing down from the Poggetti Vecchi hill drained the valley. In this stage, runoff flow over the valley floor was of sufficient energy to transport the calcareous pebbles that formed the channel lag of Unit 5.
A new flooding (third phase) once again restored the lake embayment, as attested to by the deposits of Unit 6. In this case, the depositional conditions were virtually identical to those when Unit 1 was formed.
Yet another drop in lake-level fall led to a new, deep incision of the valley floor. The erosive surface separating Unit 7 from older units remained exposed for a while, as indicated by the occurrence of the isolated skull of Palaeoloxodon, as well as the abundant presence of terrestrial gastropods.
Finally in the fourth phase, the valley floor was once again carved and infilled with calcareous mud (Subunit 7a). This recreated the lake bay, but this time most likely under thermal influence. The valley was infilled with calcareous sand and subordinate mud (Subunit 7b), these last reworked from thermal carbonates by the water flowing toward the lake. The carbonate composition of Unit 7 indirectly indicates that the thermal activity had finally stabilized.
Pollen, terrestrial mollusks, and vertebrate remains provide further information about the overall environmental conditions surrounding of the site. At the time when the fossiliferous site was formed, the Poggetti Vecchi area was dominated by extensive open grassland; Palaeoloxodon antiquus and Bos primigenius grazed the prairie, while the red deer (Cervus elaphus) and the roe deer (Capreolus capreolus) browsed in sparse groves. The abundant frequency of freshwater fens, together with a high variety of wetland plants, denotes the presence of seeps and water bodies. Given the flux of wetland vegetation throughout the sedimentary succession, the bodies of water must have repeatedly expanded and contracted. Trees likely grew on the hills but also in well-drained areas of the plain; the predominance of Buxus, in particular, indicates well-drained, calcareous soils. It is often associated with travertines.
Chronological and paleoclimatic implications
The paleontological information reported above, but also the radiometric dating, indicates that the Poggetti Vecchi site developed during the late Quaternary, a period characterized by frequent climate fluctuations. Although infrequent, the presence of Tertiary pollen grains in the lowermost samples of Unit 1 is striking because these plants were thought to have disappeared from central Italy during the early-middle Pleistocene. Specifically, Tsuga and Carya had a marked decline in northern and Central Italy during the early middle Pleistocene, disappearing between 0.75 and 0.45 Ma, although they may have survived longer in Southern Italy (Corrado and Magri, Reference Corrado and Magri2011; Orain et al., Reference Orain, Lebreton, Ermolli, Combourieu-Nebout and Sémah2013). Zelkova rapidly reduced in cold-arid periods and expanded again in warm-moist periods and finally became extinct in peninsular Italy before the latest glacial maximum (Follieri et al., Reference Follieri, Magri and Sadori1986). Based on these data, the lower part of Unit 1 could not be younger than 0.4 Ma. We cannot exclude the possibility that the Tertiary pollen grains are actually allochthonous elements that have been reworked from older stratigraphic levels and that they may have migrated into higher beds via the capillary rise of mud.
The numerical ages of 171±3 ka and 170±13 ka obtained, respectively, from the pisoliths and Auroch’s tooth found in Unit 4 locate the studied succession within an interval bracketing the transition between the latest MIS 7 and the whole of MIS 6. The physical stratigraphy of the transgressive and regressive deposits that formed during the time interval indicated by the age determination of Unit 4 suggests the following tentative calibration (Fig. 11).

Figure 11 (color online) Tentative correlation of the Poggetti Vecchi Stratigraphy, calibrated for Unit 4 with radiometric datings to the Late Quaternary Marine Isotope Stages.
Unit 1 transgression: lake highstand during the warm substage MIS 7a or during the following interstadial at the transition to MIS 6 (between 200–191 ka). Under either of these hypotheses, the Tertiary flora recorded in the lower part of Unit 1 would have survived in this area more than 0.4 Ma.
Unit 2 regression: lake lowstand and subaerial erosion at the onset of MIS 6.
Units 3 and 4: lake transgression and activation of the thermal system in the first interstadial of MIS 6 (around 170 ka).
Unit 5 regression: lake lowstand at the successive stadial.
Unit 6 transgression: lake highstand during the second interstadial.
Unit 7 regression: lake lowstand at the acme of MIS6 (around 130 ka). Deposition of Unit 7 may have occurred during the climatic rise in temperatures culminating with the peak interglacial interval MIS5e.
The paleobiological inferences based upon the Poggetti Vecchi small mammal remains provide further information supporting the tentative synchronization of the succession with the main climatic events of the associated time period. The dominant Arvicola amphibius is a typical inhabitant of moist environments. The presence of Microtus cf. arvalis in Unit 2, the decreasing abundances of M. (Terricola) ex gr. savii and the absence of Crocidura and Oryctolagus in the following units, might indicate cooler conditions moving upwards in the succession. Modern Italian populations of Microtus cf. arvalis are confined to the northern regions of the peninsula and are absent from the central Apennine further south. Therefore, its presence in southern Tuscany at very low altitudes is indicative of colder conditions at the time of Unit 2 than those existing today. In contrast the occurrence of Crocidura cf. suaveolens, Oryctolagus sp., and Microtus (Terricola) savii in Unit 6 is indicative of warmer habitats.
