


INTRODUCTION
Predicting the response of species to climate change has become an extremely active field of research because of the development of a better understanding of the mechanisms by which species and ecosystems can be affected by climate changes (e.g., Bellard et al., Reference Bellard, Bertelsmeier, Leadley, Thuille and Carchamp2012). The mechanisms of the response of animal populations to climate fluctuations and environmental shifts can be also aided by study of past analogues (Hofreiter and Stewart, Reference Hofreiter and Stewart2009). The period of the late Pleistocene, especially its final part (Marine Oxygen Isotope Stage [MIS] 2), and the transition from the last glaciation to the Holocene (MIS 1), is an example of a time interval when major reorganization of animal communities and shifts occurred in species distributions as a response to abrupt climate fluctuations (e.g., Sommer and Nadachowski, Reference Sommer and Nadachowski2006; Stewart et al., Reference Stewart, Lister, Barnes and Dalén2010; Cooper et al., Reference Cooper, Turney, Hughen, Barry, Mcdonald and Bradshaw2015; Palkoupoulou et al., Reference Palkopoulou, Baca, Abramson, Sablin, Socha, Nadachowski and Prost2016; Baca et al., Reference Baca, Nadachowski, Lipecki, Mackiewicz, Marciszak, Popović, Socha, Stefaniak and Wojtal2017). However, the degree of precision in reconstruction of the late Pleistocene species’ range dynamics, migrations, and extinction events is related to the number of direct dates available for analysis. Among late Pleistocene large mammals the largest number of direct dates is available for the woolly mammoth (Mammuthus primigenius) (more than 1500 dates, according to Puzachenko et al. [Reference Puzachenko, Markova, Kosintsev, van Kolfschoten, van der Plicht, Kuznetsova, Tikhonov, Ponomarev, Kuitems and Bachura2017]); moreover, every year new dates are published. In the last decades, an increase in the number of direct radiocarbon dates on mammoth remains has allowed researchers to correlate shifts in distribution with even minor climatic events (e.g., Stuart et al., Reference Stuart, Sulerzhitsky, Orlova, Kuzmin and Lister2002, Reference Stuart, Kosintsev, Higham and Lister2004; Lorenzen et al., Reference Lorenzen, Nogués-Bravo, Orlando, Weinstock, Binladen, Marske and Ugan2011; Nadachowski et al., Reference Nadachowski, Lipecki, Wojtal and Miękina2011; Ukkonen et al., Reference Ukkonen, Aaris-Sørensen, Arppe, Clark, Daugnora, Lister and Lõugas2011).
We do not produce new dates here; instead we address the problem of climate change impacts on the range dynamics of the mammoth in Europe during the most severe period of the late Pleistocene, MIS 2, on the basis of already published results. For this purpose, we gathered direct radiocarbon dates of the woolly mammoth and plotted them against the latitude of sites and the short climatic intervals distinguished within MIS 2 in Europe (Rasmussen et al., Reference Rasmussen, Bigler, Blockley, Blunier, Buchardt, Clausen and Cvijanovic2014). The problem of defining the changes in the European distribution of the mammoth has been addressed recently by Markova et al. (Reference Markova, Puzachenko, van Kolfschoten, van der Plicht and Ponomarev2013) and Puzachenko et al. (Reference Puzachenko, Markova, Kosintsev, van Kolfschoten, van der Plicht, Kuznetsova, Tikhonov, Ponomarev, Kuitems and Bachura2017); however, in these studies the quality of radiocarbon dates was not evaluated. What is more, they tracked changes of mammoth distribution within relatively long time intervals (e.g., the last glacial maximum [LGM] defined as the period between Greenland stadial [GS] 4 and GS-2.1). Recently, it has been shown that late Pleistocene faunas were affected by abrupt and short-term climate changes such as the Greenland interstadial (GI) intervals (Cooper et al., Reference Cooper, Turney, Hughen, Barry, Mcdonald and Bradshaw2015); therefore, a coarse-grained analysis may fail to detect important events in the demographic history of a species.
A second aim of this article is to shed light on the relationship between the distribution of the woolly mammoth and humans from an archaeological perspective.
MATERIALS AND METHODS
In recent years, thanks to new research projects, the amount of chronometric data for Mammuthus has increased, especially direct dates on mammoth material. We examined 318 radiocarbon dates (both conventional and accelerator mass spectrometry [AMS]) from published and a few unpublished sources (Table 1) coming from sites located across the whole of Europe, stretching east to the Ural Mountains and the Ural River (Supplementary Table 3), and we used these data to explore range shifts of M. primigenius in space and time intervals distinguished within MIS 2. We follow the MIS 2 definition of Andersen et al. (Reference Andersen, Svensson, Johnsen, Rasmussen, Bigler, Röthlisberger and Ruth2006) and Svensson et al. (Reference Svensson, Andersen, Bigler, Clausen, Dahl-Jensen, Davies and Johnsen2006), where GI-4 and GI-3 belong to MIS 3 and MIS 2 starts from the beginning of GS-3 and ends with the GI-1 warming.
Table 1 New accelerator mass spectrometry dates of Mammuthus primigenius remains from central Europe in Marine Oxygen Isotope Stage 2. Origin of the material dated: AIMM, Anthropos Institute, Moravian Museum, Brno, Czech Republic; ISEZ, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland; ML, Lublin Museum, Lublin, Poland; MMZ, City Museum in Żywiec, Poland; MOR, The Regional Museum in Rzeszów, Poland; MSO, Museum of Opole Silesia, Opole, Poland; MT, The Tatra Museum in Zakopane, Poland; UMCS, Maria Curie–Skłodowska University, Lublin, Poland.

a Date replaces conventional date (Lu-1346) 14,080±165 14C yr BP (Nadachowski et al., Reference Nadachowski, Lipecki, Wojtal and Miękina2011).
b Material extracted from a tusk of the G. Ossowski collection (nineteenth century).
c Date previously reported by Nadachowski et al. (Reference Nadachowski, Krajcarz, Krajcarz, Madeyska, Ridush, Valde-Nowak, Wojtal and Zarzecka-Szubińska2015) without laboratory number and detailed description of the sample.
Most of the European mammoth remains are bones or teeth recovered from Paleolithic archaeological sites deposited by humans or carnivores in situ in cultural layers, usually in well-recognized stratigraphic contexts. We retained the original numbering and/or subdivisions of large archaeological sites in spite of the fact that in most cases different sectors may be located close to each other or were later pooled into a single large site. For instance, for practical reasons we retained the subdivision of the Kraków Spadzista (Poland) site as five separate localities (B, C, C2, E1, and trench III), which correspond to various trenches but are now considered parts of one large campsite (Wilczyński et al., Reference Wilczyński, Wojtal and Sobczyk2012). The same system was used in the case of the Brno-Štýřice III (Czech Republic) site complex where four sites were distinguished in spite of fact that they belong to one Epigravettian settlement (Nerudová and Neruda, Reference Nerudová and Neruda2014). The Kostenki-Borshchevo complex of Paleolithic sites on the west bank of the Don River, in the central East European Plain (Russia), is another example (Praslov and Rogachev, Reference Praslov and Rogachev1982; Sinitsyn, Reference Sinitsyn2003; Anikovich and Platonova, Reference Anikovich and Platonova2014). This complex includes more than 60 Upper Paleolithic open-air sites (Zheltova, Reference Zheltova2015), and many of them are found not far distant from each other in the villages of Kostenki, Alexandrovka, and Borshchevo, along ravines and in till of the first and second terraces of the Don River. Most of the sites with mammoth remains analyzed in this article are located in the second terrace and are correlated with MIS 2 (Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997; Holliday et al., Reference Holliday, Hoffecker, Goldberg, Macphail, Forman, Anikovich and Sinitsyn2007).
In most cases, cultural archaeological levels containing mammoth remains are well dated, and ages are consistent, as in the case of layer 6 in the Kraków Spadzista site, where the medians of more than 30 AMS radiocarbon dates (Wojtal and Sobczyk, Reference Wojtal and Sobczyk2005; Arppe and Karhu, Reference Arppe and Karhu2010; Wilczyński et al., Reference Wilczyński, Wojtal and Sobczyk2012, Reference Wilczyński, Wojtal, Sobieraj and Sobczyk2015c) fall between ~29.0 and 27.3 cal ka BP. However, some dates, at least 8–10 of them, do not fit well to the proposed age of layer 6 (see Wilczyński et al., Reference Wilczyński, Wojtal, Sobieraj and Sobczyk2015c); these probably were obtained from remains deposited after the formation of the main occupation level. Six of these dates were accepted for further analyses (Supplementary Table 3).
