


Introduction
Increased levels of negative mood has been related to a processing and recall bias for negative information (Teasdale & Fogarty, Reference Teasdale and Fogarty1979; Teasdale & Russell, Reference Teasdale and Russell1983). This processing bias has been shown in experimentally induced transient sadness in healthy individuals as well as in clinical depression. This mood-congruent heightened sensitivity towards negative stimuli and prolonged processing of emotionally negative information has been proposed to play an important role in the development and maintenance of unipolar depression. Cognitive theories of mood-biased processing, such as schema theory (Beck et al. Reference Beck, Rush, Shaw and Emery1979) and network theory of affect (Bower, Reference Bower1981) postulate that during a depressive state memory resources are bound to negative stimuli, which leads to an attenuation in memory capacity for incoming information of a positive emotional value (Ellis & Ashbrook, Reference Ellis, Ashbrook, Fiedler and Forgas1988).
Some corroborative evidence for these theories comes from behavioural studies investigating processing of negative stimuli (Gotlib & McCann, Reference Gotlib and McCann1984; Williams & Broadbent, Reference Williams and Broadbent1986; Gotlib & Cane, Reference Gotlib and Cane1987) and mood-congruent memory biases in depression (Lloyd & Lishman, Reference Lloyd and Lishman1975; Clark & Teasdale, Reference Clark and Teasdale1982; Bradley et al. Reference Bradley, Mogg and Millar1996).
The emotional Stroop task is a potentially useful tool to assess mood-congruent processing, by measuring the attentional bias towards negative stimuli. In this task affective words are presented in various colours, and subjects are required to name the colour of words rather than perform the automatic and overlearned response of reading the word itself. Interference effects arise when attention drawn to disorder-specific emotional expressions trigger memories of personal loss and failure. These disorder-specific self-schemas are then in conflict with a cognitive colour-naming instruction (MacLeod, Reference MacLeod1991; Segal et al. Reference Segal, Gemar, Truchon, Guirguis and Horowitz1995). Increased latencies are the result of this process.
A number of behavioural studies of emotional Stroop paradigms have observed these interference effects in depression (Williams & Broadbent, Reference Williams and Broadbent1986; Gotlib & Cane, Reference Gotlib and Cane1987; Segal et al. Reference Segal, Gemar, Truchon, Guirguis and Horowitz1995; Kerr et al. Reference Kerr, Scott and Phillips2005). Negative, positive and neutral words have been used to investigate, whether these interferences are elicited simply through the affect component rather than the negative information per se. Although a few studies report interference to affective material in general (Taylor & John, Reference Taylor and John2004), the majority of reports support the notion that depressed individuals display a memory bias for negative material centred around loss and failure (Watkins et al. Reference Watkins, Vache, Verney, Muller and Mathews1996; Neshat-Doost et al. Reference Neshat-Doost, Taghavi, Moradi, Yule and Dalgleish1998; Dudley et al. Reference Dudley, O'Brien, Barnett, McGuckin and Britton2002; Koster et al. Reference Koster, De, Goeleven, Franck and Crombez2005).
Neuroimaging studies suggest that the anterior cingulate cortex (ACC) plays a major role in Stroop task performance (Pardo et al. Reference Pardo, Pardo, Janer and Raichle1990; Bench et al. Reference Bench, Frith, Grasby, Friston, Paulesu, Frackowiak and Dolan1993; Taylor et al. Reference Taylor, Kornblum, Minoshima, Oliver and Koeppe1994, Reference Taylor, Kornblum, Lauber, Minoshima and Koeppe1997; George et al. Reference George, Ketter, Parekh, Rosinsky, Ring and Casey1994; Carter et al. Reference Carter, Mintun and Cohen1995; Bush et al. Reference Bush, Whalen, Rosen, Jenike, McInerney and Rauch1998; Peterson et al. Reference Peterson, Skudlarski, Gatenby, Zhang, Anderson and Gore1999; Banich et al. Reference Banich, Milham, Atchley, Cohen, Webb, Wszalek, Kramer, Liang, Wright, Shenker and Magin2000; Mead et al. Reference Mead, Mayer, Bobholz, Woodley, Cunningham, Hammeke and Rao2002; Gruber et al. Reference Gruber, Rogowska, Holcomb, Soraci and Yurgelun-Todd2002; Langenecker et al. Reference Langenecker, Nielson and Rao2004; Kerns et al. Reference Kerns, Cohen, MacDonald, Cho, Stenger and Carter2004). The ACC is at the core of organizing endogenous, individual-driven control processes and exogenous control processes, that are influenced by environmental distractions (Posner, Reference Posner1980). It is implicated in error monitoring and attentional processes (Paus, Reference Paus2001), coordinating cognitive control and conflict processes experienced during incompatible conditions (Carter et al. Reference Carter, Macdonald, Botvinick, Ross, Stenger, Noll and Cohen2000), and it is further involved in the elicitation of autonomic responses during cognitive challenges (Critchley et al. Reference Critchley, Wiens, Rotshtein, Ohman and Dolan2004).
