INTRODUCTION
Since the Late Miocene, African rainforests have exhibited large variations in their extent (Maley, Reference Maley1996). Several investigations have shown that, beginning from 10 Ma, African rainforests regressed considerably and fragmented during major arid periods (Maley, Reference Maley2010), such as during the last glacial maximum (LGM) between ca. 24,000 and 16,000 cal yr BP (Dupont et al., Reference Dupont, Jahns, Marret and Ning2000; Dalibard et al., Reference Dalibard, Popescu, Maley, Baudin, Melinte-Dobrinescu, Pittet, Marsset, Dennielou, Droz and Suc2014). However, biogeographic and genetic research has demonstrated that forests survived in several refugia (Maley, Reference Maley1996; Hardy et al., Reference Hardy, Born, Budde, Daïnou, Dauby, Duminil and Eben2013; Duminil et al., Reference Duminil, Mona, Mardulyn, Doumenge, Walmacq, Doucet and Hardy2015) during these arid periods. After the LGM arid period, a major phase of forest expansion began ca. 12,000 cal yr BP, particularly with its northern fringe reaching ca. 7°N on the Adamaoua Plateau (Vincens et al., Reference Vincens, Buchet and Servant2010; Fig. 1, site 12). Moreover, during the late Holocene, two major climatic crises impacted the Central African forest block. The first one, around 4000 cal yr BP, caused the contraction of the forest domain in favor of savanna expansion at its northern and southern peripheries. The second one, around 2500 cal yr BP, markedly perturbed the central core of the forest, leading to forest fragmentation and an associated major erosional phase. The main goal of this paper is to clearly describe these two major climatic crises, particularly the second phase that led to forest fragmentation. Using available data, a new schematic map is presented to describe the fragmentation of the rainforest between ca. 2500 and 2000 cal yr BP from the Gulf of Guinea to the Rift Mountains (Fig. 1). Finally, an assessment is made of how this fragmentation phase could provide a model for projected global warming-related environmental changes in Central Africa.
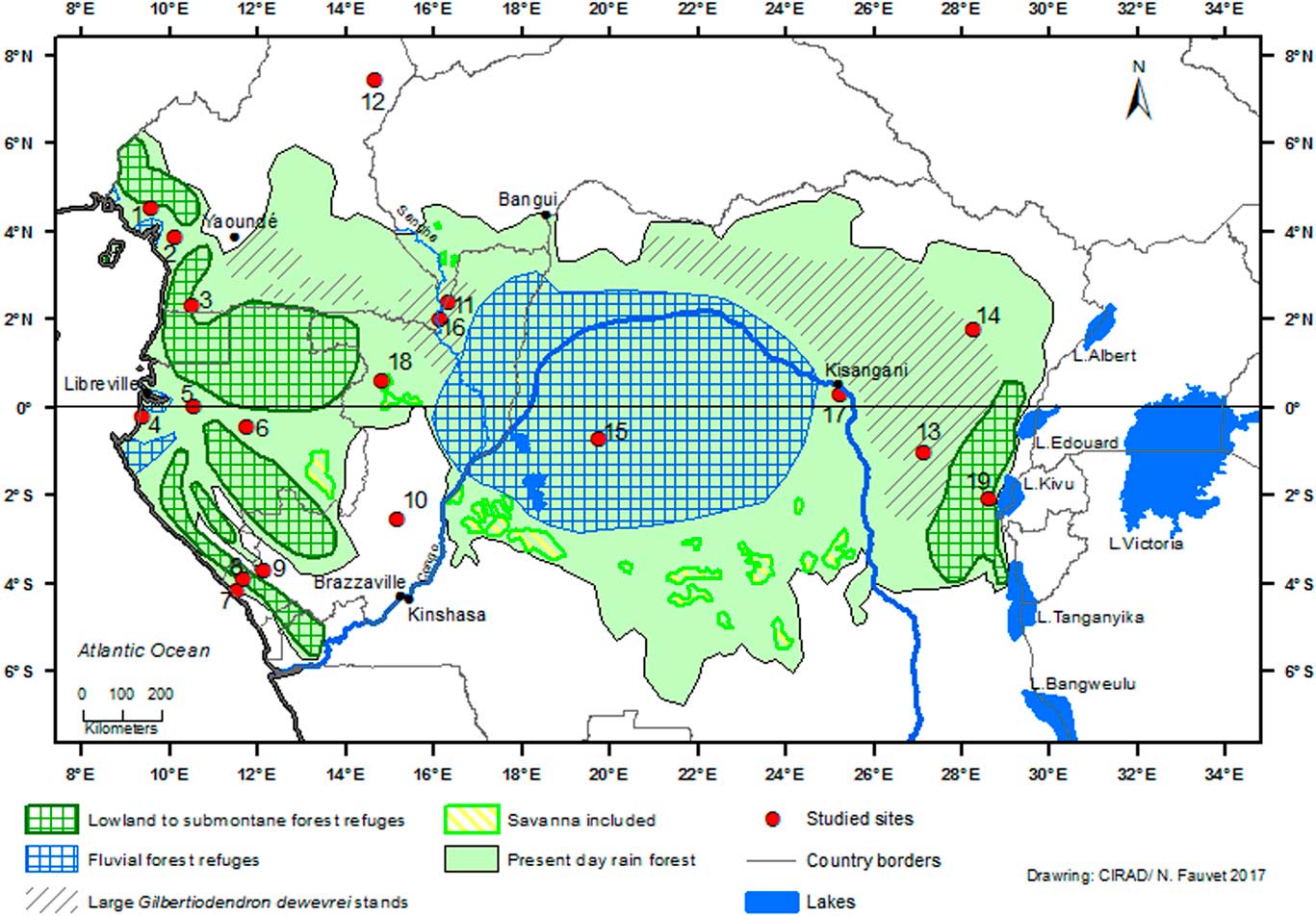
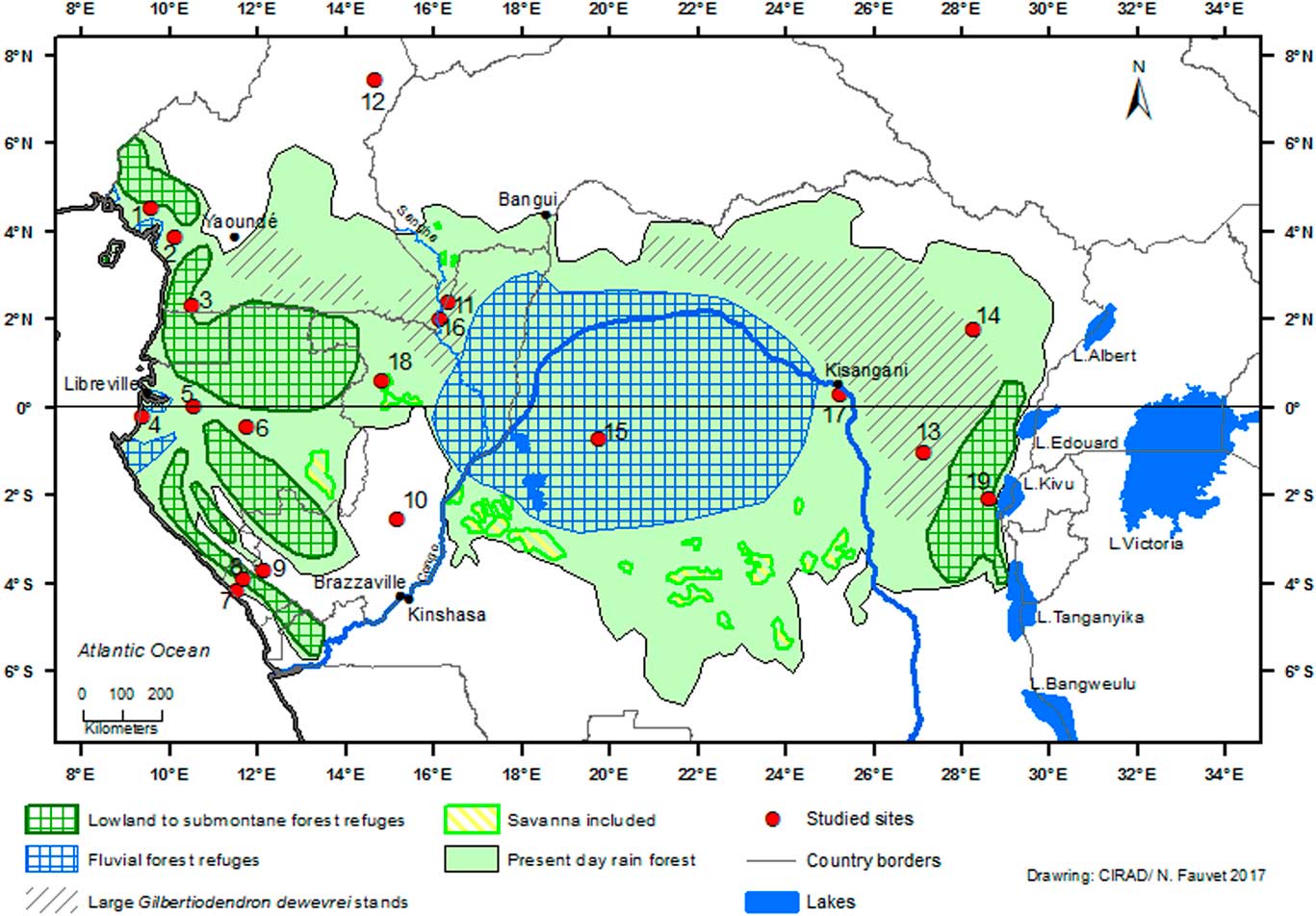
Figure 1 (color online) Schematic map of rainforest refugia across Central Africa ca. 2500–2000 cal. yr BP in the Equatorial Forest domain, and present-day area of monodominant forest with large Gilbertiodendron dewevrei stands. The basis of this map presents the current status of Central African biotopes (Mayaux et al., Reference Mayaux, Richards and Janodet1999; partly adapted from Maley, Reference Maley2001, Reference Maley2002). Sites: (1) Barombi-Mbo. (2) Ossa. (3) Nyabessan. (4) Maridor. (5) Nguène. (6) Lopé. (7) Kakamoéka. (8) Kitina. (9) Sinnda. (10) Ngamakala. (11) Mopo. (12) Balang. (13) Osokari. (14) Ituri. (15) Imbonga. (16) Kabo. (17) Yoko. (18) Odzala-Kokoua. (19) Kahuzi-Biega.
GENERAL SETTING
Diverse paleoenvironmental data, mainly sedimentological and palynological, are available from sites across Central Africa: southern Cameroon, Gabon and western Congo, as well as several parts of the Congo Basin (in the Democratic Republic of Congo [DR Congo]), and into southern West Africa, Bénin, and Ghana.
The first climatic crisis was characterized by a major contraction of the Equatorial Central African forest domain, which occurred abruptly around 4000 cal yr BP, accompanied by a marked expansion of the adjacent northern and southern savannas. To the north, major savanna expansion occurred in Togo and Bénin (Guinean domain) with the opening of the “Dahomey Gap,” as indicated by the high representation of grasses in the pollen record from lake Sélé in Bénin (Salzmann and Hoelzman, Reference Salzmann and Hoelzmann2005). To the west in Ghana, the crater lake Bosumtwi experienced an important highstand, which began at ca. 11,000 cal yr BP and ended with a major regression around 4000 cal yr BP (Shanahan et al., Reference Shanahan, Overpeck, Wheeler, Beck, Pigati, Talbot, Scholz, Peck and King2006). At this time, synchronous with the opening of the “Dahomey Gap,” the savannas probably also extended in northern Ghana, although isotopic data (δ15N) shows that the rainforest persisted in the periphery of lake Bosumtwi (Talbot and Johannessen, Reference Talbot and Johannessen1992). Farther to the east, in southern Cameroon, at the same time, isotopic data (δ13C) from several soil profiles reveal that savanna expansion also took place south of the Adamaoua Plateau (ca. 7°N) until near to the Sanaga River (Desjardins et al., Reference Desjardins, Turcq, Nguetnkam, Achoundong, Mandeng-Yogo, Cetin and Lézine2013).