Other dated remains of mammoths are isolated finds recovered from gravel pits, from river banks, or in glacial, fluviogacial, or loess sediments, sometimes transported either by the flow of streams or meltwaters from the retreating glaciers or by advancing ice sheets. In these cases, we have to take into account that material was shifted from the place of deposition, like erratics, and the transport distance in most cases is not known. However, taphonomic studies of mammoth finds have demonstrated that in Finland the final deposition of the remains from the original death site occurred in most cases less than 10 km away (Ukkonen et al., Reference Ukkonen, Lunkka, Jungner and Donner1999), and in Sweden less than 50 km away (Ukkonen et al., Reference Ukkonen, Arppe, Houmark-Nielsen, Kjær and Karhu2007). The next, rare category of direct dates analyzed here was taken from artifacts produced from mammoth bone or ivory. Almost all of them (except four) were rejected because of low collagen content, as in the case of the so-called boomerang from Obłazowa Cave, in Poland, manufactured from a mammoth tusk (Valde-Nowak et al., Reference Valde-Nowak, Nadachowski and Wolsan1987; Housley, Reference Housley2003; Davies et al., Reference Davies, White, Lewis and Stringer2015). In addition, we always have to take into account the possibility that precious items could have been transported over significant distances by people.
We focus on the dates for which there is little doubt about their origin, as well as on those dates that have been verified (Supplementary Tables 1, 2, and 3). Published dates from all laboratories, especially earlier ones based on non-ultrafiltered collagen, have been carefully analyzed according to the criteria given by Lister and Stuart (Reference Lister and Stuart2013). Criteria for rejection of the date included, first of all, whether these old conventional dates were replaced by later AMS dating of the same sample. Also, grounds for rejection of the earlier dating was inadequate identification—for instance, when the date was made on material other than Mammuthus or the dated skeletal element was not specified (e.g., ivory, tooth, bone). We always tried to return to the original description of the date because in quite a number of cases the citation of the dates by later authors did not match. The most common errors were connected with citing the wrong ranges of standard errors or using an incorrect order of numerals (swapping given numerals), as well as cases where dates had been originally taken on associated material (e.g., charcoal, other mammal species) and not on the mammoth (for details, see comments in Supplementary Table 3). Specimens were dated in many laboratories, and the methods used for pretreatment and dating were often different. This is an element of uncertainty in comparing dates. We are aware of the fact that dates obtained before 1980 are usually considered to require confirmation by redating before they can be accepted, primarily because of sample impurities (Stuart et al., Reference Stuart, Sulerzhitsky, Orlova, Kuzmin and Lister2002; Stuart and Lister, Reference Stuart and Lister2014); therefore, where possible, the AMS dates were used in our analysis. However, an improvement of the method does not automatically mean that any conventional date is wrong (Van der Plicht and Palstra, Reference Van der Plicht and Palstra2016). In some cases, old conventional dates and newer ones do not differ substantially. This is the case with the Lockarp specimen from south Sweden, dated by Berglund et al. (Reference Berglund, Håkansson and Lagerlund1976) and redated by Kjær et al. (Reference Kjær, Lagerlund, Adrielsson, Thomas, Murray and Sandgren2006) to a similar age. Another example where earlier and more recent dating agree is the youngest mammoth age in central Europe, determined on the mammoth skull from Bzianka, Poland (Kubiak, Reference Kubiak1980; Nadachowski et al., Reference Nadachowski, Lipecki, Wojtal and Miękina2011): the conventional date median is 17,113 cal yr BP (95.4% range, 17,585–16,585 cal yr BP; Lu-1346: 14,080±165 14C yr BP), which is similar to the redating median 16,844 cal yr BP (95.4% range, 17,104–16,548 cal yr BP; Poz-39511: 13,900±70 14C yr BP) obtained on the same specimen (Table 1). The homogeneity of dates was confirmed with χ2-test performed in OxCal v. 4.2 (df=1, T=1 [5%, 3.8]). Compiling dates from different dating laboratories and using both conventional and AMS methods increased the number of dates in our study; we reasoned that excluding dates only because they are conventional risks losing novel information. When two (or more) dates have been obtained on the sample skeletal elements, the older (conventional) dates were ignored or rejected and replaced by AMS dates, or in some cases the results were averaged using the “combine” function in OxCal. The combined dates are listed in Supplementary Tables 2 and 3.
Unfortunately, there are many more mammoth finds in Europe that have not been dated and, therefore, could not be included in our sample. Lack of direct dating on so many finds might lead to incorrect interpretations of the distribution of mammoths in time and space, especially in areas south of the Carpathians, Alps, and Pyrenees. For example, ~400 remains of Mammuthus, including six almost complete skeletons, have been recovered in the Pannonian Basin from MIS 2 and MIS 3 contexts, but among these, only three specimens were directly dated in Hungary (Konrád et al., Reference Konrád, Kovács, Halász, Sebe and Pálffy2010; Katona et al., Reference Katona, Kovács, Kordos, Szappanos and Linkai2012).
The analysis of absolute datings was performed using Bayesian models by means of the OxCal 4.2.4 software (Bronk Ramsey, Reference Bronk Ramsey2009; Bronk Ramsey et al., Reference Bronk Ramsey, Scott and van der Plicht2013). Dates are presented in a chronological chart (Supplementary Table 2) from the youngest to the oldest one used in the analysis. The obtained radiocarbon ages were calibrated with the IntCal13 radiocarbon atmospheric calibration curve for terrestrial samples (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronk Ramsey and Buck2013). All calibrated dates are presented in calibrated years BP, with 95.4% probability ranges. Changes in mammoth distribution are also presented in a series of maps (Figs. 1–9) corresponding to eight time intervals that match GS and GI intervals distinguished within the MIS 2 (GS-3, GI-2, GS-2.1c, GS-2.1b, and GS-2.1a) and adjacent time periods covering the end of MIS 3 (GS-4 and GI-3) and the onset of MIS 1 (GI-1e, Bølling). Chronological ranges of particular time slices, especially the beginning and the end of a given interval, followed the updated INTIMATE event stratigraphy (Rasmussen et al., Reference Rasmussen, Bigler, Blockley, Blunier, Buchardt, Clausen and Cvijanovic2014). The median value of the date (after calibration) determined the stadial/interstadial event that would include the given radiocarbon date. Dates with medians close to the GS/GI boundaries were verified on whether the probability of a date was unimodal or not, and as a consequence, multimodal dates were rejected. Maps were plotted in QGIS v. 2.4 (QGIS Development Team, 2009); the extent of ice sheets is from Hughes et al. (Reference Hughes, Gyllencreutz, Lohne, Mangerud and Svendsen2016), and changes of eustatic sea levels around Europe are from Fleming et al. (Reference Fleming, Johnston, Zwartz, Yokoyama, Lambeck and Chappell1998).
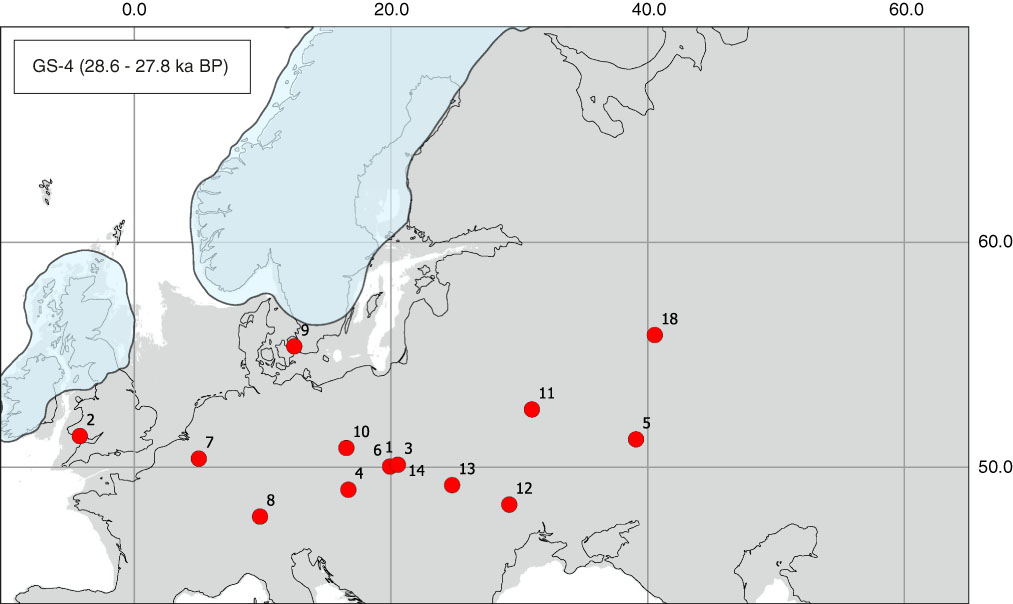
Figure 1 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland stadial 4 (GS-4). Sites (number of dates in parentheses): 1, Kraków Spadzista E1 (3); 2, Paviland Cave (1); 3, Jaksice II (1); 4, Dolní Věstonice (1); 5, Kostenki 12 (1); 6, Kraków Spadzista B (5); 7, Goyet, Third Cave (1); 8, Hochwacht (1); 9, Kopenhagen, Kamstrup (1); 10, Zastruże (1); 11, Berdyzh (1); 12, Leski (1); 13, Halych I (1); 14, Kraków Spadzista C2 (1); 18, Sungir (1).
