


Introduction
Antisocial behavior describes a wide variety of actions and attitudes that violate societal norms and the personal or property rights of others (e.g. running away, vandalism, hurting animals, setting fires, theft and bullying/assault). Although generally conceptualized as a unitary construct, extant research has highlighted meaningful distinctions between physically aggressive (AGG; e.g. fighting, hitting, bullying) and non-aggressive rule-breaking (RB; e.g. lying, stealing, vandalism) antisocial behaviors (Loeber & Schmaling, Reference Loeber and Schmaling1985; Frick et al. Reference Frick, Lahey, Loeber, Tannenbaum, Van Horn, Christ, Hart and Hanson1993; Tackett et al. Reference Tackett, Krueger, Sawyer and Graetz2003; Burt, Reference Burt2012). AGG seems to be fairly stable, such that those who are most aggressive in early childhood (∼5–10% of children, mostly boys) continue to be so later in life (Tremblay, Reference Tremblay, Lahey, Moffitt and Caspi2003, Reference Tremblay2010). Even so, overall levels of aggression decrease precipitously from early childhood to adulthood (Tremblay, Reference Tremblay, Lahey, Moffitt and Caspi2003). By contrast, non-aggressive delinquency increases sharply over the course of adolescence, demonstrates less rank-order stability, and although this dimension is also more common in males, the sex difference is less pronounced (Stanger et al. Reference Stanger, Achenbach and Verhulst1997; Moffitt, Reference Moffitt, Lahey, Moffitt and Caspi2003).
These differential developmental trajectories can be most readily understood through the known links between the behavioral dimensions and Moffitt's age-of-onset typology. As discussed explicitly in Moffitt (Reference Moffitt, Lahey, Moffitt and Caspi2003), early-onset youth evidence far higher rates of AGG in particular, whereas adolescence-onset youth are characterized primarily by RB in the absence of AGG (Burt et al. Reference Burt, Donnellan, McGue and Iacono2011; Burt, Reference Burt2012). These links with Moffitt's typology also have provocative implications for the adult outcomes of AGG and RB. High levels of AGG would accordingly be expected to increase risk for academic delay/drop-out, low professional achievement, substance dependency and incarceration/legal problems in adulthood (Moffitt, Reference Moffitt, Lahey, Moffitt and Caspi2003; Burt, Reference Burt2012), speculations that have thus far been borne out (Tremblay, Reference Tremblay, Lahey, Moffitt and Caspi2003; Burt & Hopwood, Reference Burt and Hopwood2010; Burt et al. Reference Burt, Donnellan, McGue and Iacono2011). Adult outcomes for youth with only RB seem to be significantly better. Nevertheless, follow-up in early adulthood (Moffitt et al. Reference Moffitt, Caspi, Harrington and Milne2002) has unexpectedly revealed that these adolescence-onset or RB-only youth continued to commit low-level crimes such as property offenses, evidenced high levels of impulsivity, and reported problems with mental health and substance abuse (Nagin et al. Reference Nagin, Farrington and Moffitt1995; Moffitt et al. Reference Moffitt, Caspi, Harrington and Milne2002; Burt & Hopwood, Reference Burt and Hopwood2010).
AGG and RB also evidence personological distinctions. AGG, for example, is specifically linked to negative emotionality (Moffitt, Reference Moffitt1993, Reference Moffitt, Lahey, Moffitt and Caspi2003; Verona et al. Reference Verona, Patrick and Lang2002; Burt & Larson, Reference Burt and Larson2007; Burt & Donnellan, Reference Burt and Donnellan2008, Reference Burt and Donnellan2009; DeMarte, Reference DeMarte2008; Tackett, Reference Tackett2010). Burt & Donnellan (Reference Burt and Donnellan2008) examined the associations of AGG and RB with personality traits in two independent samples of college students. They reported that, once the overlap across behavioral dimensions was removed, AGG was uniquely associated with high stress reaction (e.g. unaccountable mood changes). Tackett (Reference Tackett2010) later extended these results to the five-factor model of personality in a sample of 907 children aged 6 to 17 years, and again found that high neuroticism was particularly characteristic of AGG. Similarly, the potentiation of negative affect following completion of an aversive task has been associated with AGG but not RB (Burt & Larson, Reference Burt and Larson2007), implying that state levels of negative affect are also largely specific to AGG. Other work indicates that affect dysregulation may be important for precipitating aggressive behavior in particular (Verona et al. Reference Verona, Patrick and Lang2002). Together, these findings suggest that negative emotionality/affective dysfunction represents a core deficit specifically in those with AGG.