This phenomenon similarly occurred south of the equator, with the expansion of coastal savannas in Gabon (lake Maridor: Giresse et al., Reference Giresse, Makaya, Maley and Ngomanda2009), as well as in the Niari area (western Congo), where the drying out of lake Sinnda (3°50'S) around ca. 4000 cal yr BP is linked with savanna development (Vincens et al., Reference Vincens, Schwartz, Bertaux, Elenga and De Namur1998). Hubau et al. (Reference Hubau, Van den Bulcke, Kitin, Mees, Baert, Verschuren, Nsenga, Van Acker and Beeckman2013) studied charcoal from several soil profiles of the Luki forest reserve located at the southernmost edge of the Mayumbe Guinean forest (lat. 5°45'S; DR Congo) and surrounded by savanna. They concluded that “increasing aridity between 4000 and 2500 cal yr BP did not cause severe rainforest breakdown through fire” (Hubau et al., Reference Hubau, Van den Bulcke, Kitin, Mees, Baert, Verschuren, Nsenga, Van Acker and Beeckman2013, p. 337). Indeed, during this period pollen records from several sites in the central part of the forest domain, such as lakes Barombi Mbo and Ossa (Reynaud-Farrera et al., Reference Reynaud-Farrera, Maley and Wirrmann1996; Maley and Brenac, Reference Maley and Brenac1998; Giresse et al., Reference Giresse, Maley and Kossoni2005) and Nyabessam swamp, Cameroon (Ngomanda et al., Reference Ngomanda, Neumann, Schweitzer and Maley2009); lake Nguène, Gabon (Giresse et al., Reference Giresse, Makaya, Maley and Ngomanda2009); and lake Kitina, Congo, (Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004) show the persistence of evergreen forests, particularly comprising trees belonging to the families Fabaceae-Caesalpinioideae and Sapotaceae:
The second climatic crisis began around 2500 cal yr BP and continued until 2000 cal yr BP (Maley, Reference Maley2001, Reference Maley2002). As shown particularly by pollen analyses, this phase is characterized by climate change impacting the remaining forest domain, leading to a decrease of many mature forest taxa, accompanied by a rapid expansion of pioneer-type forest vegetation, and locally a brief savanna extension. The main result of this major perturbation was forest fragmentation. Curiously, however, this period is not characterized by an arid climate but greater seasonality in rainfall distribution. Indeed, annual rainfall overall remained rather high as indicated by large-scale erosional phenomena, and particularly the formation of stone-lines that resulted from abrupt lateral movements of coarse debris over large areas (Thiéblemont et al., Reference Thiéblemont, Flehoc, Ebang-Obiang, Rigollet, Prian and Prognon2013; Maley et al., Reference Maley, Giresse, Favier and Doumenge2014). The existence of large debris flows suggests important rainfall events of rather short duration. During this period, large-scale erosion also impacted the northern part of the tropical African strip as far as the Sahara (Maley, Reference Maley2010; Maley and Vernet, Reference Maley and Vernet2015).
EVIDENCE FOR THE 2500 – 2000 CAL YR BP CLIMATE CRISIS IN CENTRAL AFRICA
Dating of the event and pollen data
Regular lacustrine sedimentation during the period 2500 to 2000 cal yr BP at many coring sites has allowed the development of reliable radiocarbon chronologies for reconstructing precisely the vegetation history using pollen analysis. The following sites have provided important data: in southern Cameroon, at lake Barombi Mbo (Maley, Reference Maley1992; Maley and Brenac, Reference Maley and Brenac1998), Nyabessan swamp (Ngomanda et al., Reference Ngomanda, Neumann, Schweitzer and Maley2009; Neumann, et al., Reference Neumann, Bostoen, Höhn, Kahlheber and Ngomanda2012a), and at lake Ossa (Reynaud-Farrera et al., Reference Reynaud-Farrera, Maley and Wirrmann1996; Giresse et al., Reference Giresse, Maley and Kossoni2005). For the lake Ossa site, pollen analysis carried out on the 5.50 m core OW4 determined the level when abrupt paleoenvironmental change occurred. In conjunction with four radiocarbon dates obtained near this level, the beginning of the climatic crisis can be fixed at 2500±50 cal yr BP (Reynaud-Farrera et al., Reference Reynaud-Farrera, Maley and Wirrmann1996; Giresse et al., Reference Giresse, Maley and Kossoni2005). Similar results were obtained in Gabon, at lake Kitina (Elenga et al., Reference Elenga, Schwartz, Vincens, Bertaux, de Namur, Martin and Servant1996) and lake Nguène (Giresse et al., Reference Giresse, Makaya, Maley and Ngomanda2009).
Palynological and also biogeographic and genetic investigations have shown that the remaining dense humid forests were largely but not solely restricted to a number of major distinct refugia or “macro-refugia” at this time (Maley, Reference Maley1987, Reference Maley1996, Reference Maley2001, Reference Maley2002; Colyn et al., Reference Colyn, Gautier-Hion and Verheyen1991; Hardy et al., Reference Hardy, Born, Budde, Daïnou, Dauby, Duminil and Eben2013; Duminil et al., Reference Duminil, Mona, Mardulyn, Doumenge, Walmacq, Doucet and Hardy2015). These landscapes were characterized by dry open forests, with wooded savannas and dense rainforests “micro-refugia” (sensu Leal, Reference Leal2001) in favorable edaphic areas along rivers. Pollen analyses showed that, in some places, savanna expansion occurred briefly, such as in southwestern Cameroon around Barombi Mbo (Maley and Brenac, Reference Maley and Brenac1998). Indeed, remnants of the formerly extensive savanna exist close to this site today, just north of Mount Cameroon (fig. 2 in Maley and Brenac, Reference Maley and Brenac1998). However, Poaceae pollen in the Barombi Mbo record never exceded 40% of the total pollen count, and thus the landscape may have been a mosaic of more-or-less dense forests with some savanna encroachments. Phytoliths and isotopic data from south-eastern Cameroon and northern Congo showed that the crisis at ca. 2500 cal yr BP favored the development of pioneer and disturbed forest vegetation, but not a true general savanna phase (Bremond et al., Reference Bremond, Bodin, Bentaleb, Favier and Canal2017).
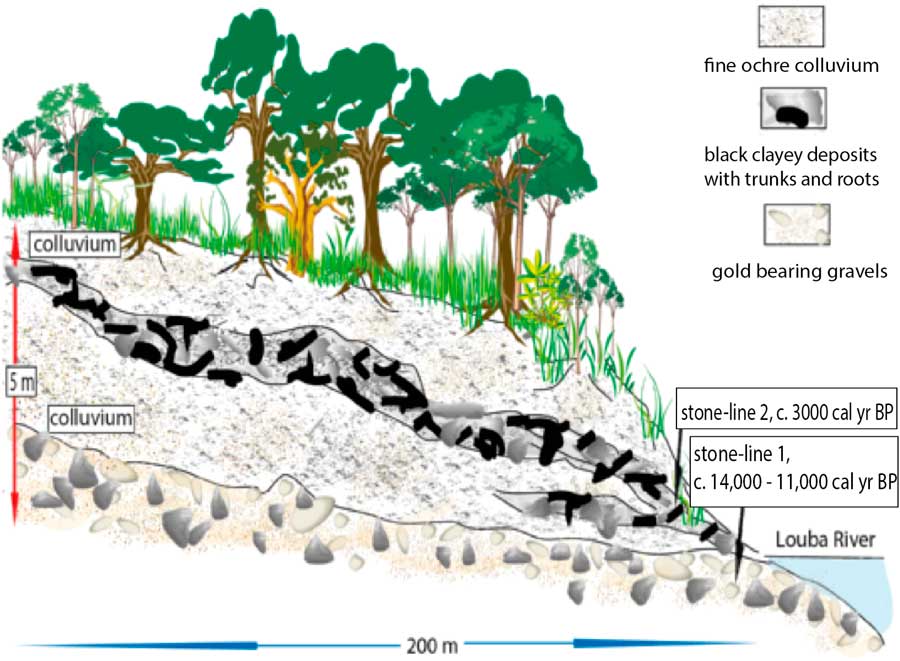
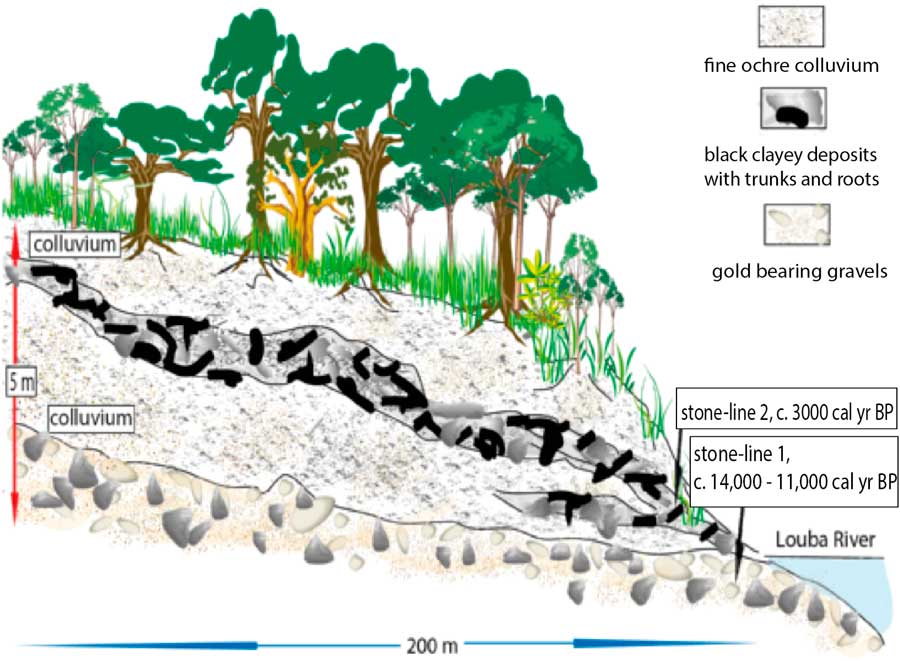
Figure 2 (color online) Kakamoeka soil profile from the Kouilou River catchment, Mayumbe (Fig.1, site 7; Maley and Giresse, Reference Maley and Giresse1998).
Nevertheless, some data showed that the forest became highly disturbed locally from about 4200 cal yr BP and particularly from 2500–2000 cal yr BP, as noted by an increase of δ13C at site F15 of Bremond et al. (Reference Bremond, Bodin, Bentaleb, Favier and Canal2017; Fig. 1, near site 18), under a present-day wooded savanna. Nowadays, isolated savannas persist in this region, as well as in northern Congo and southwestern Central African Republic (CAR; Fig. 1; see also Mayaux et al., Reference Mayaux, Richards and Janodet1999). Several studies have also indicated that the savannas of northern Congo, in Odzala-Kokoua National Park (Fig. 1, site 18), were formerly more extensive (Hecketsweiler et al., Reference Hecketsweiler, Doumenge and Mokoko Ikonga1991; Kouka, Reference Kouka2002). The transgression of forest vegetation upon savannas observed in that region is a general one, spanning from West Africa to Central Africa (Maley and Doumenge, Reference Doumenge2012), as reported also below.
Sedimentological characteristics
In many profiles dated to the period 2500–2000 cal yr BP, the sediments are coarse and linked to abrupt erosive phenomenon (Maley et al., Reference Maley, Giresse, Doumenge and Favier2012b, Reference Maley, Giresse, Favier and Doumenge2014). First, to the west in Ghana, as observed in the sediment of lake Bosumtwi, a major change occurred around 2650 cal yr BP, when the black and massive organic-rich mud deposited during the early and middle Holocene in a deep and well-mixed eutrophic lake system shifted to annual finely laminated sediments similar to today, resulting from a shallower, stratified meromictic system. The dominant control on this abrupt change is greater seasonality, directly caused by an enhanced summer monsoon resulting in increased erosion delivering terrigenous sediments to the lake (Shanahan et al., Reference Shanahan, Beck, Overpeck, McKay, Pigati, Peck, Scholz, Heil and King2012). In Gabon and Congo, these erosive phenomena were very large and resulted in the emplacement of a thick and widespread stone-line sequence commonly composed of huge blocks covering the slopes and accumulating in the valley bottoms (Thiéblemont et al., Reference Thiéblemont, Flehoc, Ebang-Obiang, Rigollet, Prian and Prognon2013; Maley et al., Reference Maley, Giresse, Favier and Doumenge2014).