In healthy individuals, during performance of an emotional Stroop task Whalen and colleagues (Reference Whalen, Bush, McNally, Wilhelm, McInerney, Jenike and Rauch1998) observed greater neural activity in the ACC with negative words and in absence of a behavioural interference effect. In individuals with a mood disorder, George and colleagues (Reference George, Ketter, Parekh, Rosinsky, Ring, Pazzaglia, Marangell, Callahan and Post1997) found greater latencies during a sad word condition relative to healthy controls but did not observe any significant differences in their neural activity. However, the patient group had a number of diagnoses and their affective state varied from euthymic to moderate depression. In individuals with bipolar disorder during a euthymic state, Malhi and colleagues (Reference Malhi, Lagopoulos, Sachdev, Ivanovski and Shnier2005) reported an emotional Stroop effect and increased ACC engagement, although all patients were taking medication.
A further, related, method of assessing a mood-influenced bias is through emotional go/no-go tasks. Sets of target and distractor words are presented and participants are instructed to respond to target words immediately by pressing a corresponding button. In these paradigms low latencies are a direct measure of a mood-congruent bias (Murphy et al. Reference Murphy, Sahakian, Rubinsztein, Michael, Rogers, Robbins and Paykel1999). These tasks have been used in behavioural as well as functional magnetic resonance imaging (fMRI) research (Murphy et al. Reference Murphy, Sahakian, Rubinsztein, Michael, Rogers, Robbins and Paykel1999; Elliott et al. Reference Elliott, Rubinsztein, Sahakian and Dolan2002; Erickson et al. Reference Erickson, Drevets, Clark, Cannon, Bain, Zarate, Charney and Sahakian2005). Increased activation during sad target words has been found for major depressive disorder (MDD) patients, relative to positive words and to controls, in the rostral ACC and medial prefrontal cortices (Elliott et al. Reference Elliott, Rubinsztein, Sahakian and Dolan2002), further supporting the role of the ACC in selective attention and mood-congruent processing.
In a fMRI study utilizing an original Stroop task, increased activation was found in the rostral ACC and the dorsolateral prefrontal cortex in unmedicated MDD patients, compared to controls (Wagner et al. Reference Wagner, Sinsel, Sobanski, Kohler, Marinou, Mentzel, Sauer and Schlosser2006). Dorsal ACC activation did not differ between groups.
Due to a high variability in sample characteristics and paradigm specifications findings of previous research are not always easy to interpret and compare. The present study aimed to control some factors, such as treatment effects and co-morbidity to investigate the relationship between attentional biases and mood-congruent processing in a group of medication-free patients with unipolar MDD. All MDD subjects were experiencing an acute depressive episode of moderate severity. We hypothesized that MDD patients would show an attentional bias in response to negative words which would be expressed in increased latencies during colour-naming emotional words. On a neural level, we expected an engagement of the ACC during processing of negative emotional words. As we did not use positive emotional stimuli our study cannot answer the question of effects of emotional arousal per se; however, as outlined above, emotional Stroop effects in depression have been shown to be mostly specific to negative stimuli.
Method and materials
Participants
After full explanation of the study procedures, all participants provided written, informed consent. The study was approved by the South London and Maudsley NHS Trust Research Ethics Committee.