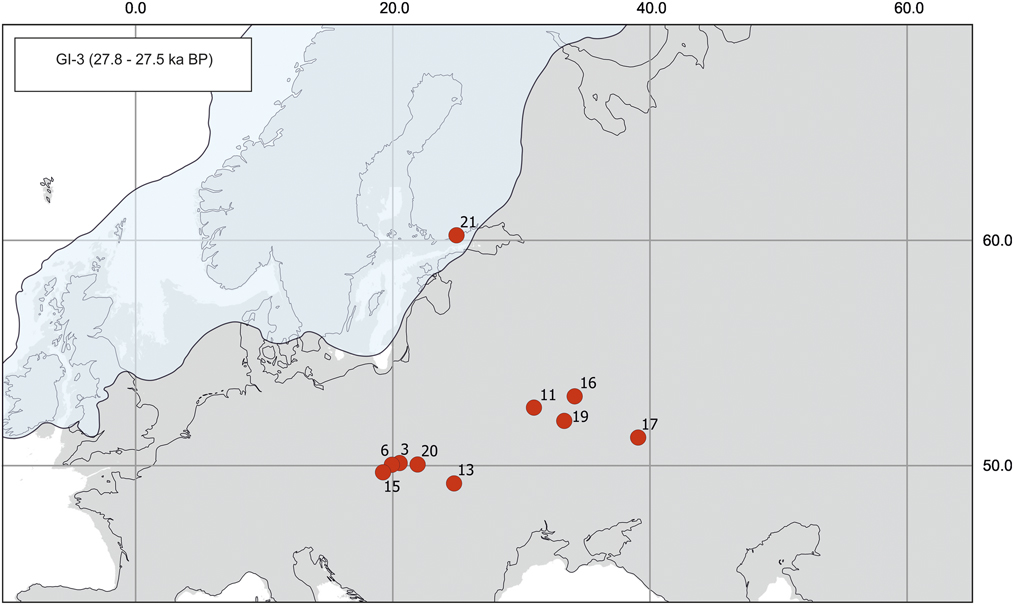
Figure 2 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland interstadial 3 (GI-3). Sites (number of dates in parentheses): 3, Jaksice II (1); 6, Kraków Spadzista B (1); 11, Berdyzh (1); 13, Halych I (1); 15, Żywiec, Koszarawa River (1); 16, Khotylevo 2 (4); 17, Kostenki 1 (1); 19, Pogon (1); 20, Świlcza (1); 21, Helsinki, Töölö (1).
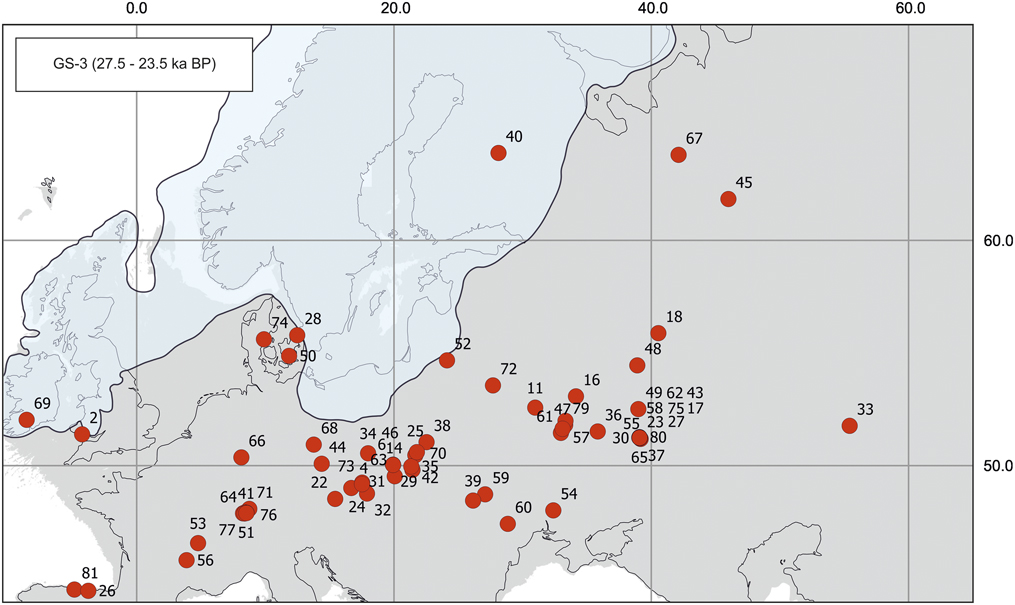
Figure 3 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland stadial 3 (GS-3). Sites (number of dates in parentheses): 2, Paviland Cave (2); 4, Dolni Věstonice (2); 6, Kraków Spadzista B (2); 11, Berdyzh (1); 14, Kraków Spadzista C2 (5); 16, Khotylevo 2 (8); 17, Kostenki 1 (10); 18, Sungir (2); 22, Willendorf II (1); 23, Kostenki 2 (1); 24, Jarošov I (2); 25, Sandomierz (1); 26, Pámanes (2); 27, Kostenki 17 (2); 28, Hornbæk (1); 29, Kraków Zwierzyniec (1); 30, Kostenki 4 (1); 31, Kraków Spadzista C (1); 32, Moravany, Podkovica (2); 33, Sokmarskaya Cave (1); 34, Opole–Groszowice (1); 35, Mokrzec, Wisłoka River (1); 36, Avdeevo (5); 37, Borschevo 5 (2); 38, Lublin-Zemborzyce (1); 39, Tsuren, Prut River (1); 40, Nilsiä (1); 41, Hüntwagen (1); 42, Tarnobrzeg-Machów (1); 43, Kostenki 8 (2); 44, Jenerálka (1); 45, Krasnoborsk (1); 46, Kraków Spadzista, trench III (1); 47, Mezin (1); 48, Zaraysk (4); 49, Gagarino (2); 50, Myrup Banke (1); 51, Wil (2); 52, Tuřenų Quarry (1); 53, Villafranche-sur-Saône (1); 54, Sagaidak 1 (1); 55, Kostenki 6 (1); 56, Rond-du-Barry (1); 57, Pushkari 1 (4); 58, Kostenki 5 (2); 59, Molodova (1); 60, Valea Morilor, Chişinău (3); 61, Obolonnya (1); 62, Kostenki 18 (1); 63, Nowy Targ (1); 64, Mellikon (1); 65, Kostenki 11 (1); 66, Wildscheuer Cave (1); 67, Lukhnevschina (1); 68, Dresden (1); 69, Castlepook Cave (1); 70, Wróblowa-Ujazd (1); 71, Binningen (2); 72, Minsk (1); 73, Spytihněv-Duchonce (1); 74, Højballegård (1); 75, Kostenki 14 (2); 76, Hardtwald (1); 77, Böttstein (1); 79, Novgorod-Severskii (1); 80, Kostenki 3 (1); 81, Cueto de la Mina E (1).

Figure 4 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland interstadial 2 (GI-2). Sites (number of dates in parentheses): 49, Gagarino (1); 62, Kostenki 18 (1); 78, Arrie (1); 82, English Channel (1); 83, Mezhirichi (3); 84, Kiev-Kirilovskaya (1); 85, Radomyshl (1).