In the hills of Mayumbe, western Congo, along a small tributary of the Kouilou River, a soil section at Kakamoéka (4°10' S–11°58' E; Fig. 1, site 7 and Fig. 2), 5 m thick, contains two stone-lines: one at its base and one near the middle of the profile that contains large broken pieces of trees in a silty clay. This detritic level, which is rather similar to the stone-line sequence described in Gabon, likely resulted from a catastrophic hillwash phase, pulling down a pre-existing forest that grew on the slope of the valley (Maley and Giresse, Reference Maley and Giresse1998). Within this upper stone-line, a wood sample has been dated to ca. 3000 cal yr BP. Given that the general erosive phase began around 2500 cal yr BP, it seems likely that the fossil tree from which the Kakamoéka radiocarbon date was obtained lived in the forest some time before the catastrophic gullying event. Nevertheless, interpretation of this age should be treated with caution as: (i) only one date is currently available, and (ii) there is a large (500 yr) gap between this date and the regionally recognized erosive phase with which it is correlated. However, this correlation is supported by new data obtained 50 km to the south, near Pointe-Noire (Congo) and close to Loango (4°39' S–11°48' N), where, along the mouth of a small coastal river draining from the Mayumbe, a profile exhibits a sequence with five successive floods events, with the oldest one dated ca. 2500 cal yr BP (Malounguila-Nganga, Reference Malounguila-Nganga, Giresse, Boussafir and Miyouna2017, fig. 2). Furthermore, in the southern part of the Mayumbe region (lat. 5°45' S; DR Congo) Hubau et al. (Reference Hubau, Van den Bulcke, Kitin, Mees, Baert, Verschuren, Nsenga, Van Acker and Beeckman2013) collected charcoal from several soil profiles, which all had dates younger than 2300 cal yr BP, including three related to the perturbed forest phase discussed here.
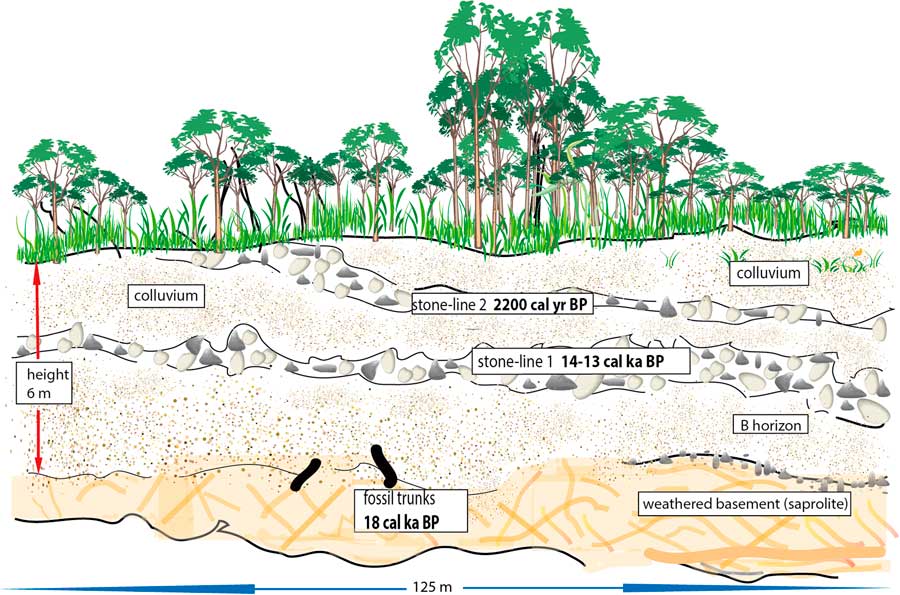
Moreover, a similar soil profile was described by Runge (1997, Reference Runge2001) and Runge et al., (2016) at Osokari (1°16' S - 27°49' E; Fig. 1, site 13) in the eastern part of the Congo Basin (DR Congo). This 6-m-thick section, cut along a new road, exhibited two phases of erosion and gullying linked to two stone-line sequences (Runge, Reference Runge1996, Reference Runge2001; Runge et al., Reference Runge, Sangen, Neumer, Eisenberg and Becker2014; Fig. 3). At the base of the profile, just above the substratum, the remains of two large trunks were dated to ca. 18,000 cal yr BP. As these dates correspond to the period of maximum rainforest fragmentation in central equatorial Africa during the LGM (Maley, Reference Maley1987, Reference Maley1996), these two fallen dead trees can be linked to this phase. In the Osokari soil profile, the lowest stone-line was deposited after the remains of these trunks and hence occurred towards the end of the Pleistocene, and very probably about ca. 14,500 – 13,000 cal yr BP, because a strong increase in the sediment accumulation rate of detritic sediments occurred in the Congo deep-sea fan at this time (Giresse et al., Reference Giresse, Bongo-Passi, Delibrias and Duplessy1982; Bayon et al., Reference Bayon, Dennielou, Etoubleau, Ponzevera, Toucanne and Bernell2012; Maley et al., Reference Maley2012a; see below). Later, during the Holocene, soil colluvium developed, but finally the upper part of this soil profile was cut by another erosional phase linked to a detritic level that was dated at ca. 2200 cal yr BP on charcoal contained in the sediments (Runge, Reference Runge1996, Reference Runge2001; Runge et al., Reference Runge, Sangen, Neumer, Eisenberg and Becker2014). This soil profile thus exhibits two detritic sequences, which can be linked to two phases of strong forest disturbance: the first at the end of the LGM, and the second one synchronous with the perturbation as described from the western part of Central Africa (see above). The main features of this profile can be compared to those of the Kakamoéka section as described from western Congo (Fig. 2) and suggest a similar paleoclimatic evolution from the Gulf of Guinea until the Rift of Central Africa.
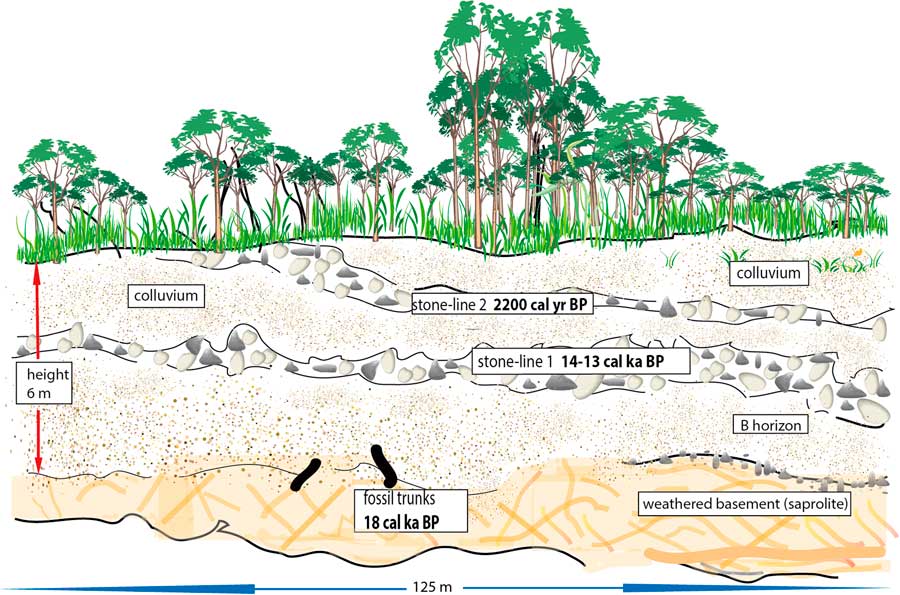
Figure 3 (color online) Osokari soil profile from the eastern part of the Congo Basin, DR Congo (Fig.1, site 13; Runge, 1997, 2013).
Other data confirm the very general character of the disturbed climate phase between 2500 and 2000 cal yr BP. The analysis of a core from the submarine delta of the Congo River revealed a major erosional phase dating from this interval and affecting the whole of the Congo River Basin, including its northern, southern, and eastern margins (Bayon et al., Reference Bayon, Dennielou, Etoubleau, Ponzevera, Toucanne and Bernell2012; Maley et al., Reference Maley, Giresse, Doumenge and Favier2012; Neumann et al., Reference Neumann, Bostoen, Höhn, Kahlheber and Ngomanda2012a; Fig. 4). The climatic disturbance that caused the erosional phase extended beyond the eastern edge of the Congo Basin as far as East Africa. Indeed, pollen and diatom records from sediments sampled in Lake Victoria showed that the water level was very low about 2200 cal yr BP and that this phenomenon was accompanied by an increase in savanna at the expense of regional forest environments (Stager et al., Reference Stager, Cumming and Meeker1997). These savanna variations have been further substantiated by recent pollen analyses in the Lake Kivu region of eastern DR Congo (Fig. 1, site 19; Kabonyi et al., Reference Kabonyi and Roche2015a, Reference Kabonyi, Roche and Gerrienne2015b).
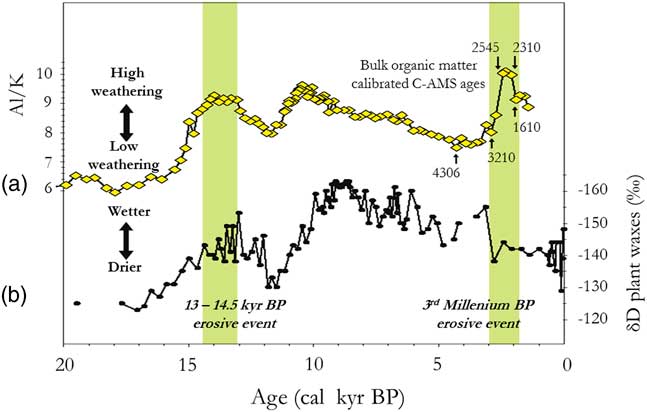
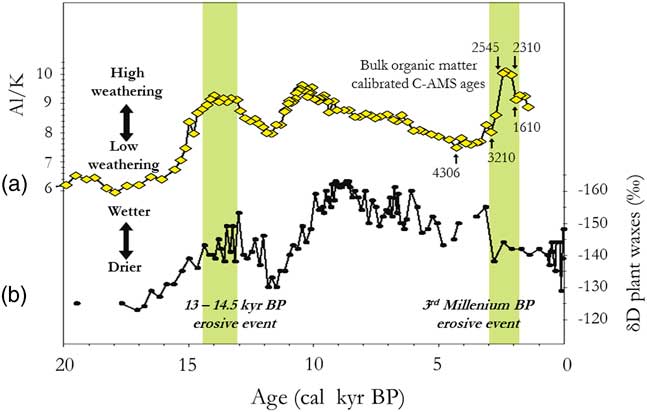
Figure 4 (color online) Palaeo-hydrologic and climatic variations in the Congo Basin for the last two stone-line formations (partly adapted from Bayon et al., Reference Bayon, Dennielou, Etoubleau, Ponzevera, Toucanne and Bernell2012, fig. 3). (a) Variations in the terrestrial plant-wax δ D values in core GeoB6518-1, as an index of precipitation changes in Central Africa (from Schefuß et al., Reference Schefuß, Schouten and Schneider2005). (b) Variations in detritic flux, mainly kaolinite, as an erosive proxy in the Congo River catchment, from Core KZAI-1 collected in the Congo deep-sea fan sediments (from fig. 3b in Bayon et al., Reference Bayon, Dennielou, Etoubleau, Ponzevera, Toucanne and Bernell2012, fig. 3b; cf. Maley et al., Reference Maley, Giresse, Doumenge and Favier2012).
The Sangha River Interval in the western part of the Congo Basin
Several biogeographical investigations, particularly in floristic studies, have allowed the identification of sub-centers of plant endemism in the Equatorial African rainforests (Letouzey, Reference Letouzey1968; White, Reference White1979) and the separation of two main floristic domains: (i) the Guinean to the west and (ii) the Congolian to the east. This research showed that the boundary between these two domains lies in the Sangha River Interval (SRI) area, in the northwest region of the Congo Basin extending south to the Batéké Plateaus (Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004). Some species, especially in the sub-family Fabaceae-Caesalpinioideae, are present in the Guinean and Congolian domains but totally absent or only marginally present in the SRI. The extension of opened habitats in the past, during relatively drier or more seasonal periods, is inferred from the presence of savannas (Fig. 1, site 18) and the existence of relict savanna species such as Phoenix reclinata (Arecaceae) in some isolated sites (Doumenge et al., Reference Doumenge, Gonmadje, Doucet and Maley2014; Bostoen et al., Reference Bostoen, Clist, Doumenge, Grollemund, Hombert, Muluwa and Maley2015). The current vegetation of the SRI area (Gillet, Reference Gillet2013) is mainly composed of a mosaic of forest formations, including evergreen dense forests with the monodominant tree Gilbertiodendron dewevrei, semi-deciduous dense humid forests characterized by Cannabaceae (Celtis spp.), and Malvaceae-Sterculioideae (Gond et al., Reference Gond, Fayolle, Pennec, Cornu, Mayaux, Camberlin, Doumenge, Fauvet and Gourlet-Fleury2013). The SRI area includes a particular open canopy forest formation with a low density of large trees but with the understory covered by giant herbaceous species, mainly Marantaceae and Zingiberaceae, including some higher than 5 m tall. The open canopy forest formation covers a number of large areas, including about 500,000 hectares in the Odzala-Kokoua National Park (Fig. 1, site 18; Hecketsweiler et al., Reference Hecketsweiler, Doumenge and Mokoko Ikonga1991) and farther east until west of the Sangha River. Similar open canopy forest with Marantaceae is also widespread farther north, in southeastern Cameroon (Letouzey, Reference Letouzey1968, Reference Letouzey1985).