Seventeen right-handed individuals (mean age 39.3 years, s.d.=9.4) meeting DSM-IV criteria for a diagnosis of unipolar major depression, acute depressive episode, were recruited through local newspaper advertisement. The diagnosis was obtained through the SCID-I, patient version (First et al. Reference First, Spitzer, Gibbon and Williams1997) and a clinical interview with a board-certified psychiatrist. Inclusion criteria included a score of ⩾18 on the 17-item Hamilton Depression Rating Scale (HAMD; Hamilton, Reference Hamilton1960). All participants were free of psychotropic medication for a minimum of 4 weeks at the time of inclusion into the study or a minimum of 8 weeks if the previous medication had been fluoxetine. Apart from psychotropic medication, one patient took trimethoprim, one received treatment for high cholesterol, one had an oestrogen patch, one patient had a nicotine patch and one patient sporadically used a nasal spray for hay fever. Exclusion criteria were co-morbidity with other Axis I disorders, current or past neurological disorders, history of neurological trauma resulting in loss of consciousness, reported substance or alcohol abuse within 2 months prior to study participation, and criteria that are contra-indicative for having a MRI scan. Seventeen right-handed volunteers, matched by age, gender and IQ (mean age 39.4 years, s.d.=9.2), without a personal or family history of psychiatric disorders, head injury resulting in unconsciousness, neurological disorder and current substance abuse were recruited through local advertisements. The ratio of smokers per group was 3:2, with three smokers in the patient group.
All participants completed the Beck Depression Inventory (BDI; Beck et al. Reference Beck, Ward, Mendelson, Mock and Erbaugh1961), and were assessed with the HAMD (Hamilton, Reference Hamilton1960) and the Wechsler Abbreviated Scale of Intelligence (WASI; Wechsler, Reference Wechsler1999) (Table 1).
Table 1. Demographics and mean scores of depression measures (HAMD, BDI)

MDD, Major depressive disorder; HAMD, Hamilton Depression Rating Scale; BDI, Beck Depression Inventory.
Experimental paradigm
Selection and assessment of words
A pilot study was first carried out to confirm the appropriateness of 40 negative and 40 neutral words and to assess their suitability in a clinical population. Fifteen healthy individuals (mean age 27.9 years, s.d.=7.8) and 15 medication-free individuals with mild depression (mean age 32.6, s.d.=10.9; mean BDI score 15.4, s.d.=2.9) participated in the pilot study. Eighty words were chosen from two lists of emotional and non-emotional words (John, Reference John1988; Bradley & Lang, Reference Bradley and Lang1999) and divided into two groups based on sad or neutral valence. Stimulus groups were matched for standardized lexical word frequency (Francis & Kucera, Reference Francis and Kucera1982), pronounceability (expressed in number of syllables) and word length. The John (Reference John1988) list contained word associations, and words were chosen that produced the lowest number of negative and happy associations for neutral words and the lowest number of happy associations for sad words. The Bradley & Lang (Reference Bradley and Lang1999) affective word norms were used to determine emotional valence and word frequency. Mean sad word frequency was 28.20 (s.d.=35.21) and mean word length was 6.75 (s.d.=1.81). Mean neutral word frequency was 33.95 (s.d.=47.03) and mean word length was 6.83 (s.d.=1.82). Independent-sample t tests confirmed that words did not differ significantly on frequency, pronounceability, or word lengths (all p>0.54), but did differ in emotional valence.
Forty negative and 40 neutral words (see Appendix) were presented on a laptop computer. Words were collated into blocks of eight words per emotional category, repeated five times whereby each word was presented only once with a presentation time of 700 ms per word. All words appeared on black background in red, blue, green or yellow colour, pseudo-randomized across the two valence categories.
Participants were informed that they would see words appearing individually on the screen, coloured in red, green, yellow or blue. They were instructed to name the colour of the word as quickly as possible, rather than reading the word. Reaction time recordings were done through onset of speech.