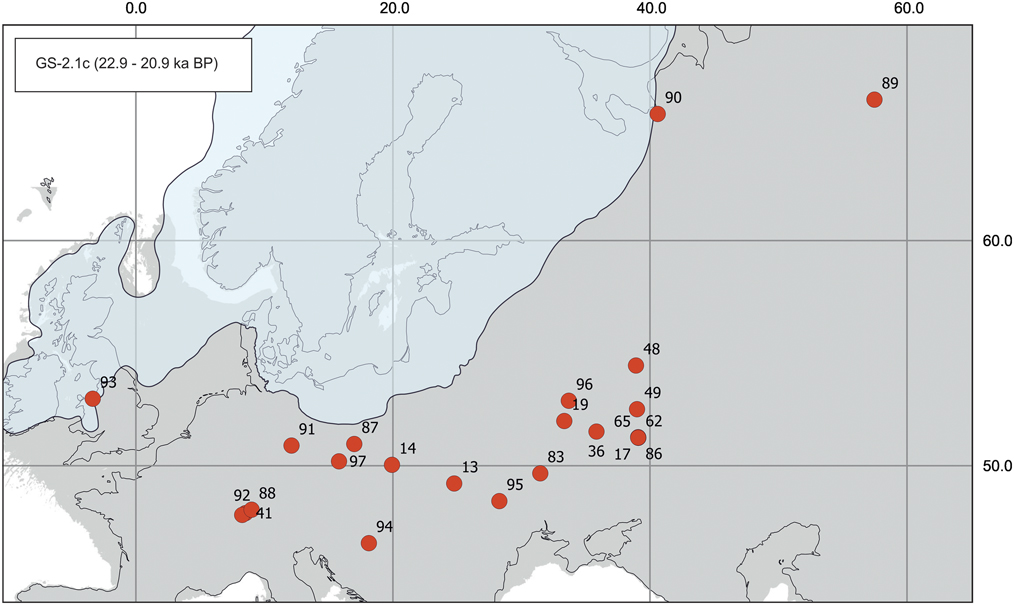
Figure 5 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland stadial 2.1c (GS-2.1c). Sites (number of dates in parentheses): 13, Halych I (1); 14, Kraków Spadzista C2 (1); 17, Kostenki 1 (1); 19, Pogon (1); 36, Avdeevo (1); 41, Hüntwagen (2); 48, Zaraysk (1); 49, Gagarino (1); 62, Kostenki 18 (1); 65, Kostenki 11 (1); 83, Mezhirichi (1); 86, Kostenki 19 (1); 87, Wrocław–Oporów (1); 88, Markelfingen (1); 89, Byzovaya (1); 90, Archangelsk (1); 91, Breitenbach B (1); 92, Turgi-Geeling (1); 93, Cae Gwyn Cave (1); 94, Zók (1); 95, Cosăuţi 1 (1); 96, Eliseevichi 1 (1); 97, Svobodné Dvory (1).
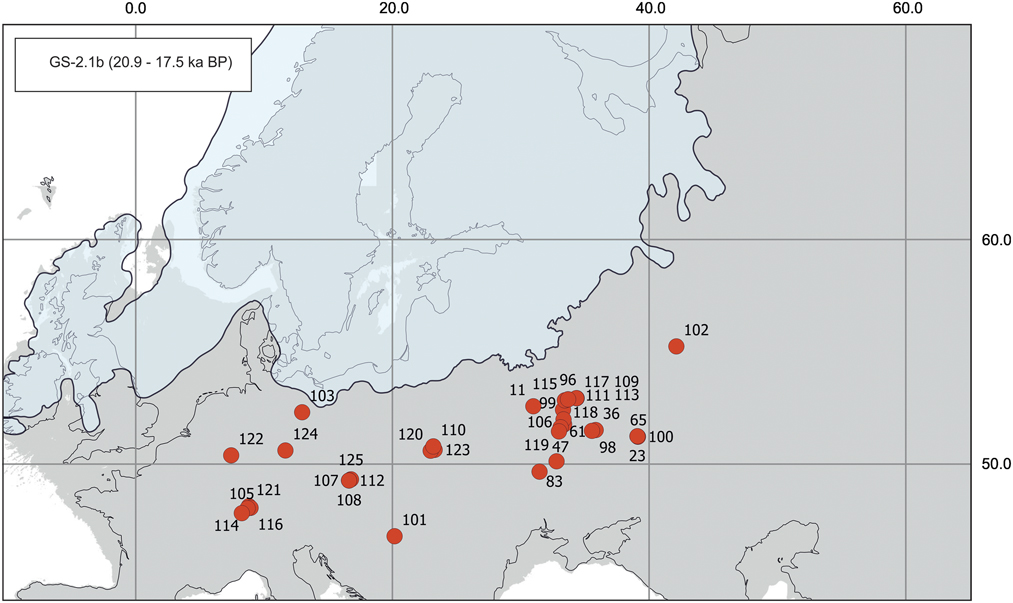
Figure 6 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland stadial 2.1b (GS-2.1b). Sites (number of dates in parentheses): 11, Berdyzh (1); 23, Kostenki 2 (1); 36, Avdeevo (3); 47, Mezin (1); 61, Obolonnya (1); 65, Kostenki 11(1); 83, Mezhirichi (9); 96, Eliseevichi 1 (3); 98, Pieny (1); 99, Pushkari IX (2); 100, Borschevo 1 (4); 101, Szeget–Őthalom (1); 102, Karacharovo (4); 103, Wustermark 22 (1); 105, Gebensdorf (1); 106, Yudinovo (4); 107, Brno-Štýřice III (Videňská St.–Hospital grounds) (1); 108, Brno-Štýřice III (Videňská St.) (1); 109, Eliseevichi 2 (1); 110, Izbica (1); 111, European Russia (center) (1); 112, Brno-Štýřice III (2); 113, Timonovka 2 (1); 114, Kesslerloch Cave (1); 115, Bryansk Region (5); 116, Steisslingen (1); 117, Timonovka 1 (2); 118, Chulatovo 1 (1); 119, Gontsy (6); 120, Kawęczyn (1); 121, Engen (1); 122, Gönnersdorf (2); 123, Zamość (1); 124, Kniegrotte (1); 125, Pékarna (1).

Figure 7 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland stadial 2.1a (GS-2.1a). Sites (number of dates in parentheses): 36, Avdeevo (1); 83, Mezhirichi (2); 89, Byzovaya (1); 96, Eliseevichi 1 (2); 105, Gebensdorf (1); 106, Yudinovo (3); 114, Kesslerloch Cave (1); 119, Gontsy (4); 122, Gönnersdorf (1); 126, Shatrishchi I (1); 127, Buzhanka 2 (1); 128, Zbranki (1); 129, La Croze (1); 130, Suponevo (3); 131, Brno-Štýřice III (Kamenná St.) (1); 132, Semenivka 2 (1); 133, Dzierżysław (6); 134, Lublin-Kalinowszczyzna (2); 135, Oelknitz (1); 136, European Russia, center (1); 137, Risch-Rotkreuz (1); 138, Sevsk (2); 139, Obukhiv (1); 140, Bzianka (1); 141, Jiesia River (1); 142, Praz-Rodet (1); 143, La Colombière Rock-Shelter (1); 144, Uster-Oberuster (1); 145, Csajág(1); 146, Lockarp (1); 147, Jičin (1); 148, Kopachiv (1); 149, Wilczyce (1); 150, La Grotte des Romains (1); 151, Rucava (1); 152, Marolles-sur-Seine (1); 153, Dobranichivka (1); 154, Cherepovets (1).

Figure 8 (color online) Map of radiocarbon dates for Mammuthus primigenius from Greenland interstadial 1e (GI-1e). Sites (number of dates in parentheses): 106, Yudinovo (3); 117, Timonovka 1 (1); 155, Gough’s Cave (1); 156, Pin Hole Cave (1); 157, Mamontovaya Kurya (1); 158, Condover (4); 159, Robin Hood’s Cave (1); 160, Etiolles (1); 161, Shatrishchi 2 (1); 162, Verberie (1).

Figure 9 (color online) Distribution of directly dated mammoth remains in Europe during Marine Oxygen Isotope Stage 2 (MIS 2). (A) Spatiotemporal distribution of remains. (B) Generalized additive model explaining relative mammoth abundance during MIS 2 (degree of smoothing, k=15). (C) Number of dates (N) in the 0.2 ka bins. (D) NGRIP (North Greenland Ice Core Project) ice core δ18O record on the GICC05 time scale. GI, Greenland interstadial; GS, Greenland stadial.
We used generalized additive models (GAMs) implemented in R (R Development Core Team, 2016) using the “mgcv” package (Wood, Reference Wood2017) to describe temporal variation in the relative abundance of mammoths in relatively short interstadial (GI) intervals and much longer stadial (GS) intervals. For this purpose, we compared our selected 318 radiocarbon dates (ranging from 28,355 14C yr BP to 14,128 14C yr BP; Supplementary Table 2) with 3180 (i.e., 10 times the number of radiocarbon dates) random dates sampled from the uniform distribution of the same range. These random dates represent the null model under which mammoths occurred evenly (with random variation) over the study period. In the GAM (with binomial error distribution and logit link), type of date was the response variable (318 radiocarbon dates were coded as “1,” and 3180 random dates were coded as “0”), and time was the explanatory variable fitted with thin plate regression splines (Wood, Reference Wood2017). Splines allow for the determination of nonlinear fit directly from data (i.e., function does not have to be determined a priori), which in our case allowed us to test nonlinear trends of mammoth abundance over time (i.e., periods of higher and lower relative abundance). We considered splines of different complexity (fixed “k” parameter ranging from 3 to 30), but as all of them showed similar patterns (as well as spline fit without fixed “k” parameter), we presented the one (Fig. 9) that seemed most relevant, and showed all others in Supplementary Figure 1.