By contrast, low constraint/behavioral inhibition is associated more with RB than AGG (Moffitt, Reference Moffitt, Lahey, Moffitt and Caspi2003; Burt & Donnellan, Reference Burt and Donnellan2008, Reference Burt and Donnellan2009; DeMarte, Reference DeMarte2008; Hopwood et al. Reference Hopwood, Burt, Markowitz, Yen, Shea, Sanislow, Grilo, McGlashan, Gunderson, Zanarini, Skodol and Morey2009; Tackett, Reference Tackett2010; Burt et al. Reference Burt, Donnellan and Tackett2012). Burt & Donnellan (Reference Burt and Donnellan2008) found that RB was independently associated with trait levels of impulsivity (i.e. reverse-scored control) once the overlap between the two behavioral dimensions was removed. Similar results were obtained by Tackett (Reference Tackett2010), DeMarte (Reference DeMarte2008), Burt et al. (2012) and Hopwood et al. (Reference Hopwood, Burt, Markowitz, Yen, Shea, Sanislow, Grilo, McGlashan, Gunderson, Zanarini, Skodol and Morey2009), a noteworthy set of replications given that analyses were conducted on independent child and adolescent, college, adjudicated and clinical samples.
AGG and RB also evidence etiological distinctions. A meta-analysis of 103 twin and adoption studies (Burt, Reference Burt2009a) revealed that AGG was a highly heritable condition (genetic influences accounted for 65% of the variance), with little role for the shared environment (i.e. influences that make family members similar to each other regardless of genetic similarity). By contrast, although genetic influences also contributed to RB, these effects were significantly smaller (48%) than those on AGG. Moreover, there was an important role for shared environmental influences on RB (18%). These meta-analytical results have since been extended to other types of ‘natural experiments’ with implications for etiology (Breslau et al. Reference Breslau, Borges, Saito, Tancredi, Benjet, Hinton, Kendler, Kravitz, Vega, Aguilar-Gaxiola and Medina-Mora2011). Breslau et al. (Reference Breslau, Borges, Saito, Tancredi, Benjet, Hinton, Kendler, Kravitz, Vega, Aguilar-Gaxiola and Medina-Mora2011) contrasted AGG and RB across individuals in various stages of migration to the USA: (a) Mexicans living in the USA as adults but who were raised in Mexico, along with Mexicans living in Mexico but with an immediate family member living in the USA, (b) Mexican-Americans born in the USA or Mexicans who came to the USA as children, and (c) those of Mexican ancestry who were born in the USA to at least one US-born parent. Analyses revealed that the effects of migration were markedly stronger for RB compared to AGG. The odds ratios (ORs) for 3 or more symptoms of RB in groups b and c (compared to group a) were 3.45 and 10.50 respectively whereas the ORs for 3 or more symptoms of AGG in groups b and c were 1.51 and 3.07 respectively. These findings imply that RB is more susceptible to environmental/societal influences than is AGG, a conclusion that dovetails nicely with prior meta-analytical conclusions (Burt, Reference Burt2009a).
Although these consistent differences in the relative importance of genetic and environmental influences on AGG compared to RB are suggestive of etiologically driven distinctions between the two dimensions, the results are not as conclusive as one would like in large part because they reveal very little about the presence of common etiological influences on AGG and RB. It thus remains unclear whether the specific genetic and environmental factors contributing to AGG also contribute to RB. Some etiological overlap is to be expected, given that AGG and RB are typically correlated in the 0.4 to 0.6 range and are subsumed within the broader construct of antisocial behavior. However, should the hypothesis of etiological distinctions between AGG and RB be correct, there should be evidence that different genetic and environmental influences also contribute to AGG and RB respectively. Put another way, if AGG and RB are indeed meaningfully distinct from one another, we should find evidence that the specific genetic and environmental effects influencing AGG differ to some extent from those influencing RB.
Available studies have shown some support for etiological differences between AGG and RB. A family study of 273 clinically referred probands and their first-degree biological relatives (n=807) found evidence for specificity in the familial aggregation of AGG and RB (Monuteaux et al. Reference Monuteaux, Fitzmaurice, Blacker, Buka and Biederman2004). Although AGG in the proband predicted AGG in their relatives and RB in the proband predicted RB in their relatives, there was no evidence that proband AGG predicted relatives' RB or vice versa. This specificity in the intergenerational transmission of AGG and RB argues that, in addition to being differentially heritable, AGG and RB may also influenced by distinct etiological factors. Unfortunately, because this was a family study, the specific source of these distinctions (i.e. genetic or environmental) could not be resolved.