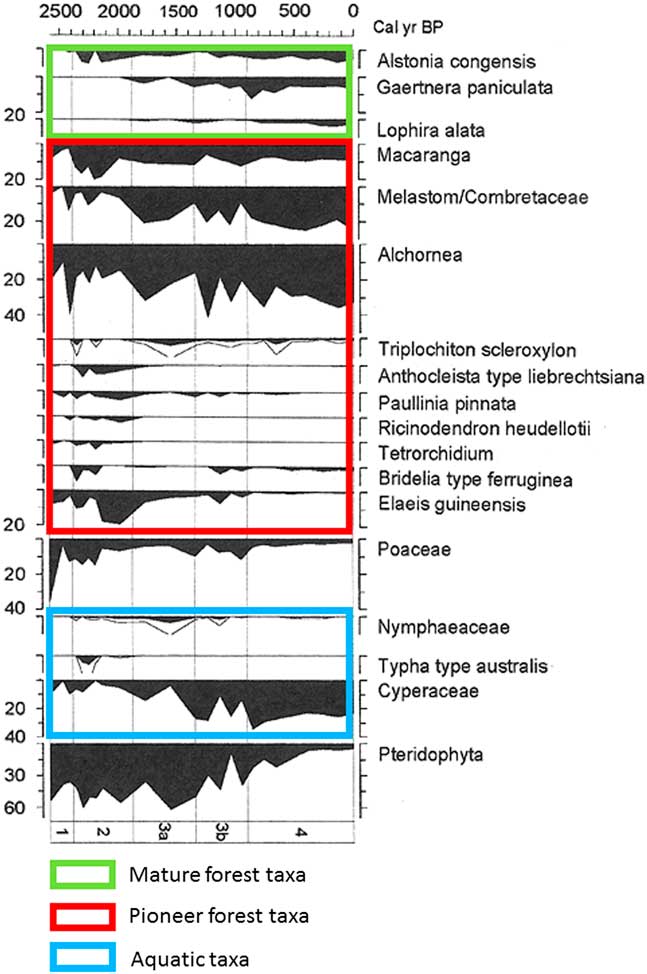
Brncic et al. (Reference Brncic, Willis, Harris, Telfer and Bailey2009) collected a 1-m-long core inside the SRI region, in the swampy area of Mopo Bai (in northern Congo; Fig. 1, site 11), east of the Sangha River. The chronology, based on several AMS radiocarbon and thermoluminescence dates, shows that the bottom of the core (dated ca. 2500 cal yr BP) corresponds to the beginning of a disturbed phase characterized by major erosion and detrital input. The pollen diagram obtained by Brncic et al. (Reference Brncic, Willis, Harris, Telfer and Bailey2009) was adapted by Maley and Willis (Reference Maley and Willis2010 and Fig. 5). Variations in the dominant pollen taxa show:
-
(1) A phase at the base, dated ca. 2500 cal yr BP, characterized by a marked development of Poaceae (36%, but not aquatic, as the swamps only developed later), which is probably related to a brief period of local savanna expansion.
-
(2) A notable subsequently rapid increase of several pioneer taxa, particularly 40% of Alchornea pollen and already 10% Elaeis guineensis (oil palm) pollen. Towards the end of this period, around ca. 2300 cal yr BP, Poaceae pollen declined to less than 13% and some shade-tolerant tree taxa, such as Celtis and Funtumia africana, began to increase.
-
(3) During the following phase extending until ca. 2000 cal yr BP, pioneer taxa, especially oil palm, continued to dominate, which increased to 20%. The importance of Pteridophyta spores during this period indicates heavily disturbed environments.
-
(4) During the next phase, beginning ca. 2000 cal yr BP, the present-day swampy environment appeared progressively, mainly characterized by a large increase in Cyperaceae pollen. Oil palm pollen remained abundant during this phase, until ca. 1000 cal yr BP. These data are confirmed by the numerous oil palm kernels found in the soils of the northern Congo and southern CAR and dated between 2400 to 900 cal yr BP (see below).
-
(5) Finally, after ca. 1000 cal yr BP, a large increase in rainforest diversity is observed, linked to the progressive spread of numerous more mature trees belonging to the present-day forest habitat (Alstonia congensis, Gaertnera paniculata, and Lophira alata), and accompanied by the continuous colonization of pioneer trees.
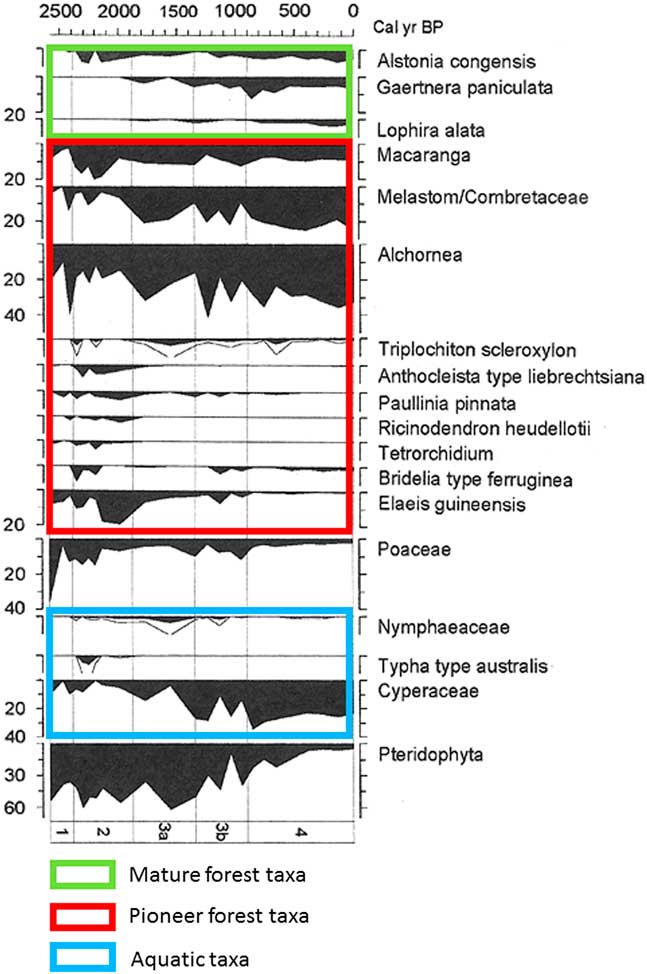
Figure 5 (color online) Mopo Bai pollen diagram in northern DR Congo (Fig.1, site 11; Maley and Willis, Reference Maley and Willis2010; adapted from Brnic et al., Reference Brncic, Willis, Harris, Telfer and Bailey2009). Percentages calculated from the total pollen count (left side). Pollen types (right side) plotted against age of the core (top), and arranged in ecological groups (mature forest taxa, pioneer forest taxa, and aquatic taxa). Lines next to filled diagrams indicate 3×exaggeration of pollen taxa.
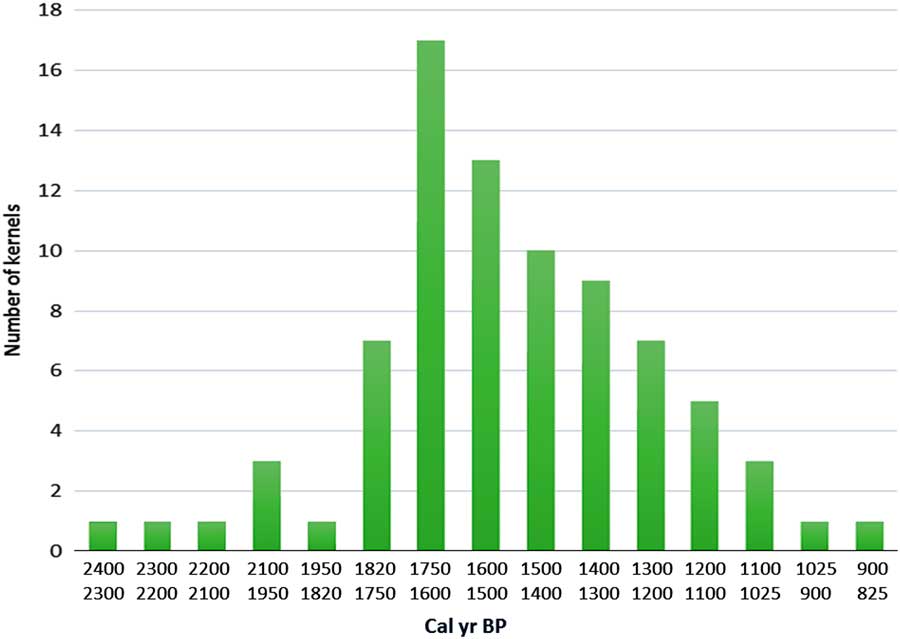
Given that Mopo Bai lies within the perimeter of the SRI interval, its pollen diagram demonstrates that the reemergence of this interval began by a short savanna phase induced by the ca. 2500 cal yr BP climate crisis. The present-day development of extensive open canopy forests through this interval is a legacy of the perturbation’s persistence (see next section). Many charcoal remains have been found in the soils of these open canopy forests, rather than under dense forests (Gillet, Reference Gillet2013; Morin-Rivat et al., Reference Morin-Rivat, Fayolle, Gillet, Bourland, Gourlet-Fleury, Oslisly and Bremond2014), and the numerous samples dated are from the period ca. 2300 to 1200 cal yr BP. Moreover, abundant oil palm kernels were collected in the soils of the Dzanga-Ndoki National Park, southernmost CAR, near Mopo Bai, and dated from 2400 to 900 cal yr BP (Fay, Reference Fay1993, Reference Fay1997; Maley and Willis, Reference Maley and Willis2010; Fig. 6). This phase of major oil palm development in southern CAR, similar to that observed in the Mopo Bai pollen record, northern Congo, also occurred in several sites from southern Cameroon: the pollen records from lake Barombi Mbo (Maley and Brenac, Reference Maley and Brenac1998) and lake Ossa (Giresse et al., Reference Giresse, Maley and Kossoni2005), as well as from the marine core (C61) taken offshore the Sanaga River mouth, which drains a large part of southwestern Cameroon (Maley, Reference Maley1999). Therefore, these data demonstrate that this widespread quasi-synchronous phase of oil palm development resulted from the expansion of Elaeis guineensis as a natural pioneer preceding the reinstatement of the mature rainforest, mainly on sandy alluvial soils formed after the 2500 cal yr BP erosive phase, and not linked with cultivation carried out by the Bantu people (see data and discussions in Maley, Reference Maley1999; Maley and Chepstow-Lusty, Reference Maley and Chepstow-Lusty2001; Maley and Doumenge, Reference Doumenge2012).
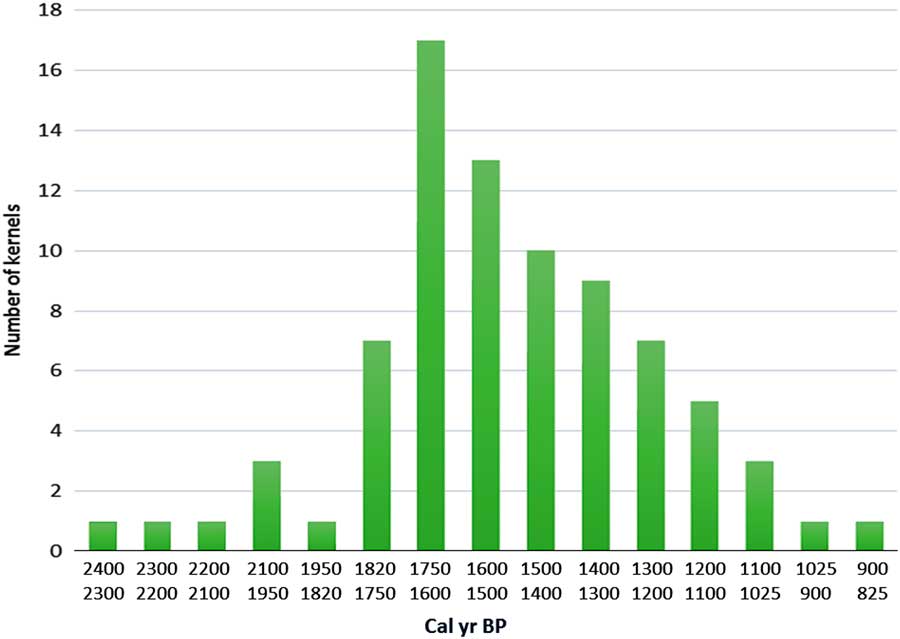
Figure 6 (color online) Variations of Elaeis guineensis kernels in southernmost CAR between ca. 2400 to 900 cal. yr BP. In the present day mature rain forest of the Dzanga-Ndoki National Park, southernmost CAR, the Biologist Fay (Reference Fay1993, Reference Fay1997) collected oil palm kernels in numerous soil profiles. 80 samples of these kernels were radiocarbon dated. The frequency diagram of the calibrated dates is presented by ca. 100 years intervals. The shape of this diagram is characteristic of a natural phase of progressive replacement of a pioneer vegetation by a more mature one (Maley and Willis, Reference Maley and Willis2010).