RESULTS
In the analysis, as many direct mammoth dates as possible were compared. The mammoth record presented here was compiled by the authors from published papers, and only a few are new or were mentioned in the literature without detailed information (Table 1). We gathered 373 radiocarbon dates (Supplementary Table 3); however, as expected, the quality of radiocarbon dates was not equal from case to case, and the final number of dates used was 318 (including combined dates) from 162 localities (Supplementary Table 2). We rejected 43 dates (Supplementary Table 3), but a few controversial dates were retained on maps for further discussion (e.g., Fig. 4).
GS-4 interval (28.6–27.8 ka)
In this short cold interval, mammoths were not numerous but present in the whole European Plain (Fig. 1). There are dated records ranging from the East European Plain, for example, in Kostenki 12 (Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997), Sungir (Sulerzhitsky et al., Reference Sulerzhitsky, Pettitt and Bader2000), and Berdyzh in Belarus (Kalechits, Reference Kalechits2013), through southern Poland in Jaksice II (Wilczyński et al., Reference Wilczyński, Wojtal, Robličkova and Oliva2015b), Kraków Spadzista (Wojtal and Sobczyk, Reference Wojtal and Sobczyk2005; Wojtal and Wilczyński, Reference Wojtal and Wilczyński2015a), and Zastruże (Wiśniewski et al., Reference Wiśniewski, Wojtal, Krzemińska, Zych, Przybylski, Badura and Ciszek2009), and farther west to Copenhagen (Denmark) (Brace, Reference Brace2011), Goyet Cave (Belgium) (Barnes et al., Reference Barnes, Shapiro, Lister, Kuznetsova, Sher, Guthrie and Thomas2007), and Paviland Cave (United Kingdom) (Jacobi and Higham, Reference Jacobi and Higham2008). Mammoths are also recorded farther south in Dolní Vĕstonice (Moravia, Czech Republic) (Fig. 1). The chronological record is continuous, and the number of dated specimens is 21, despite the fact that this climatic event lasted only ~0.8 ka.
GI-3 interval (27.8–27.5 ka)
It is not obvious whether a very short last MIS 3 warming reduced the range of the mammoth in western Europe, because the observed pattern could also be attributable to lack of sampling (Fig. 2). Confirmed finds (n=13) are restricted to East European Plain sites in Kostenki 1, Khotylevo 2 (Gavrilov et al., Reference Gavrilov, Voskresenskaya, Maschenko and Douka2015), Pogon (Barns et al., Reference Barnes, Shapiro, Lister, Kuznetsova, Sher, Guthrie and Thomas2007), and Berdyzh (Arslanov et al., Reference Arslanov, Voznyachuk, Kalechits and Kolesnikov1972). The dated premolar from Helsinki (Töölö) (Fig. 2, site 21) was probably deposited in a large ice-free area in the southeastern part of Fennoscandia (Ukkonen et al., Reference Ukkonen, Lunkka, Jungner and Donner1999). Another concentration of sites (Świlcza, Żywiec, Jaksice II, and Kraków Spadzista; Arppe and Karhu, Reference Arppe and Karhu2010; Nadachowski et al., Reference Nadachowski, Lipecki, Wojtal and Miękina2011; Wilczyński et al., Reference Wilczyński, Wojtal, Łanczont, Mroczek, Sobieraj and Fedorowicz2015a) is situated mainly along the Vistula River valley, in south Poland.
GS-3 interval (27.5–23.5 ka)
The number of dated records during the relatively long-lasting GS-3 (4.2 ka) is 115 (Fig. 3), but chronology is not continuous in all parts of the continent. Although data are still limited, there is enough evidence confirming that during the first millennium of GS-3 (~27.5–26.5 ka), the population of M. primigenius was stable, especially in the eastern part of the European Plain (Russia, Ukraine, and Poland). One dated molar from Nilsiä (Fig. 3, site 40) was probably originally deposited in a large ice-free area in Finland (Ukkonen et al., Reference Ukkonen, Lunkka, Jungner and Donner1999). Mammoths were able to expand to the south by following river valleys or crossing the low mountain passes, as evidenced from Moravany–Podkovica (Slovakia) (Verpoorte, Reference Verpoorte2002), Jarošov I in southern Moravia (Czech Republic) (Oliva, Reference Oliva2007), and Willendorf II in Krems (Lower Austria), in the Danube River valley (Verpoorte, Reference Verpoorte2001). Farther westward, records are documented from Hornbæk in Denmark (Aaris-Sørensen, Reference Aaris-Sørensen2009) and perhaps Pámanes (Cantabria, north Spain) (Álvarez-Lao and Garcia, Reference Álvarez-Lao and García2012). Two radiocarbon dates on mammoth molars were published from the same mammoth mandible from Pámanes, one older (median 27.3 ka, recent calibration) and another younger (median 25.6 ka, recent calibration) (Álvarez-Lao and Garcia, Reference Álvarez-Lao and García2010, Reference Álvarez-Lao and García2012; Álvarez-Lao, D.J., personal communication, 2017). The reliability of the first date can be questioned because no samples of similar age are known from France and Germany. On the other hand one can assume that the Pámanes specimen belongs to the Iberian relict population of Mammuthus present in Cantabria earlier, during MIS 3 (e.g., in Labeko Koba; Álvarez-Lao and Garcia, Reference Álvarez-Lao and García2012). We accepted the later date in our analysis. During the next three millennia (between ~26.5 ka and 23.5 ka), mammoths expanded farther west and north through Germany (Binningen, Hartwald: De Jong et al., Reference De Jong, de Graaff, Seijmonsbergen and Böhm2011) to England (Paviland Cave: Aldhouse-Green and Pettitt, Reference Aldhouse-Green and Pettitt1998; but see Jacobi and Higham, Reference Jacobi and Higham2008) and reached most probably even Ireland (Castlepook Cave: Woodman et al., Reference Woodman, McCarthy and Monaghan1997). Southwest expansion took place using mainly migration corridors in the northern outskirts of the Alps via Switzerland (e.g., Hünwagen, Wil, Mellikon: Kock et al., Reference Kock, Huggenberger, Preusser, Rentzel and Wetzel2009; Huber and Reinhard, Reference Huber and Reinhard2016) and Massif Central (e.g., Rond-du-Bary: Raynal et al., Reference Raynal, Lafarge, Rémy, Delvigne, Guadelli, Costamagno and Le Gall2014) to Cantabria (Pámanes, Cueto de la Mina: Álvarez-Lao and Garcia, Reference Álvarez-Lao and García2012). Wide distribution of mammoths during that time is accompanied by the decline in number of records with a local minimum around 26.5 ka BP (Fig. 9B and C). It is not clear whether this reflects a real decline in population density. Towards the end of GS-3, after a short-term increase in the number of dated mammoth finds, during the last millennium between ~24.3 ka and 23.5 ka, a gradual decrease is observed in central Europe, especially in Poland (Nadachowski et al., Reference Nadachowski, Lipecki, Wojtal and Miękina2011; Baca et al., Reference Baca, Nadachowski, Lipecki, Mackiewicz, Marciszak, Popović, Socha, Stefaniak and Wojtal2017). At the same time, the number of dates in western and eastern Europe remains almost the same. Although mammoths reached Cantabria and Ireland during GS-3, a gradual deterioration of the climatic conditions at the beginning of MIS 2 probably in general favored gradual reduction of the mammoth’s population in Europe.
GI-2 interval (23.5–22.9 ka)
There are only seven direct dates with medians in this period (Fig. 4). It is possible that at least five dates are erroneous because of insufficient laboratory procedures or sample impurities. The first one from Arrie, Sweden, is a combined date (Supplementary Table 3) and can be questioned because of the partial dissolution of the collagen and the possible contamination by preservatives (for further details, see Berglund et al., Reference Berglund, Håkansson and Lagerlund1976). Another date obtained on a mammoth tooth from the English Channel is a conventional date (Delibrias et al, Reference Delibrias, Guillier and Labeyrie1971) and is also uncertain. Five dates are from the western part of the East European Plain: Radomyshl (Gowlett et al., Reference Gowlett, Hedges, Law and Perry1987) and Kiev-Kirillovskaya (Gowlett et al., Reference Gowlett, Hedges, Law and Perry1987) are both AMS dates, whereas three others from Mezhirichi (Soffer, Reference Soffer1985), Kostenki 18 (Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997), and Gagarino (Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997) are conventional dates. Therefore, there is also a possibility that mammoths entirely withdrew from most of Europe or survived only in some parts of the East European Plain. This is supported by the GAM analysis where the relative decrease of mammoth remains is visible around GI-2 (Fig. 9), despite that all seven dates were used in the analysis.