Twin/sibling studies overcome this interpretive limitation of family studies, and are therefore well-suited to clarifying the presence of etiological differences between AGG and RB. To date, however, their results have been mixed (Eley et al. Reference Eley, Lichtenstein and Stevenson1999; Bartels et al. Reference Bartels, Hudziak, van den Oord, van Beijsterveldt, Rietveld and Boomsma2003; Gelhorn et al. Reference Gelhorn, Stallings, Young, Corley, Rhee, Hopfer and Hewitt2006; van Hulle et al. Reference van Hulle, Rodgers, D'Onofrio, Waldman and Lahey2007). For example, Gelhorn et al. (Reference Gelhorn, Stallings, Young, Corley, Rhee, Hopfer and Hewitt2006) and van Hulle et al. (Reference van Hulle, Rodgers, D'Onofrio, Waldman and Lahey2007) examined adolescent self-reports of AGG and RB and found evidence of largely independent genetic architectures for AGG and RB (genetic overlap was estimated at 36% and 15% respectively). Shared environmental influences did not overlap at all in either study. By contrast, Eley et al. (Reference Eley, Lichtenstein and Stevenson1999) examined parental informant reports of AGG and RB in two independent samples and reported genetic overlap as high as 76% and shared environmental overlap as high as 100%. Results for the non-shared environment have been more consistent across studies and, moreover, have indicated relatively little overlap. However, given that measurement error is also contained in estimates of the non-shared environment, these differences may or may not be indicative of meaningful distinctions between AGG and RB.
In sum, although the available studies clearly indicate that AGG and RB are differentially heritable, additional work is needed to confirm the presence of meaningful etiological distinctions between them. The current meta-analysis thus sought to clarify the extent to which the etiology of AGG was distinct from that of RB. The moderating roles of sampling and methodological characteristics on effect size estimates were also examined, as these may at least account partially for the aforementioned variation in effect sizes across studies. I specifically examined sex, age and informant effects, in part because they differed across the studies available for analysis, but also because all three have emerged as important moderator variables in prior studies (Eley et al. 1999) and meta-analyses (Burt, Reference Burt2009a,Reference Burtb).
Method
Search strategy
To identify relevant twin/sibling studies, the PsycINFO and Medline databases were examined in summer 2011. The search terms combined each of the following phenotype terms (i.e. delinquency, delinquent, rule-breaking, antisocial, aggression, aggressive) with genetically informative study terms (i.e. twin, adoption, sibling, genetic, environmental) and terms related to their covariance (i.e. genetic correlation, environmental correlation, genetic covariance, environmental covariance). The reference section of each article was then closely examined to identify any studies that may have been missed. Unpublished data examining the covariation between AGG and RB were also collected. Those studies examining genetic and environmental influences on antisocial behavior in general were excluded (n=61), as were those that examined AGG and/or RB but did not examine the overlap between them (n=42). This strategy yielded a total of 13 twin/sibling studies specifically evaluating the covariation between AGG and RB. Inclusion criteria (i.e. construct validity, inability to calculate effect size) are detailed in the following section. Using these criteria, 11 studies examining 12 samples were retained for analysis. All included samples were independent of one another. In cases where the identified study examined longitudinal data (i.e. Burt, in press), the age with the largest n was examined here. The Appendix lists the included samples, along with effect sizes, age, sex, informant and number of sibling pairs.
Inclusion criteria
Construct validity
Included studies were required to meet at least one of the following criteria: (1) the study discriminated between aggressive and non-aggressive symptoms of DSM-III-R or DSM-IV conduct disorder; or (2) there was empirical evidence that the study successfully measured and discriminated between AGG and RB (using factor-analytic techniques, associations with previously validated measure(s) of AGG and RB, etc.). As it was not clear that Barker et al. (2009) met these criteria, the data in question were obtained from the principal investigator of the Twins Early Development Study and analyzed here. Factor analyses did produce evidence of oblique AGG and RB factors, and thus the data were retained for analysis (although unlike in Barker et al. 2009, maternal and teacher informant reports, in addition to opposite-sex and female–female twin pairs were also included).
Inability to compute study effect sizes
The study effect sizes used in this meta-analysis were genetic and environmental correlations, which index the proportion of genetic and environmental influences, respectively, that overlap across AGG and RB (described later in more detail). Proportions of co-occurrence explained by genetic and environmental influences (e.g. the proportion of their phenotypic correlation that was genetic in origin) were not examined here, and this information reveals relatively little about the proportion of genetic influences shared between two traits. For example, should two phenotypes share 100% of their genetic influences but also share some environmental influences, the proportion of covariance accounted for by genetic effects would be less than 100%.
One study was excluded because effect sizes were not reported and could not be calculated (Tackett et al. Reference Tackett, Krueger, Iacono and McGue2005). Tackett et al. (Reference Tackett, Krueger, Iacono and McGue2005) evaluated the origins of self-reported aggressive and rule-breaking DSM-III-R symptom counts across two independent cohorts of adolescent male twins participating in the Minnesota Twin Family Study. The covariance between AGG and RB was reported to be 51% genetic, 28% shared environmental and 21% non-shared environmental in origin. However, genetic and environmental correlations, the effect sizes of interest here, were not presented. Fortunately, relevant data from one of the two cohorts were examined in another study (Burt, in press), and were thus already included here. Data from the second cohort were obtained from the principal investigators of the Minnesota Twin Family Study (see Appendix). In both cases, female twins were also examined.