The large mammal assemblage and the herpetofauna from Unit 2 including thermophilic taxa, such as the roe deer Capreolus capreolus, the European pond turtle Emys orbicularis, and the European whip snake Hierophis viridiflavus, show a wide distribution ranging from the warm peri-Mediterranean area, all the way into the much cooler northern European contexts (Lelièvre et al., Reference Lelièvre, Blouin-Demers, Bonnet and Lourdais2010a, Reference Lelièvre, Le Hénanff, Blouin-Demers, Naulleau and Lourdais2010b). The European pond turtles are usually only active above 5°C and so are another indicator of temperatures across the region (Novotný et al., Reference Novotný, Danko and Havaš2004). Palaeoloxodon antiquus is typical of temperate contexts but also occurred in cooler conditions (Mazza et al., Reference Mazza, Martini, Sala, Magi, Colombini, Giachi, Landucci, Lemorini, Modugno and Ribechini2006).
On the whole, the fauna of Unit 2 comprises of species with varied climatic distributions that seem to be well-tuned with the fluctuations that occurred within the relatively short period between the onset of MIS 6 (lake regression and erosion at the top of Unit1), and the next interstadial (lake transgression and deposition of Units 3 and 4). Intense seasonal oscillations can reasonably be expected to have occurred during the time span recorded by the Poggetti Vecchi succession. The occurrence of Microtus cf. arvalis is not at odds with the hypothesis that perhaps a cold snap occurred during the time of the deposition of Unit 2, which might have caused the sudden death of the elephant clan.
The molluscan fauna discloses relevant information that may assist in defining the climate regime during the deposition. The recorded presence of open-landscape species, such as P. muscorum and V. pulchella from Unit 6, indicates that this Unit formed under cool climatic conditions (Rousseau, Reference Rousseau1989; D’Amico et al., Reference D’Amico, Esu and Magnatti2014). These conditions are compatible with the global cooling trend that culminated in the glacial maximum of MIS 6.
The ostracod fauna that was collected from Units 1–6 and analyzed with the MOTR method suggest there were mean annual temperatures about 6°C lower than today, which is in line with the temperatures at the onset of MIS 6 based on estimates from the Vostok Ice core in Antarctica, which indicates 7.5°C lower than today (Petit et al., Reference Petit, Jouzel, Raynaud, Barkov, Barnola, Basile and Bender1999). In light of Ayalon et al.’s (Reference Ayalon, Bar-Matthews and Kaufman2002) study, the humid period dated at 171 ka, which is also the calculated age of Unit 4, was likely the first interstadial of the glacial interval MIS 6.
CONCLUSIONS
The radiometric dating of about 171 ka obtained from the pisoliths and third lower molar of Bos primigenius of Unit 4 places the Poggetti Vecchi archaeo-paleontological site in a very crucial time span. This time period tells us that early Neanderthal populations produced the implements. Archaeo–paleontological sites attesting to the middle to late Pleistocene transition are scarcely documented in Europe. Poggetti Vecchi therefore offers the chance of obtaining an unprecedented amount of information on the behavior of early Neanderthals under the circumstances that have been documented through from this study.
Our analysis shows that at the time when the bone accumulation was formed the area consisted of widespread, open, wet grassland, scattered with water bodies, woods, and, perhaps, riverine belts of trees. The hot thermal springs in one of these water bodies located in a small embayment, probably created an attractive recess for the resident fauna, especially in a time period when the climate was globally deteriorating. It would appear from the artifacts found that the area could have been routinely patrolled by early Neanderthal hunter-gatherers who were monitoring the movements of the fauna that tended habitually to congregate in the warmer valley of Poggetti Vecchi. We can speculate that during a cold snap, large amounts of game animals would gather in the recess. Many of them were possibly debilitated by the harsh, inhospitable circumstances, or most probably already lay dead, as perhaps happened in the case of the elephants. Their concentrated presence represented a valuable resource that could have attracted the local human pursuers. If this reconstruction is correct, the collected evidence indicates that these early Neanderthals were particularly skilled at modifying their survival strategies to take advantage of the particular environmental circumstances and opportunities presented to them in a changing environment.
ACKNOWLEDGMENTS
We are particularly indebted to the owner of the site A. Ceccarelli, who funded the excavation. We wish to thank A. Pessina and G. Poggesi for their support during the completion of this study, S. Pozzi, S. Caloni and S. Caramiello for the restoration of the fossil bones. We are very grateful to M.-H. Moncel, D. Bridgland, and C. Marean for significantly improving the manuscript. We thank K. Eadie and J. A. Thonn for the language editing. This study was financially supported by the Italian Ministry of Cultural Heritage, the “Fondazione Banca Federico del Vecchio” and the “Ente Cassa di Risparmio di Firenze”. Additional funding for the research was provided by Italian MIUR (the Italian Ministry of Education, University and Research) ex60% grants to the Department of Earth Sciences and to the Department of Biology, both of the University of Florence, and by Italian Mibact (Ministry of Cultural Heritage and Activities and Tourism).
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/qua.2017.51


