GS-2.1c interval (22.9–20.9 ka)
During the first part of the longest stadial interval of the last glacial period, mammoths were present in a major part of the former range occupied during GS-3, except in southern France and northern Spain (Fig. 5). However, there is an important decrease in the number of records in Europe (total n=24) in comparison with GS-3 (Fig. 9B and C). For the first time during MIS 2, mammoths were able to migrate south, to the Pannonian Basin, confirmed by a record from Zók, Hungary (Konrád et al., Reference Konrád, Kovács, Halász, Sebe and Pálffy2010). At the end of this interval, ~21.4 ka, mammoths probably withdrew from all of Europe for the next ~0.5 ka. After this event, there are only two records of Mammuthus in Eliseevichi 1, Desna River valley, Russia (Kurenkova, Reference Kurenkova1978), and Svobodné Dvory, Czech Republic (Šída et al., Reference Šída, Nývltová-Fišáková and Verpoorte2006), close to the termination of the GS-2.1c interval. Two AMS dates from Svobodné Dvory have been obtained from the same piece of a mammoth tusk. A sample that contained too little collagen for dating, in the opinion of Šída et al. (Reference Šída, Nývltová-Fišáková and Verpoorte2006), returned a date of 17,400±80 14C yr BP (GdA-460); a second date, 17,340±130 14C yr BP (GrA-29390) is also not reliable, because of low carbon content and the possibility of contamination from preservative.
GS-2.1b interval (20.9–17.5 ka)
In the next two millennia (~20.9 ka to 19.0 ka), the maximum extent of the Scandinavian Ice Sheet had a profound impact on mammoth populations in Europe (Fig. 6). Earlier published comparisons of direct Mammuthus dates from Europe suggested an apparent absence of mammoths in the interval ~22.0–18.0 ka, or at least between ~21.0 and 19.0 ka (Stuart et al., Reference Stuart, Kosintsev, Higham and Lister2004; Lister and Stuart, Reference Lister and Stuart2008; Nadachowski et al., Reference Nadachowski, Lipecki, Wojtal and Miękina2011; Ukkonen et al., Reference Ukkonen, Aaris-Sørensen, Arppe, Clark, Daugnora, Lister and Lõugas2011). Our analysis shows a continuous reduction of available dates after 21.4 ka (Supplementary Table 2) in all parts of the continent. There are indications of gaps in the pattern of dates in certain areas. Available records between 20.9 ka and 20.0 ka (altogether seven conventional dates) come from sites in the central East European Plain along the Don River valley (Kostenki, Borshchevo) and the Desna River valley and its tributaries (Avdeevo, Eliseevichi, Pushkari, Pieny) (Svezhentsev, Reference Svezhentsev1993; Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997; Sulerzhitsky, Reference Sulerzhitsky2004; Khlopachev, Reference Khlopachev2011). All dates from these sites, obtained mainly in the 1980s or before, have not been AMS redated, and their quality may be questioned. Therefore, the possibility that mammoths were absent in Europe at that time should be reconsidered. In subsequent millennia (between 20.0 ka and 19.0 ka), mammoths withdrew from Europe and probably returned only 19.2–19.0 ka; this suggestion is supported by the lack of dates for almost 0.8 ka (Supplementary Table 2). Conventional dates around 19.2–19.0 ka are known from Karacharovo, East European Plain (Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997), and Szeget-Őthalom in Hungary (Sümegi and Hertelendi, Reference Sümegi and Hertelendi1998). AMS 14C dates are available from Wustermark 22, eastern Germany, where prehistoric ivory fishhooks have been described (Gramsch et al., Reference Gramsch, Beran, Hanik and Sommer2013), and from Gebenstorf, Switzerland, where a mammoth tooth root was directly dated (Huber and Reinhard, Reference Huber and Reinhard2016). Since ~19.0 ka, mammoth finds sharply increase in eastern and central Europe, and around the end of GS-2.1b (~17.5 ka), the number of dates is the largest in the whole of MIS 2.
GS-2.1a interval (17.5–14.7 ka)
The post-LGM time was characterized by intensive deglaciation processes. Mammoths were still numerous in eastern and central Europe, reaching in the west at least to France, as confirmed by AMS dates from Marolles-sur-Seine (Barnes et al., Reference Barnes, Shapiro, Lister, Kuznetsova, Sher, Guthrie and Thomas2007) and La Grotte des Romains (Oberlin and Pion, Reference Oberlin and Pion2009) (Fig. 7). Northward migrations are indicated in our record after ~16.8 ka by dates from Lockarp, Sweden (Kjær et al., Reference Kjær, Lagerlund, Adrielsson, Thomas, Murray and Sandgren2006); Jiesia River, Lithuania (Arppe and Karhu, Reference Arppe and Karhu2010); and Rucava, Latvia (Arppe and Karhu, Reference Arppe and Karhu2010). After ~15.6 ka, the number of dates decreases, especially in central Europe, and before the GI-1e warming, mammoths withdrew from many places in Europe, except in the northern and eastern areas.
GI-1e interval (14.7–14.1 ka)
At the beginning of the late-glacial period, an abrupt GI-1e warming (Steffensen et al., Reference Steffensen, Andersen, Bigler, Clausen, Dahl-Jensen, Fischer and Goto-Azuma2008) reduced the size of the European mammoth population and fragmented mammoth distribution (Fig. 8). The first group of dates is concentrated in the northwestern part of the continent in England and northern France (Hedges et al., Reference Hedges, Housley, Law and Bronk Ramsey1989; Stuart et al., Reference Stuart, Sulerzhitsky, Orlova, Kuzmin and Lister2002; Lister, Reference Lister2009), and the second group is in the central part of the East European Plain, Russia (Sinitsyn et al., Reference Sinitsyn, Praslov, Svezhentsev and Sulerzhitsky1997; Khlopachev, Reference Khlopachev2015). Mammoths were probably still present farther northeast, as documented by a single date from Mamontovaya Kurya (Svendsen and Pavlov, Reference Svendsen and Pavlov2003). However, we note that all dates from Russia are conventional and have not been confirmed by more AMS redating.
DISCUSSION
The woolly mammoth (M. primigenius) is one of most spectacular megafaunal species that became extinct in Europe, Asia, and North America during the Late Quaternary (Stuart, Reference Stuart2015). It was the largest herbivore in a cold and arid steppe-tundra ecosystem that covered a large part of the Northern Hemisphere (Guthrie, Reference Guthrie1990). Being well adapted to this environment, the mammoth was widespread in almost all the northern Holarctic from western Europe to North America (Kahlke, Reference Kahlke2015); however, the species distribution varied through time because of population fluctuations, expansions and reductions of range, and population replacements (Palkopoulou et al., Reference Palkopoulou, Dalén, Lister, Vartanyan, Sablin, Sher and Nyström Edmark2013; Baca et al., Reference Baca, Nadachowski, Lipecki, Mackiewicz, Marciszak, Popović, Socha, Stefaniak and Wojtal2017; Chang et al., Reference Chang, Knapp, Enk, Lippold, Kircher, Lister and MacPhee2017; Fellows Yates et al., Reference Fellows Yates, Drucker, Reiter, Heumos, Welker, Münzel and Wojtal2017; Puzachenko et al., Reference Puzachenko, Markova, Kosintsev, van Kolfschoten, van der Plicht, Kuznetsova, Tikhonov, Ponomarev, Kuitems and Bachura2017).
Europe is an important region for the study of migrations and extinctions, not only because of the abundance of radiocarbon dates made directly on mammoth remains, but also because of the large amount of archaeological and environmental data. In the late Pleistocene, the most extensive migrations of herbivores in Europe took place mostly in the Great European Plain north of the east–west axis of the higher mountain ranges—namely, the Carpathians, Sudetes, Alps, and Pyrenees (Lorenzen et al., Reference Lorenzen, Nogués-Bravo, Orlando, Weinstock, Binladen, Marske and Ugan2011; Stuart and Lister, Reference Stuart and Lister2012; Sommer et al., Reference Sommer, Kalbe, Ekström, Benecke and Liljegren2014; Nadachowski et al., Reference Nadachowski, Lipecki, Ratajczak, Stefaniak and Wojtal2016). During colder phases, M. primigenius expanded southward of the geographic mountain barriers, most probably along river valleys, such as the Pannonian Basin (Kovács, Reference Kovács2012) and the Sava River valley in Croatia (Mauch Lenardić, Reference Mauch Lenardić2012). On the Apennine Peninsula, mammoths reached Apulia in southern Italy, possibly during MIS 2 (Rustioni et al., Reference Rustioni, Ferretti, Mazza, Pavia and Varola2003); even the south of the Iberian Peninsula was occupied during stadial intervals of MIS 3 (Álvarez-Lao and García, Reference Álvarez-Lao and García2010, Reference Álvarez-Lao and García2012).