Reaction times were evaluated using a valence×group (2×2) repeated-measures ANOVA. The analysis revealed main effects of valence [F(1, 28)=21.98, p<0.001] and group [F(1, 28)=5.60, p=0.03] and a valence×group interaction effect [F(1, 28)=8.07, p=0.01]. This preliminary study confirmed an emotional interference effect for sad emotional stimuli in the patient group through prolonged latencies in colour naming of emotionally charged words.
fMRI experiment
The emotional Stroop task was projected onto a mirror inside the scanner. Head movement was restricted with foam padding and a Velcro strap across participant's forehead. Participants were required to name the colour of the presented word. Stimuli were then presented in a blocked design of ten alternating blocks of eight sad and eight neutral words, as described above. Volunteers' vocal responses were recorded by using a MRI compatible microphone and software that monitors the input to the computer soundcard. As soon as a vocal response was detected reaction times were measured. For each subject microphone settings were adjusted in order to account for individual differences in loudness of speaking voice. Each word was presented for 700 ms within a 2000 ms quiet period which allowed recording of their vocal responses in the relative absence of scanner noise. After 2000 ms (700 ms word presentation+1300 ms blank screen) image collection was carried out. The clustered/sparse image acquisition process has been used previously and has been found suitable for recording of vocal responses and minimizing the effects of head movement artefacts associated with verbal responses in healthy and patient groups (Fu et al. Reference Fu, Vythelingum, Brammer, Williams, Amaro, Andrew, Yaguez, van Haren, Matsumoto and McGuire2006). An auditory (.wav) file was recorded for each individual to ensure accuracy of response. Additionally, volunteers' responses were monitored by a researcher (M. T. M. or N. D. W.) through headphones and by pressing a corresponding coloured button on a button box.
Image acquisition
A 1.5 T General Electric Signa MR Imaging system (General Electric Medical Systems, Milwaukee, WI, USA) was used to acquire 84 T2-weighted echoplanar images depicting blood-oxygen-level dependent (BOLD) contrast. For each acquired volume 22 near-axial slices parallel to the intercommissural plane were collected over 2000 ms allowing for a silent period of 2000 ms (repetition time 4000 ms, echo time 40 ms, flip angle 90°, slice thickness 5 mm, interslice gap 0.5 mm, matrix size 64×64).
Statistical analysis
Behavioural data
MANOVA was carried out using group (MDD, controls) as independent variable to investigate differences between MDD patients and controls on psychometric measures (dependent variables: HAMD, BDI, WASI).
To test differences in reaction time and stimulus category between patients and controls, repeated-measures ANOVA was carried out using valence (sad, neutral) as within-group variable, group (MDD, controls) as between-group variable and reaction time (in ms) as dependent variable. Post-hoc t tests were carried out to follow-up significant main and interaction effects, two independent-sample t tests (sad and neutral) and two paired-sample t tests (MDD and controls). The α-level was set at 0.025 for each pair of tests.
fMRI data
Images were pre-processed and analysed using the Statistical Parametric Mapping (SPM2, Wellcome Department of Cognitive Neurology, London) software package on a Matlab platform. The 84 images functional time series was realigned, normalized into a standard stereotaxic space using a Montreal Neurological Institute EPI template and the coordinate system of Talairach & Tournoux (Reference Talairach and Tournoux1988) and smoothed using an 8 mm Gaussian kernel filter full-width-at-half-maximum to permit application of Gaussian random-field theory and to facilitate inter-subject averaging. The first four images were then discarded.
Group analysis was performed using a random-effects model that incorporated a two-stage hierarchical procedure. The first level analysis allowed computation of individual mean images that corresponded to the main contrasts of interest. The time series was modelled as a block-design with the temporal delay in BOLD response corrected for by a canonical haemodynamic response function. Contrasts were performed of the main effect of each affective state: negative>neutral and neutral>negative.
The significance of individual observations was tested in two second-level analyses by combining contrast images using one-sample t tests [p<0.05, family-wise error (FWE)] to assess groups individually and a two-sample t test. A two-sample t test examined differences between groups on key contrasts. A threshold of p<0.05, FWE was used to discuss brain areas without an a priori hypothesis. To discuss activation in the ACC, an uncorrected threshold of p<0.001 and small-volume correction (SVC) were applied. A general linear model was used to determine voxel-wise t statistics. This model estimates the error variance for each condition of interest across subjects, rather than across scans and therefore provides a stronger generalization to the population from which data are acquired. The t statistics were normalized to Z scores and significant clusters of activation were determined. To confirm validity of results acquired through an uncorrected p<0.001, region of interest (ROI) analysis using the Marsbar toolbox implemented in SPM2 was performed to identify significant activation levels in the ACC. Extracted data were analysed with SPSS software (SPSS Inc., Chicago, IL, USA) using an independent-sample t test. Furthermore, ROI data was used to investigate relations between latency and clinical measures (BDI, HAMD). Pearson correlations were calculated on activation levels, mean latencies as well as clinical variables.