Furthermore, as shown by Bostoen et al. (Reference Bostoen, Clist, Doumenge, Grollemund, Hombert, Muluwa and Maley2015), the opening of the SRI following the ca. 2500 cal yr BP crisis constituted an important corridor for the rapid and large-scale spread of Bantu-speaking communities, who benefited from the major pioneering nature of the oil palm, which provided an important food source. From the SRI, the Bantu were subsequently able to reach the Inner Congo Basin, as recorded at the Imbonga archaeological site dated to ca. 2350–2050 cal yr BP (Fig. 1, site 15; Eggert, Reference Eggert1987; Wotzka, Reference Wotzka1995). The Imbonga site is defined as the earliest ceramic sequence in the western Congo Basin and probably represents the first signature of the early Bantu language dispersal (Bostoen et al., Reference Bostoen, Clist, Doumenge, Grollemund, Hombert, Muluwa and Maley2015).
The dynamics of the Gilbertiodendron dewevrei forest formation
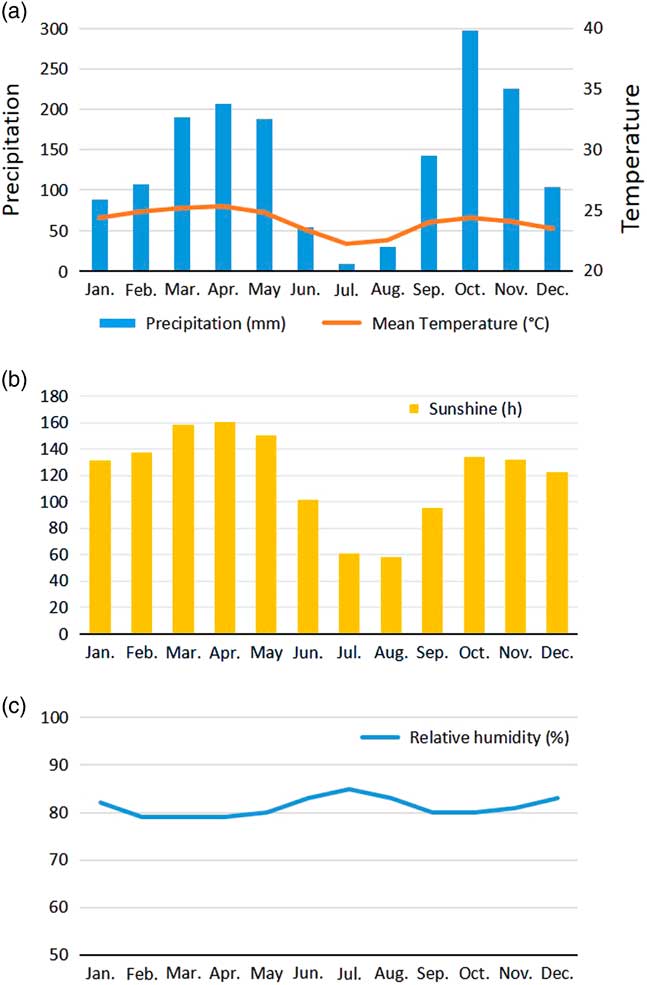
The large disturbance beginning ca. 2500 yr BP, which strongly disturbed and eliminated previous forests, provided space for pioneer vegetation and, later, to new forest formations (as shown in the Mopo Bai pollen record, Fig. 5), particularly those characterized by Gilbertiodendron dewevrei (“Limbali”), a shade-tolerant tree belonging to the Fabaceae-Caesalpinioideae. This canopy tree forms very dense, practically monospecific evergreen stands in which the species regenerates well under its own shade, while growth of young individuals can be favored by small openings in the canopy (Gérard, Reference Gérard1960; Torti et al., Reference Torti, Coley and Kursar2001; Doumenge, Reference Doumenge2012). It develops either on poor dryland soils or on soils where the water table is high or even inundated for short periods. Although the species is present in all Central Africa from the Atlantic coast to the Western rift, it forms vast monodominant stands only from southeastern Cameroon to the northeast and east of the Congo Basin (see distribution in Fig. 1; Louis and Fouarge, Reference Louis and Fouarge1949; Léonard, Reference Léonard1953; Letouzey, Reference Letouzey1985; Doumenge, Reference Doumenge2012), north of the so-called “climatic hinge” (near ca. 2°N latitude), which delimits areas under the climatic influence of the Northern Hemisphere from those of the Southern Hemisphere (Gonmadje et al., Reference Gonmadje, Doumenge, Sunderland, Balinga and Sonké2012). It is noteworthy that forests growing under the influence of the Northern Hemisphere climate experience a drier main dry season (December to February) than the one developing under the Southern Hemisphere climate (June to September). This southern climate (Fig. 7, Makokou example, Gabon), shows that the relative humidity is highest during the dry season, in contrast to northern climates, suggesting that the northern dense humid forests are growing under a more fragile equilibrium.
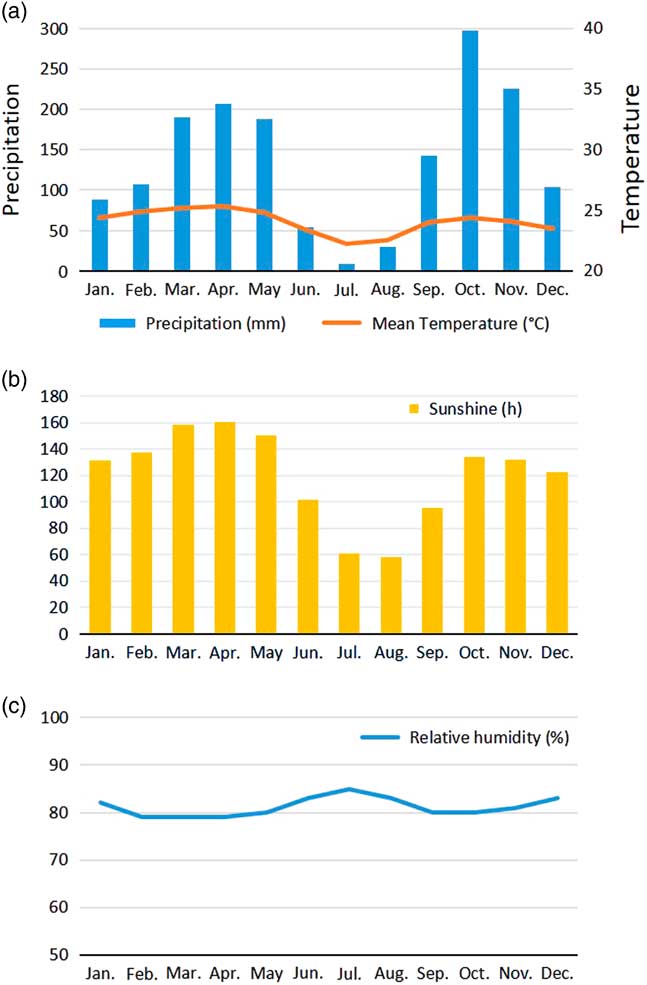
Figure 7 (color online) Yearly seasonal climatic diagram, Makokou station, northeastern Gabon. (adapted and updated from Doumenge, Reference Doumenge1990). (a) Mean monthly precipitation and temperatures. (b) Monthly sunshine variation. (c) Mean monthly relative humidity.
Historical and dynamic investigations
Charcoal found in soil profiles can provide precise information about the history and dynamics of forest formations (Hubau et al., Reference Hubau, Van den Bulcke, Kitin, Mees, Baert, Verschuren, Nsenga, Van Acker and Beeckman2013). Charcoal can be produced either from natural fires during dry periods, in relation to thunderstorms, or from anthropogenic activities linked to land clearing (Cochrane, Reference Cochrane2003). Several sites from the G. dewevrei forest formation have yielded fossil charcoal information, notably at Epulu and Yoko (both DRC).
In the Epulu area (1°20'N–28°35'E, Ituri region, northeastern DR Congo; Fig. 1, site 14), radiocarbon dating of charcoals indicates that this forest region was subjected to major disturbance during its post-4200 cal yr BP history (Hart et al., Reference Hart, Hart, Dechamps, Fournier and Ataholo1996), similar to that previously described in southern Cameroon and western Gabon (Bostoen et al., Reference Bostoen, Clist, Doumenge, Grollemund, Hombert, Muluwa and Maley2015). Indeed, in this area, which is more than 100 km west of the Eastern African savanna, Hart et al. (Reference Hart, Hart, Dechamps, Fournier and Ataholo1996) dated at ca. 2200 cal yr BP charcoal samples identified as Trichilia emetica, a tree typical of savanna and woodlands, and Tarchonanthus camphoratus, a taxon typical of open stony environments in woody savanna. The presence of these savanna species confirms that savannas and woodlands had replaced a previous forest, at least partially. Moreover, these authors note the almost complete local disappearance of various Fabaceae-Caesalpinioideae species at Epulu during the late Holocene and the rapid spread of G. dewevrei, which today has become one of the most abundant tree species in the forest of central Ituri.
The Yoko Forest Reserve site also show that G. dewevrei may have spread there from 2500–2000 cal yr BP. This site is situated ca. 20 km to the south-southeast of Kisangani (Fig. 1, site 17), and deeper in the Congo Basin than the Epulu site. In the Yoko forest, G. dewevrei is today the dominant tree in several sectors (Tshibamba, Reference Tshibamba2016; Lokonda, Reference Lokonda2017), and charcoal samples from G. dewevrei have been dated to between 2300 and 1400 cal yr BP (Tshibamba, Reference Tshibamba2016). However, many other charcoal fragments from the Yoko forest soil layers belonged to other taxa, so G. dewevrei was probably not the dominant species at these sites during this time. Furthermore, other data from the Yoko forest show that fires impacted forests in the region between ca. 2300 and 2200 cal yr BP (Lokonda, Reference Lokonda2017). Interestingly, the soils on the plateaus are richer in charcoal fragments than soils on slopes or in valley bottoms, which suggests stronger and more regular fires occurred on the plateaus (Lokonda, Reference Lokonda2017). All these data indicate that Limbali forests may have been less extensive before the 2500–2000 cal yr BP climate crisis and survived mainly in valley bottoms, from where they spread towards the plateaus about 2000 cal yr ago.
Recurring fire events on the rather vulnerable plateaus may have paved the way for the spread of Limbali. It is only more recently, since ca. 600 cal yr ago, that G. dewevrei trees have been largely burned in this area (Tshibamba, Reference Tshibamba2016). Comparing charcoal identifications with forest inventory data showed that the monodominant Limbali forests were sometimes converted to other forest types such as open-canopy Marantaceae forests (Tshibamba, Reference Tshibamba2016). The forest transformation could have been partly driven by heavier anthropogenic pressure since probably 600 cal yr ago, as artifacts were sometimes associated with the charcoal fragments (Tshibamba, Reference Tshibamba2016).
Other sites were studied in northern Congo where charcoal samples were collected (Gillet, Reference Gillet2013), mainly south of Mopo Bai (Fig. 1, site 11), including the sectors of Kabo (Fig. 1, site 16), limited to the east by the Congo swamps and as far south as Odzala-Kokoua National Park (Morin-Rivat et al., Reference Morin-Rivat, Fayolle, Gillet, Bourland, Gourlet-Fleury, Oslisly and Bremond2014; Fig. 1, site 18). Similar work was conducted in southeastern Cameroon, mainly between Lomié and Yokadouma, and up to southwestern Cameroon near the Campo-Ma’an National Park (Vleminckx et al., Reference Vleminckx, Morin-Rivat, Biwolé, Dainou, Gillet, Doucet, Drouet and Hardy2014). From a total of 43 radiocarbon dates obtained from these sites, 41 were more recent than ca. 2300 cal yr BP. They group into two main phases: the first one between 2300 and 1300 cal yr BP, and the second one between 670 and 250 cal yr BP. It is worth noting that the dates of the first period, although originating from different areas of the northern part of the rainforest domain, correspond to the beginning of the climate crisis when the fires were more widespread in open canopy forests and pioneer vegetation than in Limbali forests, which spread and have remained since that time (Gillet, Reference Gillet2013). These new data reinforce an earlier hypothesis of Aubréville (Reference Aubréville1957), then formulated by Maley (Reference Maley2001, Reference Maley2002) “that the G. dewevrei forest formation is a legacy of the disturbance about 2500 cal yr BP.