MIS 3
The middle part of the last glacial period, known as MIS 3, was characterized by variable climate with a series of longer and/or shorter and milder D/O (Dansgaard/Oeschger) events (Van Andel and Davis, Reference Van Andel and Davis2003). The spatial distribution record of mammoth remains at the end of MIS 3, between ~32.5 ka and 27.5 ka (from GS-5 to GI-3), shows that the mammoth was widespread in Europe (Ukkonen et al., Reference Ukkonen, Aaris-Sørensen, Arppe, Clark, Daugnora, Lister and Lõugas2011). The high density of mammoth populations during the relatively long cold stage GS-5 is not surprising. During the climatically variable end of MIS 3 (two short warmings, GI-4 and GI-3, interrupted by GS-4), the large number of mammoth remains was maintained as confirmed by data from European Gravettian sites such as Khotylevo (Gavrilov et al., Reference Gavrilov, Voskresenskaya, Maschenko and Douka2015) and Kraków Spadzista (Wojtal and Sobczyk, Reference Wojtal and Sobczyk2005; Wilczyński et al., Reference Wilczyński, Wojtal and Sobczyk2012). Gravettian hunter-gatherers specialized in hunting mammoths and accumulated hundreds of bones of Mammuthus close to occupation areas (Wojtal and Wilczyński, Reference Wojtal and Wilczyński2015a). This pattern probably partly correlates with the real size of the mammoth population in the European Plains at that time, but it also probably partly reflects the intensity of specialized human hunting.
Our mammoth record from the end of MIS 3, including the cooler GS-4 and warming GI-3, does not show a distinct difference in the number of dates (Figs. 1 and 2). However, analysis of the geographic distribution of records does reveal a visible contraction in the range during GI-3 in some parts of Europe. This reduction in dates in western Europe may have significance in terms of migration processes connected with the impact of climate change.
MIS 2
The longest cold stage in the last glacial period lasted ~12.8 ka (GS-3, GI-2, and GS-2), and within MIS 2, the LGM is traditionally placed, although the definition of LGM is not consistent (for discussion, see Mix et al., Reference Mix, Bard and Schneider2001; Clark et al., Reference Clark, Dyke, Shakun, Carlson, Clark, Wohlfarth, Mitrovica, Hostetler and McCabe2009; Hughes and Gibbard, Reference Hughes and Gibbard2015). A relatively large number of dates from MIS 2 make it possible to construct detailed chronologies for mammoth distribution in space and time (Fig. 9). During MIS 2, migration processes or range abandonments seemed to have been highly dynamic, not only in Europe, but also in Asia, during the Sartan stadial interval, the equivalent of MIS 2 in Arctic Siberia (Sulerzhitsky, Reference Sulerzhitsky1995; Nikolskiy et al., Reference Nikolskiy, Sulerzhitsky and Pitulko2011). This conclusion is in agreement with Nadachowski et al. (Reference Nadachowski, Lipecki, Wojtal and Miękina2011) and Ukkonen et al. (Reference Ukkonen, Aaris-Sørensen, Arppe, Clark, Daugnora, Lister and Lõugas2011) who believed that local extinctions had occurred first of all in central and northern Europe.
During almost the entire GS-3 interval, M. primigenius was widespread over all of Europe (Figs. 3 and 9). The distribution and frequency of dates fluctuate, and a smaller number of dates occur between ~25.5 and 24.8 ka, correlating with a slight climatic amelioration. At the end of this interval (after 24.3 ka) in central Europe, the number of dates slightly decreases. In general, the second half of GS-3 seems to be an important time of faunal turnover. For instance, the last populations of the cave bear (Ursus spelaeus s.l.) in Europe are dated to this period (Bocherens et al., Reference Bocherens, Bridault, Drucker, Hofreiter, Münzel, Stiller and van der Plicht2014; Baca et al., Reference Baca, Popović, Stefaniak, Marciszak, Urbanowski, Nadachowski and Mackiewicz2016); on the other hand, the first large migrations of saiga antelope (Saiga tatarica) from eastern to central Europe in the last glacial period are also dated to this time (Nadachowski et al., Reference Nadachowski, Lipecki, Ratajczak, Stefaniak and Wojtal2016).
A short warming of GI-2, lasting only ~0.6 ka, accelerated a longer process of mammoth population demise, and thus an important change in Mammuthus distribution in Europe. Since the beginning of MIS 2, this included most probably the first interval showing a continent-wide lack of mammoth remains, indicating either withdrawal or significant reduction in Mammuthus population size (Figs. 4 and 9).
At the beginning of the last interval of MIS 2, the GS-2 interval, which lasted ~8.2 ka, mammoths returned to Europe for a short time but were not numerous. We confirmed a progressive reduction in their range from ~21.4 ka until 19.2 ka, as suggested by Lister and Stuart (Reference Lister and Stuart2008), Nadachowski et al. (Reference Nadachowski, Lipecki, Wojtal and Miękina2011), and Ukkonen et al. (Reference Ukkonen, Aaris-Sørensen, Arppe, Clark, Daugnora, Lister and Lõugas2011). The time span coincides with the maximum extent of the Scandinavian Ice Sheet in Europe (Marks, Reference Marks2012). We do not intend to repeat arguments presented by Ukkonen et al. (Reference Ukkonen, Aaris-Sørensen, Arppe, Clark, Daugnora, Lister and Lõugas2011); however, the harsh climate south of the Scandinavian Ice Sheet, which was extremely cold and dry, was undoubtedly responsible for the retreat of mammoths for a longer time from the continent.
At the beginning of deglaciation ~19.0 ka, mammoths soon reoccupied a wide area. Although M. primigenius was widespread over most of this time span, we found a progressive increase of dates only ~18.0–17.5 ka, in the middle part of GS-2 (Figs. 7 and 9). The extirpation of Mammuthus from Europe had begun ~15.7–15.4 ka, when the number of dates in the central part of the range dramatically decreased.
MIS 1
One thousand years later, after an abrupt warming at 14.7 ka (onset of GI-1e), a rapid reduction and fragmentation of mammoth range is suggested for a wide area of Europe except in the more oceanic northwestern parts—namely, Britain and northern France—and perhaps the central and northern East European Plain. However, this has not been confirmed by more recent AMS dating. A longer survival of M. primigenius in England and northern France seems surprising. One possible explanation is that the steppe-tundra ecosystem in northwestern Europe lasted longer and was relatively much larger because of the presence of a now inundated huge landmass referred to as Doggerland in the present-day North Sea (Coles, Reference Coles2000). During the GI-1 interval, Britain, Doggerland, and the northwestern part of continental Europe were still covered by open grassy parkland vegetation suitable for mammoth populations, in contrast to central Europe, which was already covered by pine forests (Brewer et al., Reference Brewer, Giesecke, Davis, Finsinger, Wolters, Binney and de Beaulieu2017).