Results
Behavioural data
Analysis of clinical measures revealed significant differences between the two groups in HAMD [F(1, 27)=1550.47, p<0.001] and BDI [F(1, 27)=128.69, p<0.001].
An ANOVA of reaction times of 30 datasets (four datasets could not be analysed due to problems with response recordings during neuroimaging) revealed significant main effects of valence [F(1, 28)=25.01, p⩽0.001], indicating that both groups showed an increased reaction time with sad words relative to neutral words, a main effect of group [F(1, 28)=0.02], indicating that depressed patients displayed increased latencies in response compared to controls, and an interaction effect of valence×group [F(1, 28)=10.27, p=0.03], suggesting a greater difference in latency between neutral and negative stimuli in patients than in controls (Fig. 1). Post-hoc analyses to explain the interaction effect revealed that the two groups differed significantly on negative (t=3.89, df=28, p=0.004) but not neutral (t=1.81, df=28, p=0.08) stimuli. Post-hoc analyses also revealed significant effects of valence in both the MDD (t=4.59, df=14, p<0.001) and the control (t=2.90, df=14, p=0.01) groups.
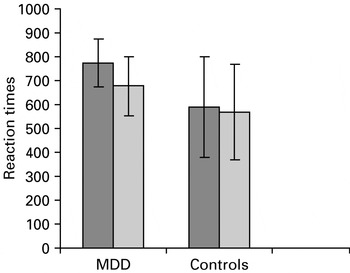
Fig. 1. Mean latencies and standard deviations for major depressive disorder (MDD) patients and controls during colour-naming negative (![]() ) and neutral (
) and neutral (![]() ) words during the fMRI experiment.
) words during the fMRI experiment.
For the purpose of completeness results are reported from the individual group analysis and between-group analysis. Results from between-group comparisons are discussed.
fMRI results
Effect of affect: negative words>neutral words
Controls
There was a significant bilateral increase in activation in the middle frontal/superior frontal gyrus (BA 10, BA 46: x=−30, y=62, z=14; p=0.003, Z=2.73).
MDD patients
In this group sad words were associated with a significant increase in activation in the left ACC (BA 32: x=−10, y=32, z=28; p=0.001, Z=4.05), right precuneus (BA 7: x=6, y=−60, z=42; p=0.01, Z=4.55), left middle temporal gyrus (BA 39: x=−52, y=−72, z=24; p=0.02, Z=3.89), left thalamus (x=−26, y=32, z=4; p=0.05, Z=3.68), right cerebellum (x=8, y=−46, z=−6; p=0.02, Z=3.60) and left middle frontal gyrus (BA 6: x=−26, y=10, z=58; p=0.01, Z=3.55).
Main effect of group: negative words>neutral words
MDD patients relative to controls activated the left ACC (BA 32: x=−5, y=30, z=24; p<0.000, Z=3.68; after SVC: p=0.004, Z=3.68) and right precuneus (BA 7: x=2, y=−60, z=44; p=0.02; FWE: Z=4.75) more strongly. Controls, relative to MDD patients, displayed no significant increase in activation in response to sad word stimuli (Fig. 2).

Fig. 2. Sagittal view of significantly increased activation in the left anterior cingulate cortex (ACC) (x=−5, y=30, z=24) in a between-group comparison, superimposed on a single-subject template. The graph on the right shows the correlation between activation in the ACC and latencies for colour-naming negative words in major depressive disorder patients (r=0.68, p=0.003).
Results from ROI analysis confirmed a significant difference between patients and controls on ACC activation (t=3.97, df=32, p⩽0.001).
Correlations with clinical measures and latencies
There was a significant correlation between mean signal change in the rostral ACC and latencies in response to negative emotional words (r=0.68, p=0.003) in MDD patients only (Fig. 2). We did not find significant correlations between negative word latencies and rostral ACC activation in healthy controls. Furthermore, correlations with clinical measures (HAMD, BDI) and brain activation in the rostral ACC did not produce significant results (r=1.39, p=0.98). This could be due to the small range of HAMD and BDI scores, since all patients were acutely depressed.