A new map of forest fragmentation between ca. 2500 to 2000 cal yr BP (Fig.1)
Palynological, biological, and climatic data allow us to estimate the delineation of the main large rainforest refuges in Central Africa during this last climatic crisis (Fig. 1), also referred to as “macro-refuges” (Ashcroft, Reference Ashcroft2010). The “central refuge” encompasses the central part of the Congo Basin (Fig. 1) and it is named a “fluvial refuge,” as it is favored by the availability of ground water close to the surface (Dupont et al., Reference Dupont, Jahns, Marret and Ning2000; Leal, Reference Leal2001). It includes the vast wetlands of the “Cuvette Centrale” (Betbeder et al., Reference Betbeder, Gond, Frappart, Baghdadi, Briant and Bartholomé2014; Dargie et al., Reference Dargie, Lewis, Lawson, Mitchard, Page, Bocko and Ifo2017). The central refuge is also recognized by its diversity of various mammal groups, particularly simian primates (Colyn et al., Reference Colyn, Gautier-Hion and Verheyen1991).
Its northern and eastern limits are circumscribed by the vast monodominant Limbali forests that mark areas where dense humid forests have been subjected to considerable changes beginning ca. 2500–2300 cal yr BP (see above). Its southern limits are between 2°S and 4°S. Both old and recent vegetation maps (Léonard, Reference Léonard1953; Mayaux et al., Reference Mayaux, Richards and Janodet1999) show several broad encroachments of savanna within dense forest towards the south of the central basin between 3°S and 4°S (Fig. 1). In contrast to the interpretation of Mullenders (Reference Mullenders1956), who considered these savannas to be recent consequences of human activities within the forest environment, it is clear that such savanna development could not have occurred as an isolated feature (Maley and Doumenge, Reference Doumenge2012). As shown for the Democratic Republic of Congo and Gabon (Delègue et al., Reference Delègue, Fuhr, Schwartz, Mariotti and Nasi2001; Vincens et al., Reference Vincens, Schwartz, Bertaux, Elenga and De Namur1998), these savannas were probably inherited from Holocene climate disturbances and a large proportion of the present dense forest is comparatively recent.
The only possible explanation for these open environments in southern DR Congo is that a large phase of forest perturbation and destruction occurred in the recent past after the substantial spread of forests during the early Holocene (Dalibard et al., Reference Dalibard, Popescu, Maley, Baudin, Melinte-Dobrinescu, Pittet, Marsset, Dennielou, Droz and Suc2014). Hence, this phase of forest destruction was very probably the massive one from ca. 2500 until 2000 cal yr BP. The borders of the savanna areas were then affected by the forest dynamics that isolated them within a spreading forest area (Maley and Doumenge, Reference Doumenge2012). This conclusion is important because these isolated savanna areas are an indicator of the southern extent of the forests in the “central” refuge (Fig. 1). Additionally, the extent of the vast monodominant Limbali forests help to circumscribe the northern limits of the central refuge.
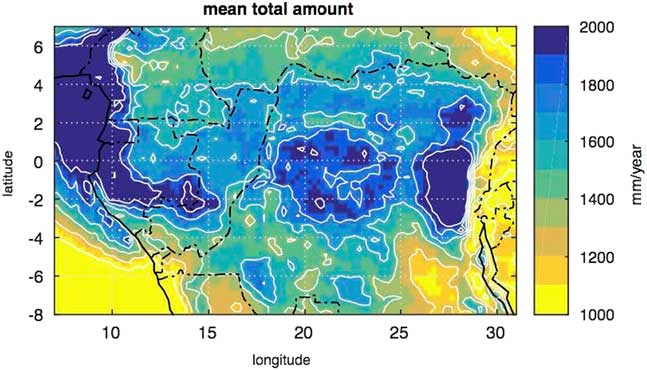
To the west, the central refuge is bordered by the SRI area, which delineates a large strip in the equatorial rainforest from the northwest region of the Congo Basin and extending southwards to the savannas of the Batéké Plateau. As explained above, the dense forests of the SRI area were highly disturbed and replaced by savannas and by extensive open canopy forests with an understory covered by giant herbaceous species, as in Odzala-Kokoua National Park (Fig. 1, site 18; Gillet, Reference Gillet2013; Gond et al., Reference Gond, Fayolle, Pennec, Cornu, Mayaux, Camberlin, Doumenge, Fauvet and Gourlet-Fleury2013). Moreover, the SRI area is also characterized by more extensive Limbali forest and drier semi-deciduous forest types than in the western and eastern regions (Gond et al., Reference Gond, Fayolle, Pennec, Cornu, Mayaux, Camberlin, Doumenge, Fauvet and Gourlet-Fleury2013). Furthermore, it is important to note that the SRI experiences slightly less precipitation than the western part of Atlantic Central Africa, i.e., in Gabon and the Central Congo Basin to the east (Fig. 8). The SRI is sheltered from the influence of the Equatorial Atlantic by small mountains and plateaus (ca. 700 to 1000 m asl) which border the Atlantic coast and extend until the eastern part of Gabon and the Congo. The SRI area lies at the eastern limit of the low cloud cover that develops in July–September over Gabon and southwestern Cameroon, which is characteristic of the dry season of the equatorial climate south of the climatic hinge, thus maintaining higher levels of relative humidity in Atlantic Central Africa compared to the SRI (Philippon et al., Reference Philippon, Lapparent, Gond, Sèze, Martiny, Camberlin and Cornu2016; Dommo et al., 2017).
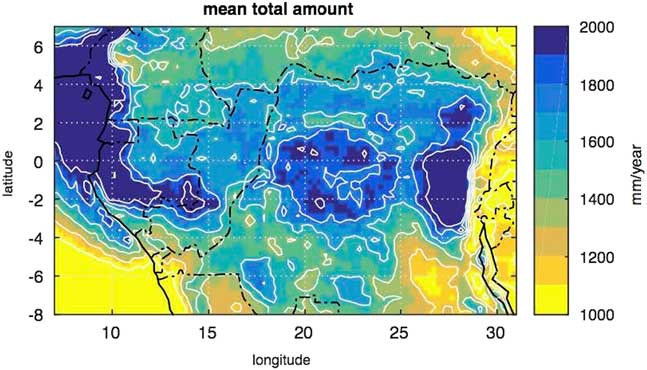
Figure 8 (color online) Mean annual rainfall in Central Equatorial Africa from the Tropical Rainfall Measurement Mission (TRMM) data for the period 1998–2015. The first maximum in the western part is due to moisture advection from the equatorial Atlantic within the low level branch of the Atlantic-Central Africa Walker cell. The secondary maximum in the central part is due to local recycling and convection within the ascending branch of the Walker cell.
Using all these available data, it is possible to precisely delineate the main rainforest refuge in the central part of the Congo Basin (Fig. 1), also referred to as a “macro-refuge” by Ashcroft (Reference Ashcroft2010). In the western part of Central Africa (southern Cameroon, Gabon, and Congo), other smaller refuges have been identified using palynological and paleoecological data, and also from previous refuges determined in this region for the major fragmentation phase during the LGM (Maley, Reference Maley1996, Reference Maley2001, Reference Maley2002, Reference Maley2012b; Maley and Brenac, Reference Maley and Brenac1998; Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004; Duminil et al., Reference Duminil, Mona, Mardulyn, Doumenge, Walmacq, Doucet and Hardy2015). These are complemented by smaller fluvial refuges along the Atlantic coast, where high water tables allowed the persistence of a mixture of inundated and terra firme forests.
In the eastern part of Central Africa, a lowland to submontane forest refuge is delineated on the western side of the Albertine Rift, where precipitation and cloudiness are important and provide sufficient water for dense humid forests (Fig. 1, site 19). The main western limit of the “oriental refuge” is represented by the areas where G. dewevrei forests are abundant.
Outside the main central refuge, forests were highly disturbed or disappeared, and replaced mainly by more-or-less drier forest types, wooded savannas, and woodlands. However, it should be stressed that, within these savanna and woodland regions, dense rainforest refuges were maintained (as “micro-refuges”) in favorable topographic positions (Leal, Reference Leal2001), which may have supported subsequent reforestation dynamics. The balance of savannas, woodlands, and dense forests (more-or-less disturbed) may have been different across the whole region, but this is difficult to assess precisely given the data available.
Various pollen data have shown that this period of strong disturbances ended at about 2000 cal yr BP with a new phase of forest expansion (Maley and Brenac, Reference Maley and Brenac1998; Maley, Reference Maley2001, Reference Maley2002, Reference Maley2004; Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004). Indeed, this new forest expansion continued, with some variations, during the last millennium (Ngomanda et al., Reference Ngomanda, Chepstow-Lusty, Makaya, Schevin, Maley, Fontugne, Oslisly, Rabenkogo and Jolly2005) until the twentieth century, as illustrated, for instance, in various maps of southern Cameroon (Letouzey, Reference Letouzey1985) and DR Congo (Mayaux et al., Reference Mayaux, Richards and Janodet1999), as well as by field observations in Central African countries (Maley and Doumenge, Reference Doumenge2012), and also in several countries of Western Africa, particularly in Ivory Coast, going back two centuries (Fairhead and Leach, Reference Fairhead and Leach1998).
In conclusion, these various data confirm the very widespread nature of the environmental disturbances that occurred between ca. 2500 and 2000 cal yr BP in the forest domain of Equatorial Central Africa, as already demonstrated in the western part of this large region (Maley and Brenac, Reference Maley and Brenac1998; Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004; Giresse et al., Reference Giresse, Maley and Kossoni2005).
PALEOCLIMATIC INTERPRETATIONS – COMPARISONS WITH PRESENT-DAY CLIMATIC VARIATIONS
Phase ca. 4000 to 2500 cal yr BP
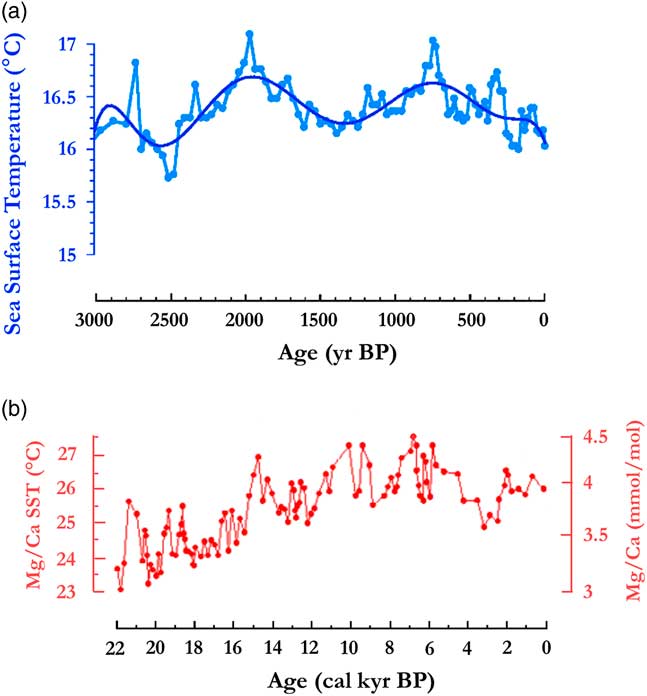
To understand the climate context that led to the large disturbance and fragmentation of the Central African rainforest between 2500 and 2000 cal yr BP, it is important to first examine a previous phase starting around 4000 cal yr BP and extending until ca. 2500 cal yr BP. This phase exhibited extensive savannas spreading at the periphery of the central forest domain, not only in southern Cameroon, where the first phase of Bantu expansion occurred (Bostoen et al., Reference Bostoen, Clist, Doumenge, Grollemund, Hombert, Muluwa and Maley2015), but also more to the south in the southern and western Congo areas. The savannas expanded in the Niari Region, as recorded by the drying out of lake Sinnda (Vincens et al., Reference Vincens, Schwartz, Bertaux, Elenga and De Namur1998; Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004), and also in Gabon where the littoral savannas developed, as revealed by the lake Maridor record (Giresse et al., Reference Giresse, Makaya, Maley and Ngomanda2009). In the western part of the Guinean forest domain, the forests contracted around 4000 cal yr BP, favoring the rapid expansion of savannas. Moreover in present-day Bénin and Togo, the savannas extended up to the Gulf of Guinea, opening up what biogeographers call the “Dahomey Gap”. Studying this phenomenon, Salzmann et al. (Reference Salzmann and Hoelzmann2005) showed that it was primarily linked to an abrupt SST (sea-surface temperature) cooling in the Gulf of Guinea (Fig. 9b; Weldeab et al., Reference Weldeab, Schneider and Müller2007), which modified the functioning of the African monsoon, increasing the length of the dry season at the periphery of the rainforest and inducing the spread of savannas. During this period, however, the rainforest persisted in the central core area, particularly for the evergreen types.