Impact of humans on range dynamics of the woolly mammoth (M. primigenius) in Europe during MIS 2
The Elephantidae played a significant role in the lifeways of Paleolithic societies. The oldest signs of the exploitation of these large mammals are known from the Lower Paleolithic (Thieme and Veil, Reference Thieme and Veil1985; Haynes, Reference Haynes1991; Gaudzinski et al, Reference Gaudzinski, Turner, Anzidei, Álvarez-Fernández, Arroyo-Cabrales, Cinq-Mars and Dobosi2005; Yravedra et al., Reference Yravedra, Domínguez-Rodrigo, Santonja, Pérez-González and Panera2010; Ben-Dor et al., Reference Ben-Dor, Gopher, Hershkovitz and Barkai2011), but the most extensive exploitation of mammoths is visible at Upper Paleolithic sites in central and eastern Europe, especially during the emergence of the Gravettian and Epigravettian (Soffer, Reference Soffer1985, Reference Soffer1993; Iakovleva and Djindjian, Reference Iakovleva and Djindjian2005; Wojtal and Sobczyk, Reference Wojtal and Sobczyk2005; Brugère and Fontana, Reference Brugère and Fontana2009; Musil, Reference Musil2010; Wilczyński et al., Reference Wilczyński, Wojtal, Robličkova and Oliva2015b; Wojtal and Wilczyński, Reference Wojtal and Wilczyński2015b; Demay et al., Reference Demay, Belyaeva, Kulakovska, Patou-Mathis, Péan, Stupak, Vasil’ev, Otte and Noiret2016; Münzel et al., Reference Münzel, Wolf, Drucker and Conard2016). The time span of these two cultural units falls at the end of MIS 3 and throughout MIS 2 and is connected with important changes in mammoth range. During this period, assemblages containing several hundred thousand bones and teeth have been discovered in central Europe’s loess localities. One such location is Předmosti (Czech Republic), where it was estimated that mammoth remains belong to a minimum of 1000 individuals (Musil, Reference Musil2010). The complexity of the site and others like it (Dolní Věstonice and Milovice in the Czech Republic, Kraków Spadzista in Poland) makes it essential to be careful when explaining the origin and function of the bone accumulations. This issue has been of interest to many researchers from the beginning of scientific study of the sites. Some of them prefer to interpret mammal bones as intentional accumulations made by humans transporting bones from elsewhere in the landscape, whereas other specialists are inclined to see them as results of mammoth hunting or as places where bones from noncultural deaths had accumulated naturally and were reused by hunter-gatherers (Steenstrup, Reference Steenstrup1889; Wankel, Reference Wankel1890; Absolon, Reference Absolon1945; Klima, Reference Klima1963, Reference Klima1990; Kozłowski et al., Reference Kozłowski, Van Vliet, Sachse-Kozłowska, Kubiak and Zakrzewska1974; Haynes, Reference Haynes1991; Soffer, Reference Soffer1993; Musil, Reference Musil1997; Svoboda, Reference Svoboda2001; Svoboda et al., Reference Svoboda, Pean and Wojtal2005; Oliva, Reference Oliva2009). Any interpretation of these assemblages is complicated by the fact that the site locations were frequently reoccupied by Paleolithic hunter-gatherers over long stretches of time, and subsequent stays left features and artifacts mixed together in a single layer as palimpsest deposits. Based on evidence from central and eastern European sites, there is a conviction among several authors that the hunting of mammoths was an essential part of Paleolithic subsistence. This first hypothesis, more eagerly portrayed in the past, favored the theory of natural formation of mammoth bone accumulations or their creation as a result of humans transporting the bones. Nowadays, more and more evidence speaks for a hypothesis related to active hunting. It has some confirmation in finds that document active hunting of mammoths (Zenin et al., Reference Zenin, Leshchinskiy, Zolotarev, Grootes and Nadeau2006; Nikolskiy and Pitulko, Reference Nikolskiy and Pitulko2013), in isotopic studies of human diets (Bocherens et al., Reference Bocherens, Drucker, Germonpré, Lázničková-Galetová, Naito, Wissing, Brůžek and Oliva2015), in analysis of lithic artifacts associated with the mammoth remains, and in taphonomic studies (Wojtal and Sobczyk, Reference Wojtal and Sobczyk2005; Kufel-Diakowska et al., Reference Kufel-Diakowska, Wilczyński, Wojtal and Sobczyk2016). The evidence from Gravettian sites in different geographic regions and from different times indicates that mammoth-hunting was a recurring practice, and not an exceptional activity.
Two main hypotheses try to explain decreases and increases of mammoth populations: in one, population fluctuations resulted from human exploitation; in the other, mammoth populations were not significantly affected by human behavior/activity, and fluctuations in their populations were caused by environmental changes. We propose that Upper Paleolithic societies exploited mammoths much more intensively when the species was abundant, by adapting to the abundance, and during other periods of scarcity or disappearance of mammoths because of climatic influences, hunter-gatherers turned to other species (reindeer Rangifer tarandus, horses Equus). Plausible arguments can be made for this interpretation.
First of all, the creation of sites focused mainly on mammoths, such as Dolní Věstonice, Předmostí, Milovice, and Kraków Spadzista, precedes the GS-3 phase, dated ~27.5–23.3 ka, when evidence for the abundance of mammoths is seen in central Europe. In that case, human populations had little to no effect on mammoth populations throughout Europe. Moreover, whether or not mammoths were actively hunted during this period, when mammoth remains are often dominant in site assemblages, there also can be found localities where other hunted prey species prevail, such as Moravany Lopata II, Trenčianske Bohuslavice, and Jaksice II (Lipecki and Wojtal, Reference Lipecki and Wojtal1998; Vörös, Reference Vörös2000; Vlačiky, Reference Vlačiky2009; Wilczyński, Reference Wilczyński2015).
Second, the scarcity or disappearance of mammoths in Europe is positively correlated with the last glacial maximum, when human populations were also withdrawing from the northern part of central Europe (Verpoorte, Reference Verpoorte2004, Reference Verpoorte2009). We suggest that only during the times of the major decreases of the mammoth population, when its range became more fragmented during interstadial intervals and especially the late-glacial period, human activity affected local mammoth subpopulations. In other words, humans affected only small-sized mammoth populations and did not affect populations during phases of mammoth expansion, which has also been argued by other authors (Haynes, Reference Haynes1991; Lister and Stuart, Reference Lister and Stuart2008; Haynes and Klimowicz, Reference Haynes and Klimowicz2015).
Only in eastern Europe do mammoths dominate at Epigravettian sites, where architectural features made of mammoth bones occur in many sites; it should be noted that structures built with mammoth bones have been dated in this geographic region to the Middle Paleolithic, as in Molodova I (Chernysh, Reference Chernysh1982). We stress that these structures were made not only with bones from hunted animals, but also with bones that had been collected in the surrounding landscape (Soffer et al., Reference Soffer, Adovasio, Kornietz, Velichko, Gribchenko, Lenz and Suntsov1997; Iakovleva and Djindjian, Reference Iakovleva and Djindjian2005; Demay et al., Reference Demay, Péan and Patou-Mathis2012). Also to be emphasized is the fact that besides the sites abounding in mammoth remains, numerous other Gravettian and Epigravettian sites indicate that people depended on other mammal species for subsistence (Vörös, Reference Vörös2000; Iakovleva and Djindjian, Reference Iakovleva and Djindjian2005; Vlačiky, Reference Vlačiky2009; Musil, Reference Musil2010; Wojtal et al., Reference Wojtal, Wilczyński, Bocheński and Svoboda2012, Reference Wojtal, Wilczyński, Wertz and Svoboda2016; Demay et al., Reference Demay, Patou-Mathis and Kulakovska2015, Reference Demay, Belyaeva, Kulakovska, Patou-Mathis, Péan, Stupak, Vasil’ev, Otte and Noiret2016; Wilczyński et al., Reference Wilczyński, Wojtal, Łanczont, Mroczek, Sobieraj and Fedorowicz2015a, Reference Wilczyński, Wojtal, Robličkova and Oliva2015b).
Conclusions
The continuous range of M. primigenius in Europe was fragmented several times during MIS 2.
Temporal variation of the relative mammoth abundance was significantly nonlinear. During GS-3, GI-2, and GS-2.1c (between ca. 27.5 ka and 21.4 ka BP), the relative mammoth abundance was largely decreasing, but after ca. 19.2 ka, it started to increase and reached the maximum ca. 18.0–17.5 ka BP. This increase, however, was followed by another decrease and fragmentation of population after ca. 15.7 ka BP.
The greatest decline of population size took place during the maximum extent of the Scandinavian Ice Sheet, the most severe climatic period, leading to the disappearance of mammoths from western, northern, central, and probably eastern Europe, between ~21.4 and 19.2 ka BP.
In our opinion, there is no positive correlation between the distribution of European mammoths and human settlement. This is especially clear during the emergence and development of the Gravettian culture, which took place between ca. 31 and 22 ka BP, the period when fluctuation of the mammoth population is well visible. Humans affected local mammoth subpopulations only at the end of the last glacial period; we offer this as a hypothesis that should be further tested.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/qua.2018.54
ACKNOWLEDGMENTS
We acknowledge the analytical work of Tomasz Goslar and the staff of Poznań Radiocarbon Laboratory for 14C AMS dating. The KBN (State Committee for Scientific Research) 303 078 32/2589 project awarded to A. Nadachowski is thanked for financial support. M. Baca was supported by the Polish Science Center (grant 2015/19/D/NZ8/03878). The current studies were also partly supported by the National Science Centre (grant decisions nos. DEC-2013/09/D/HS3/04470 and DEC-2015/18/E/HS3/00178 awarded to J. Wilczyński). We acknowledge several local museums and other institutions (listed in the caption to Table 1) for permission to take samples for dating from specimens under their care. We are especially grateful for discussions and expertise to Diego J. Álvarez-Lao, María Soledad Corchón Rodrígez, Florian A. Fladerer, Teresa Madeyska, Piotr Wojtal, and Nataliya Zaretskaya. Special thanks to Gary Haynes for useful comments on an earlier draft of this manuscript, and Thijs van Kolfschoten and an anonymous reviewer for suggested revisions.