Discussion
The present study investigated an emotional Stroop interference effect at a behavioural and a neural level in medication-free individuals with unipolar depression, during an acute depressive episode of moderate severity. We observed greater response latencies with negative compared to neutral words in MDD patients compared to healthy controls, which was associated with greater activity in the rostral ACC and precuneus.
Increased latencies in patients probably reflect an interference effect in response to self-referent schemas (Beck et al. Reference Beck, Rush, Shaw and Emery1979) and networks (Bower, Reference Bower1981). The negative self-referent schema in depressed patients provides a facilitating basis for incoming negative information resulting in prolonged and sustained processing of mood-congruent information reflected in longer reaction times. Although there was also an overall increase in reaction time across both word types in MDD patients compared to controls, psychomotor slowing and information processing is typical of depression (Caligiuri & Ellwanger, Reference Caligiuri and Ellwanger2000; Tsourtos et al. Reference Tsourtos, Thompson and Stough2002; Rogers et al. Reference Rogers, Kasai, Koji, Fukuda, Iwanami, Nakagome, Fukuda and Kato2004). However, the group×valence interaction effect indicates an even greater emotional Stroop effect in depressed patients relative to healthy controls.
The fMRI data revealed greater engagement in the rostral ACC with negative words relative to neutral words in depressed individuals. The overactive rostral ACC in response to negative words suggests a contribution to the attentional bias towards mood-congruent information. This interpretation is further supported by the finding of a positive correlation between ACC activation and latencies of negative words. This finding suggests that the rostral ACC is associated with the interference process. MDD patients may be less able to disregard the emotional content of the presented words resulting in increased reaction times. The rostral ACC is a critical component in the pathology of depression with a role in assessing emotional salience which is thought to be dysfunctional in depression (Mayberg, Reference Mayberg1997).
Elliott and colleagues (Reference Elliott, Rubinsztein, Sahakian and Dolan2002) similarly found greater ACC activation with sad words in depressed patients and during happy words in controls. In a counting Stroop paradigm, Whalen and colleagues (Reference Whalen, Bush, McNally, Wilhelm, McInerney, Jenike and Rauch1998) found that the emotional condition led to signal increase in the rostral ACC in healthy controls. Moreover, a hyperfunction of the rostral ACC has recently been observed in a fMRI study of an original Stroop task (Wagner et al. Reference Wagner, Sinsel, Sobanski, Kohler, Marinou, Mentzel, Sauer and Schlosser2006). During an incongruent colour- naming condition MDD patients displayed signal increase in the rostral ACC compared to healthy controls (Wagner et al. Reference Wagner, Sinsel, Sobanski, Kohler, Marinou, Mentzel, Sauer and Schlosser2006).
The rostral ACC is involved in attention allocation and conflict monitoring. The emotional Stroop task presents a cognitively demanding challenge to participants, as it requires suppression of a highly trained and therefore very early (in terms of information processing) response, i.e. naming the word, in favour of a more complex process, i.e. naming the colour. Resolving this conflict relies heavily on response selection and executive control. In these circumstances an initial response has to be overridden by a correct response. This overriding process could be reflected in an engagement of the rostral ACC (Botvinick et al. Reference Botvinick, Nystrom, Fissell, Carter and Cohen1999) as mood-congruent trials that withdrew attention from the task instructions led to signal increase in this region. Neutral word conditions did not conflict with the highly overlearned and automated colour- naming procedure in depression.
A recent study in healthy volunteers employing a modified version of an emotional Stroop task also found both rostral ACC and amygdala activation associated with emotional conflict (Etkin et al. Reference Etkin, Egner, Peraza, Kandel and Hirsch2006). Rostral ACC signals increased when conflict resolutions were found while amygdala activation was seen during emotionally conflicting conditions. Etkin and colleagues (Reference Etkin, Egner, Peraza, Kandel and Hirsch2006) conclude that the rostral ACC could be a key regulator of amygdalar activity. Amygdala activation is seen during emotional Stroop tasks in other patient groups in relation to disorder-specific words (van den Heuvel et al. Reference van den Heuvel, Veltman, Groenewegen, Witter, Merkelbach, Cath, van Balkom, van Oppen and van Dyck2005). In the present study, we did not observe greater amygdala activity in depressed patients relative to healthy controls, which may have reflected a top-down suppressive processing from the ACC to the amygdala. This may be examined further in an effective connectivity analysis.