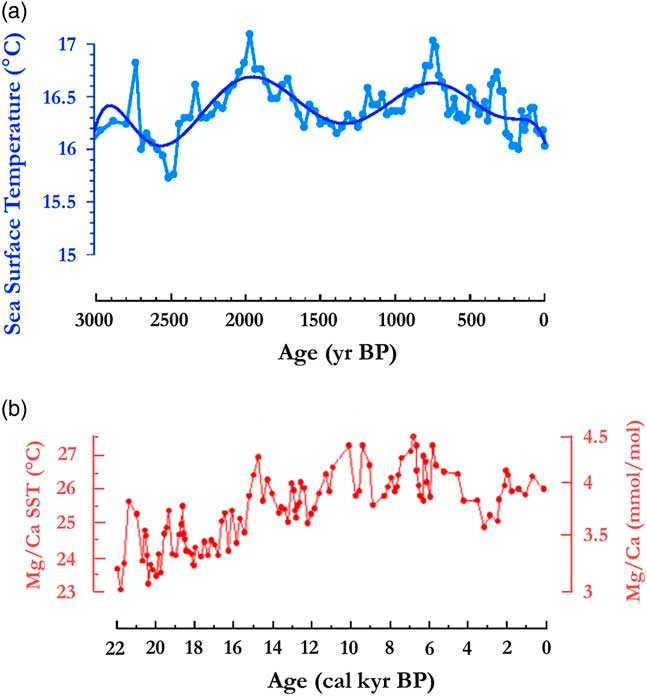
Figure 9 (color online) Sea surface temperature variations in the Gulf of Guinea. (a) Benguela sector, ca. 3000 to 0 yr BP (from fig. 3c in Leduc et al., Reference Leduc, Herbert, Blanz, Martinez and Schneider2010). Ooffshore Cameroon c. 22,000 to 0 yr BP (from fig. 3a in Weldeab et al., Reference Weldeab, Schneider and Müller2007).
The persistence of rainforest ca. 4000 cal yr BP is confirmed by the fact that typical evergreen rainforest taxa, such as Fabaceae-Caesalpinioideae and Sapotaceae, exhibited a marked increase during this phase, as recorded in several pollen profiles from southern Cameroon (Maley, Reference Maley2002) in lakes Barombi-Mbo (Maley and Brenac, Reference Maley and Brenac1998) and Ossa (Giresse et al., Reference Giresse, Maley and Kossoni2005) and the Nyabessan swamp (Ngomanda et al., Reference Ngomanda, Neumann, Schweitzer and Maley2009), as well as in central Gabon at lake Nguene near the Cristal Mountains (Giresse et al., Reference Giresse, Makaya, Maley and Ngomanda2009).
A climatic explanation for the persistence of a continuous forest block in Central Africa with evergreen forests might come from an understanding of the zonal Walker circulation (Fig. 8). Cooler SSTs in the Equatorial Atlantic favored expansion of savannas at the northern and southern peripheries of the forest domain and, by modifying the temperature gradients between the Atlantic and Central Africa, might have strengthened the zonal Walker circulation cell over the Atlantic-Central Africa zone (Wu et al., Reference Wu, Schurgers, Rummukainen, Smith, Samuelsson, Jansson, Siltberg and May2016). Currently, this circulation, mainly developing in June–October (Nicholson and Grist, Reference Nicholson and Grist2003; Cook and Vizy, 2015; Neupane, Reference Neupane2016) is responsible for humidity advection inland (Pokam et al., 2015), convection in the Central Basin and a persistent low cloud cover in Atlantic Central Africa (Fig. 7, climatic diagram at Makokou, eastern Gabon; Dommo et al., 2017). These climatic conditions currently support a continuous rainforest block through Central Africa and evergreen taxa in its western part. Indeed, the evergreen taxa are adapted to capture the atmospheric humidity from the fog in the canopy and low clouds trapped along the mountain flanks. This compensates for the low rainfall induced by these non-precipitating clouds, and allows evergreen forests to persist (Maley and Brenac, Reference Maley and Brenac1998). Currently, this ecological behavior of the western Central Africa evergreen taxa is similar to the adaptations of those species, which permit the development of montane forests (i.e., “cloud forests”; Maley and Elenga, Reference Maley and Elenga1993; Elenga et al., Reference Elenga, Maley, Vincens and Farrera2004).
Phase ca. 2500 – 2000 cal yr BP
The following phase started around 2500 cal yr BP, when the SSTs in the Equatorial Atlantic abruptly increased and remained abnormally warm until ca. 2000 cal yr BP (Fig. 9a and 9b; Weldeab et al., Reference Weldeab, Schneider and Müller2007; Leduc et al., Reference Leduc, Herbert, Blanz, Martinez and Schneider2010). These warm SST conditions might have weakened both the zonal Walker circulation cell over the Atlantic-Central Africa zone and the West African monsoon, maintaining the ITCZ abnormally southwards (Fig. 8). As a result, rainfalls and seasonality might have been exacerbated over Central Africa in the following ways: (1) a strong enhancement of local convection within the ITCZ, leading to an intensification of rainfall (i.e., higher occurrence of extreme to very extreme rainy events); and (2) an intensification in Central Africa of the main dry season, from June to September, and the associated low cloud cover that sustains the evergreen taxa.
These two aspects are supported by several types of paleoenvironmental data. The increase in rainfall seasonality and intensification of the dry season have been substantiated at two archaeological sites from southern Cameroon with the discovery of numerous charred grains of Pennisetum glaucum (pearl millet), a typical semi-arid sub-Saharan cereal, dated ca. 2400– 2200 cal yr BP (Neumann et al., Reference Neumann, Bostoen, Höhn, Kahlheber and Ngomanda2012a). These authors concluded that the cultivation of pearl millet was only possible at this time due to a large increase in seasonality.
The increase in rainfall intensity is also corroborated by the emplacement of large detritic levels between ca. 2500 and 2000 cal yr BP, frequently linked to a stone-line sequence. In Gabon, the movement of huge stones, some more than 2 m in size (Thieblemont et al., Reference Thiéblemont, Flehoc, Ebang-Obiang, Rigollet, Prian and Prognon2013), cannot be the result of a torrential regime, given the low slopes of this region, because the rivers’ transport capacity is too low (Maley et al., Reference Maley, Giresse, Favier and Doumenge2014). It is thus necessary to envisage large landslides transporting, for instance, the large broken pieces of trees found in the thick reworked level of Kakamoéka in Mayombe (Fig. 2; Maley and Giresse, Reference Maley and Giresse1998); or envisage debris flows or huge creep processes (Maley et al., Reference Maley, Giresse, Favier and Doumenge2014). Hence, the period with stone-line emplacement would not be necessarily overall rainier, but characterized by several rainy paroxysms acting in the same regions (Nicholson, Reference Nicholson2000; Trenberth and Shea, Reference Trenberth and Shea2005). Furthermore, these extreme rain events must be associated with a period of more seasonal climate, linked to abrupt periods of large SSTs increasing in the Equatorial Atlantic (Maley, Reference Maley2002; Weldeab et al., Reference Weldeab, Schneider and Müller2007; Leduc et al., Reference Leduc, Herbert, Blanz, Martinez and Schneider2010; Maley, Reference Maley2012b).
A climatic analog in the present meteorological record that gives support to such climatic conditions is the anomalous June–September season of 1984 (Guillot, Reference Guillot1986; Buisson, Reference Buisson1985). As indicated above, the June–September period corresponds to a well-developed zonal Walker circulation over the Atlantic-Central Africa zone associated with cool SSTs in the southern and equatorial Atlantic, and an ITCZ located at ~10°N (Fig. 8). In 1984, however, the SSTs remained anomalously warm in the Equatorial Atlantic, leading to large hydro-climatic anomalies over Central Africa and West Africa, which recorded its wettest rainy season since the 1950’s (Mahé and Citeau, Reference Mahé and Citeau1993). June–September 1984 was characterized by a high frequency of thunderstorms in Gabon and Congo, extending into central DR Congo, and as far as the East African Great Lakes (Guillot, Reference Guillot1986). These frequent thunderstorms were conducive to major erosion: this explains the largest discharges observed over the past decades for many rivers in southern Cameroon and Gabon, for instance the Nyong, Kienke, Kouilou, and Ogooue rivers (Mahé and Olivry, Reference Mahé and Olivry1995; Lienou et al., Reference Liénou, Mahé, Paturel, Servat, Sighomnou, Ecodeck, Dezetter and Dieulin2008; Mahé et al., Reference Mahé, Lienou, Descroix, Bamba, Paturel, Laraque and Meddi2013). Currently, rainfall and discharge for many rivers in Central Africa have been increasing constantly in July–August since the 1970’s (e.g., as attested by the Congo River discharge time series at the Brazzaville recording station [De Wasseige et al., Reference De Wasseige, Marshall, Mahé and Laraque2015; Tsalefac et al., Reference Tsalefac, Hiol Hiol, Mahé, Laraque, Sonwa, Sholte and Pokam2015]). In this way, 1984 can provide a model for the period ca. 2500–2000 cal yr BP, because the intense erosion observed that year resulted from the high frequency of stormy rainfall.
With regard to changes in rainfall seasonality, this phenomenon was particularly conspicuous in Central Africa near the Atlantic, especially in Gabon in 1984. For instance, Libreville recorded 96 mm in June–September 1984 (the normal is ~50 mm). Such an amount is equivalent to the October–December rainy season. In June–September 1983, when SST cooling in the Equatorial Atlantic was very marked, Libreville recorded only 38 mm (Buisson, Reference Buisson1985).
The warm SST anomaly in July–September 1984 in the Equatorial Atlantic suggests the existence of an El Niño-type response for the Eastern Tropical Atlantic (Hisard, Reference Hisard1980; Nicholson and Entekhabi, Reference Nicholson and Entekhabi1987; Binet et al., Reference Binet, Gobert and Maloueki2001). Teleconnections occur regularly between the Pacific and the Atlantic Oceans in response to the zonal Walker atmospheric circulation along the equator (Philander, Reference Philander1986; Delecluse et al., Reference Delecluse, Servain, Levy, Arpe and Bengtsson1994), with some major episodes, such as in 1983 and 1984, being characterized by the opposite behavior of SSTs between the two oceans. During the El Niño event (warm SST in the Eastern Pacific) of 1983, the SSTs were anomalously cold in the Equatorial Atlantic. During the following year’s (1984) La Nina event (cold SSTs in the Eastern Pacific), warm SSTs extended largely into the Equatorial Atlantic (Delecluse et al., Reference Delecluse, Servain, Levy, Arpe and Bengtsson1994).
These teleconnections hold true for the paleoclimates. Toth et al. (Reference Toth, Aronson, Vollmer, Hobbs, Urrego, Cheng, Enochs, Combosch, Van Woesik and Macintyre2012) showed that, beginning from ca. 4000 cal yr BP, very strong El Niño events, coupled with a high variability in the latitudinal migration of the ITCZ, caused the collapse of eastern Pacific coral reefs, because their mortality was induced by frequent episodes of warm water. At the same time, SSTs were cool in the eastern Atlantic. From 2500 until 2000 cal yr BP, a decoupling of ENSO and the ITCZ occurred in the Pacific, linked to an increase of La Niña activity (Lachniet et al., Reference Lachniet, Bernal, Asmerom, Polyak and Piperno2012), while SSTs in the equatorial Atlantic were warm. Hence, these two periods exhibited climatic teleconnections between the Atlantic and Pacific Oceans.
Comparison of the ca. 2500–2000 cal yr BP phase with a similar phase close to the end of the Pleistocene
As discussed, an important detrital period, synchronous with a large increase in the Congo River discharge, was dated from ca. 14,500 to 13,000 cal yr BP (Fig. 4; Giresse et al., Reference Giresse, Bongo-Passi, Delibrias and Duplessy1982; Bayon et al., Reference Bayon, Dennielou, Etoubleau, Ponzevera, Toucanne and Bernell2012). It was simultaneous with the first stone-line reported across Central Africa, particularly in Gabon, and observed in Kakamoéka from the western part of Central Africa, and in Osokari from the eastern part (Fig. 2 and 3). After ca. 13,000 and until ca. 11,000 cal yr BP, the detrital sediments greatly diminished in the Congo deep-sea fan, synchronous with a major decline of river discharge (Bayon et al., Reference Bayon, Dennielou, Etoubleau, Ponzevera, Toucanne and Bernell2012). Therefore, erosion was also greatly reduced during this period, which was synchronous with the cold Younger Dryas period (YD) in the Northern Hemisphere (Fig. 10a and b). Moreover, high river discharge was similarly observed in East Africa at ca. 14,500 cal yr BP, first connected with the abrupt refilling of Lake Victoria and subsequently to the overflow of the White Nile, leading to extensive flooding (Williams et al., Reference Williams, Talbot, Aharon, Salaam, Williams and Brendeland2006). This marked rise in discharge extended up to the Egyptian Nile Valley, where floods of exceptionally high levels have been described and called the Wild Nile phase (Butzer, Reference Butzer1980; Paulissen and Vermeersch, Reference Paulissen and Vermeersch1987; Maley, Reference Maley2000).