An additional region that was significantly activated in MDD patients during the affective condition was the precuneus. The precuneus holds widespread connections with the prefrontal cortex (Goldman-Rakic, Reference Goldman-Rakic1988) and shows a direct connectivity relationship with the dorsal ACC. Its involvement has been reported in a range of cognitive processes such as response selection (Banich et al. Reference Banich, Milham, Jacobson, Webb, Wszalek, Cohen and Kramer2001), working memory (LaBar et al. Reference LaBar, Gitelman, Parrish and Mesulam1999), problem solving and reasoning (Elliott & Dolan, Reference Elliott and Dolan1998) and verbal processing (Pinel et al. Reference Pinel, Dehaene, Riviere and LeBihan2001; Fu et al. Reference Fu, Vythelingum, Brammer, Williams, Amaro, Andrew, Yaguez, van Haren, Matsumoto and McGuire2006). Engagement of both the rostral ACC and the precuneus probably reflect increased exogenous attention in the ACC to negative words and simultaneously heightened endogenous attention (Posner, Reference Posner1980) to memory retrieval (Nyberg et al. Reference Nyberg, Persson, Habib, Tulving, McIntosh, Cabeza and Houle2000), visual imagery (Krause et al. Reference Krause, Schmidt, Mottaghy, Taylor, Halsband, Herzog, Tellmann and Muller-Gartner1999) and phonological processes (Cabeza & Nyberg, Reference Cabeza and Nyberg2000).
A number of limitations of this study have to be discussed. First, all emotional word stimuli had a negative valence. The observed behavioural and neural effects in patients could have been due to the emotional content rather than the negative valence of the words, although the findings are consistent with neuropsychological theories of Beck (Beck et al. Reference Beck, Rush, Shaw and Emery1979) and Bower (Reference Bower1981). Previous research has also shown that emotional interference in depression is mainly linked to negative word processing. Second, both patients and controls displayed increased latencies in response to negative words (although only very weak in controls), the finding could simply reflect induced conflict rather than a pathological feature of depression. This finding could further imply that negative words had a greater neuronal demand, rather then reflecting greater interference in depressed patients. Future studies should address this caveat by using comparison groups with comparable amounts of cognitive interference as well as words across different emotional categories, to investigate whether neural demands are generally higher during emotional words. This issue touches upon a general problem in fMRI research, namely that of the relationship between brain functional and behavioural measures. In the present study it is difficult to disentangle whether the increased processing of sad stimuli in depressed patients caused enhanced brain activation or vice versa. A final potential limitation of the current study could be the application of an uncorrected significance level. However, as our analysis was hypothesis driven and the findings were confirmed in a ROI analysis this approach is accepted and in accordance with previous MRI literature (Elliott et al. Reference Elliott, Rubinsztein, Sahakian and Dolan2002; Wagner et al. Reference Wagner, Sinsel, Sobanski, Kohler, Marinou, Mentzel, Sauer and Schlosser2006).
To summarize, depression is associated with an emotional interference effect in response to emotionally salient word stimuli on a behavioural and neural level. The behavioural interference effect was reflected in significantly prolonged latencies in response to negative adjectives and verbs in patients with MDD relative to healthy controls. On a neural level, increased engagement of the rostral ACC and precuneus in MDD patients was observed during processing of sad word stimuli. These findings underline the role of the rostral ACC in the determination of the degree of emotional conflict or mood-biased cognitions, particularly in depression. However, further research should be conducted addressing more specifically behavioural and neural interference effects in response to words of different emotional categories.
Acknowledgements
This study was supported by a NARSAD Young Investigator Award to Cynthia Fu. The authors thank the volunteers and the staff of the MRI Unit, Maudsley Hospital, London.
Appendix. Negative and neutral word stimuli

Declaration of Interest
None.