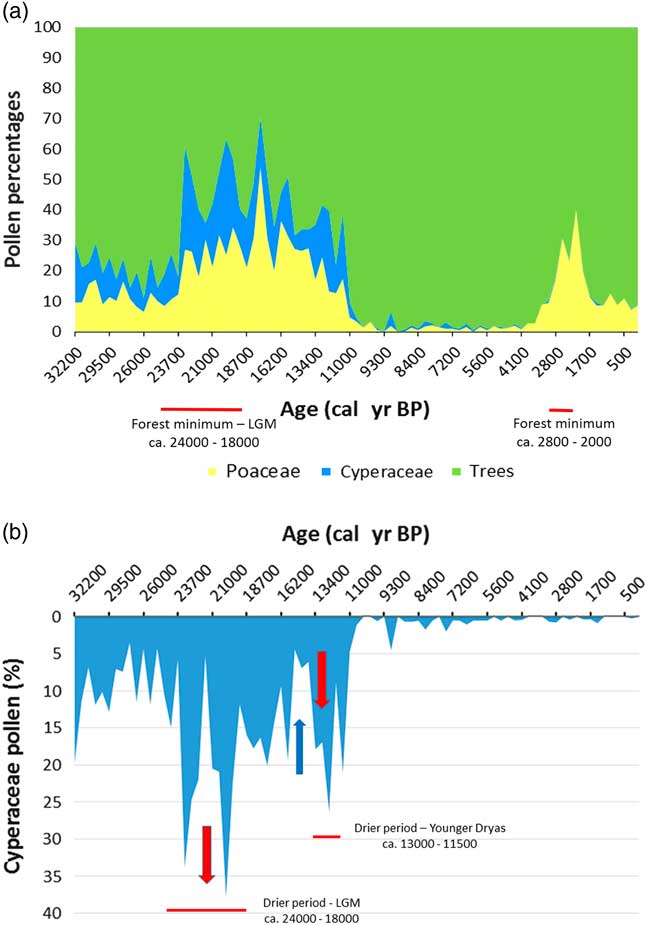
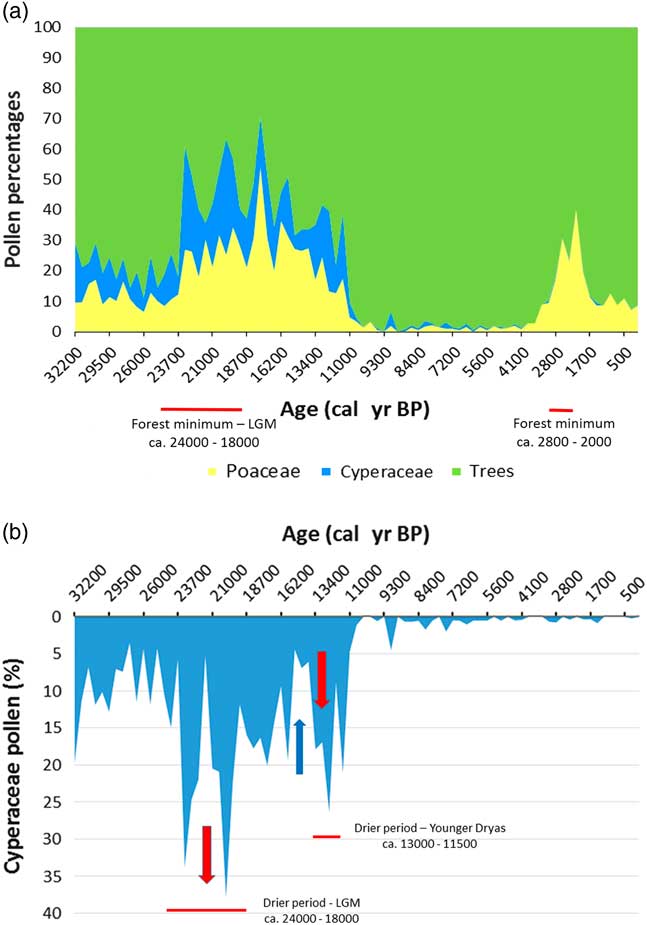
Figure 10 (a) Lake Barombi Mbo synthetic pollen diagram for the last 32,000 cal yr BP: Poaceae pollen (yellow) for savanna variations, combined with Cyperaceae pollen (wetland taxa, blue) for each level, the remaining percentages correspond to forest trees pollens (adapted from Maley and Brenac, Reference Maley and Brenac1998). (b) Barombi Mbo, lake level variations, using Cyperaceae pollen as proxy data (Maley, Reference Maley2012a). Red arrow, regression; blue arrow, transgression. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Similar variations in flood frequency were observed in western Cameroon at lake Barombi Mbo, the only central equatorial site extending its record of past environmental change from the current period until ca. 32,000 cal yr BP (Fig. 10a; Maley and Brenac, Reference Maley and Brenac1998). Indeed, at lake Barombi Mbo, which exhibits a maximum depth of 110 m, Cyperaceae, typical wetland herbs, spread usually on the western deltaic platform when the lake level decreases. Hence, the relative abundance of Cyperaceae pollen can be used as a proxy indicator for reconstructing lake-level variation (Fig. 10b). This is particularly the case before the beginning of the Holocene, as during this period the lake level was stabilized by an overflow outlet (Maley and Brenac, Reference Maley and Brenac1998). During the LGM, Cyperaceae pollen increased sharply (Fig. 10b), indicating a reduction in lake level of ca. 6 m until the edge of the deltaic platform. Subsequently, between ca. 15,000 and 13,000 cal yr BP, lake levels rose markedly, synchronous with the Congo River discharge increase and also the strong erosional phase characterized by the first stone-line (Fig. 4).
Afterwards, concomitant with the YD period, ca. 13,000 to 11,500 cal yr BP, a sharp decline in lake-level at lake Barombi Mbo occurred. Therefore, during the intermediary warming phase, synchronous with the Bölling-Allerod period (ca. 15,000 to 13,000 cal yr BP), the abrupt decrease of Cyperaceae pollen is related to a strong transgressive phase at the same time as the main erosional phase linked to the first stone-line. Moreover, the SST variations in the Gulf of Guinea exhibited a similar marked increase during the two erosional phases linked to the first (ca. 15,000 to 13,000 cal yr BP) and the second stone-line (ca. 2500–2000 cal yr BP; Fig. 6b, Weldeab et al., Reference Weldeab, Schneider and Müller2007). Therefore, the changes in the monsoon described for the second early late Holocene stone-line may also have applied during the Bölling-Allerod warm period (Fig. 10a).
CONCLUSIONS AND CLIMATIC IMPLICATIONS
From a present-day perspective, as the fragmentation of the Central African rainforests took place between ca. 2500 and 2000 cal yr BP, and overall during a rather “warm” period (Fig. 9a), this phase may provide a model for what might take place during the current period of “global warming” (Trenberth and Shea, Reference Trenberth and Shea2005). In this framework, predicted changes for the Earth´s vegetation are frequently discussed, particularly for the most vulnerable types such as tropical rainforests.
Several papers have examined the behavior of tropical rainforests in Central Africa, mainly in the Congo Basin. Zhou et al. (Reference Zhou, Tian, Myneni, Ciais, Saatchi, Liu, Piao and Chen2014) present evidence for a widespread decline in forest greenness over the past few decades, partially attributable to a long-term drying trend. Zelazowski et al. (Reference Zelazowski, Malhi, Huntingford, Sitch and Fisher2011) have also examined changes in the potential distribution of humid tropical forests under a warmer climate. In using an empirical approach together with projections from several coupled Atmosphere-Ocean-General Circulation Models, these authors concluded for the Congo Basin that climate change will manifest itself particularly in the rainfall regime and plant water deficit (i.e., evapotranspiration). Moreover, some models predict scenarios of forest retreat, possibly leading to the split of the forest cover into separate parts.
The scenario of forest fragmentation can be examined in the light of new climatic data about present-day lightning activity. Both Christian et al. (Reference Christian, Blakeslee, Boccippio, Boeck, Buechler, Driscoll, Goodman, Hall, Koshak, Mach and Stewart2003), from space observations, and Soula et al. (Reference Soula, Kigotsi Kasereka, Georgis and Barthe2016), using mainly the Lightning Imaging Sensor, concluded that the highest concentrations of lightning on the Earth are presently observed in Central Africa and mainly in DR Congo, with the maximum of annual flash density and number of stormy days. During their period of observation (2005 to 2013), Soula et al. (Reference Soula, Kigotsi Kasereka, Georgis and Barthe2016) noted that the maximum activity occurred during the austral summer, from November to May, and also during the boreal summer, from June to August, because, as shown by Collier and Hughes (Reference Collier and Hughes2011), the peak lightning follow the migration of the ITCZ. Its activity is primarily associated with violent phenomena such as thunderstorms and convective rains, which require regional sources of humidity such as the swampy forests in the central part of the Congo Basin and also the lakes close to the Rift (Victoria, Edward, and Kivu). Soula et al. (Reference Soula, Kigotsi Kasereka, Georgis and Barthe2016) also observed that the lightning density can be very high in some years near the coast of Congo and Gabon, such as in 2007, where the Gulf of Guinea provides another important humidity source for thunderstorms. Moreover, as discussed to explain the ca. 2500 cal yr BP phase, Guillot (Reference Guillot1986) showed that the 1984 summer season in Gabon was also characterized by a high frequency of thunderstorms, particularly because the SSTs in the Gulf of Guinea were very warm during this summer season.
Important advances in the areas of lightning and thunderstorm research have been made by several authors such as Rycroft et al. (Reference Rycroft, Israelsson and Price2000) and Singh et al. (Reference Singh, Gopalakrishnan, Singh, Kamra, Singh, Pant and Singh2007). Indeed, lightning activity and the global electric circuit, which are closely related around the Earth, are also associated with some important climate parameters. These authors have shown that the solar wind may influence the global electric circuit by effects on cloud microphysics and other dynamic effects in the troposphere. Within the framework of these possible Sun-Earth interactions, Chase et al. (Reference Chase, Meadows, Carr and Reimer2010) showed that solar forcing modulated by variations in the Earth’s geomagnetic shield is “a potentially important factor driving climate at suborbital timescales in both the northern and southern tropics.” In Africa particularly, “the minimum of the geomagnetic dipole moment was linked to a relatively humid phase (at 8000–7000 cal yr BP); the sharp increase in dipole strength at ca. 3800 cal yr BP is concurrent with the beginning of a drought phase, and the maximum dipole moment with a relatively arid phase (at 2500–2000 cal yr BP)” (Chase et al., Reference Chase, Meadows, Carr and Reimer2010, p.42, fig. 3 and 6). These results are important because the last two phases described by these authors correspond to the two late Holocene phases discussed here. In the forests of Central Africa, the marked seasonality increase for the phase ca. 2500 - 2000 cal yr BP was indeed a factor of “relative” climate aridification, because, as explained, it was linked to stronger seasonality with a more important dry season counterbalanced by a higher frequency of stormy rainfall, leading to the deposition of major detritic levels and stone-line emplacements.
Moreover, in the framework of the climatic teleconnections occurring between the Atlantic and the Pacific Oceans, Emile-Geay et al. (Reference Emile‐Geay, Cane, Seager, Kaplan and Almasi2007) and Gray et al. (Reference Gray, Beer, Geller, Haigh, Lockwood, Matthes and Cubasch2010) estimated that solar influences could have impacted the climatic circulation above the Pacific Ocean, as ENSO and ITCZ activity, and hence could have played a role in these climatic teleconnections, as described during the late Holocene. As the effect of these Sun-Earth interactions on global climates remains a matter of debate, it appears that the recent increase in lightning activity in Central Africa, mainly in the Congo Basin, could result from some kind of solar influence, as was also the case for forest fragmentation between ca. 2500 and 2000 cal yr BP. Further research in these fields is needed.
ACKNOWLEDGMENTS
Brian Chase is thanked for his help in providing advice and data. W. Gosling and D. Booth (QR Editorial Board) are thanked for their constructive comments. J. Maley expresses his gratitude and his friendship to the numerous African colleagues from Cameroon, Gabon, and Congo, with whom he worked in order to obtain much of the data presented in this paper.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/qua.2017.97