Analyses
Behavioral genetic analyses make use of the difference in the proportion of segregating genes shared between reared-together siblings to make inferences about etiology. Monozygotic (MZ) twins result from a single fertilized zygote splitting into two and hence share 100% of their segregating genes. Dizygotic (DZ) twins are the result of two independent conceptions and so, like all full siblings, share an average of 50% of their segregating genes. Half-siblings share only one of their two parents, and thus share an average of 25% of their segregating genetic material. Adoptive siblings do not share any segregating genetic material. Using these differences, the variance within and covariance between phenotypes can be partitioned into three components: additive genetic (a 2), shared environment (c 2) and non-shared environment plus measurement error (e 2). The additive genetic component is the effect of individual genes summed over loci. The shared environment is that part of the environment that is common to both members of a sibling pair, and acts to make the siblings within a pair similar to each other regardless of the proportion of genes shared. The non-shared environment differentiates each sibling within a pair, making them less similar. Measurement error is also contained within e 2. More information on twin and sibling studies is available elsewhere (Plomin et al. Reference Plomin, DeFries, McClearn and McGruffin2008).
The current analyses center on the genetic, shared and non-shared environmental correlations obtained from bivariate or multivariate correlated factors or decomposition models (the former is illustrated in Fig. 1). These models parse the phenotypic variances of both AGG and RB, in addition to the phenotypic covariance between them, into that which is due to genetic, shared environmental and non-shared environmental factors. These covariances can then be standardized on their respective variances to produce the genetic, shared environmental and non-shared environmental correlations pictured in Fig. 1. These statistics reveal the extent to which a specific effect on one variable is correlated with the same effect on another variable. When squared, these correlations directly index the proportion of genetic (or environmental) influences shared by two variables. A genetic correlation of 0, for example, would indicate that none of genetic influences on AGG overlap with those on RB. A genetic correlation of 1.0, by contrast, would indicate that 100% of the genetic influences on AGG overlap with those on RB. Note that, because genetic and shared environmental influences do not contain measurement error, it is entirely possible (and indeed, not uncommon; see Appendix) to see genetic and shared environmental correlations of 1.0.

Fig. 1. Bivariate model. This model estimates the genetic (A), shared (C) and non-shared environmental (E) contributions to the variance within, and the covariance between, aggressive (AGG) and non-aggressive rule-breaking (RB) antisocial behavior. Path coefficients (e.g. a 1) are squared to index the percentage of individual variance accounted for. Genetic, shared and non-shared environmental correlations (r A, r C and r E respectively) are indicated by the double-headed arrows in the upper portion of the diagram, and can be squared to index the proportion of etiological overlap.
Specific analyses
A mixed effects model, estimated using iterative maximum likelihood, was used to perform the analyses. In accordance with extant recommendations (Lipsey & Wilson, Reference Lipsey and Wilson2001; Wilson, Reference Wilson2005), individual correlations were converted to z statistics using Fisher's r to z transformation prior to analysis. The resulting effect sizes were then weighted using a combination of inverse variance weights and the random effects variance component. Following analyses, estimates were converted back into correlations using an inverse z to r transformation. Tests of homogeneity (i.e. the Q statistic, which is distributed as a χ2) were also conducted (Lipsey & Wilson, Reference Lipsey and Wilson2001; Wilson, Reference Wilson2005) to evaluate statistically the possibility that effect sizes vary across studies.
Possible moderators of the effect sizes (i.e. age, informant and sex) were then evaluated. Close inspection of the method sections revealed that at least 75% of the participants in a given study could be characterized as either children (ages 6–10 years; n=5690 pairs) or adolescents (ages 11–18 years; n=7233 pairs). This distinction was thus retained for analysis. Informant analyses compared estimates obtained from maternal, teacher and child informant reports (mother n=8406 pairs, teacher n=3843 pairs, child n=5224 pairs). Maternal reports were defined here as those reports specifically from mothers and those under the more ambiguous term of ‘parent’, as careful reading of method sections revealed that informants for parent reports were almost always mothers. Analyzing sex was less straightforward, as only three samples reported effect sizes separately for boys and girls. The remaining samples presented results without regard to sex (because they did not find evidence of sex differences in etiology). To avoid biasing the results by omitting these data, constrained effect sizes from these samples were included in the analyses of boys (n=6238 pairs) and girls (n=6685 pairs).
Although there are no formal guidelines for semantically describing the proportion of variance explained within the field of behavioral genetics, a meta-analysis of univariate results (Burt, Reference Burt2009b) identified the following descriptive pattern from 30 articles: effects <20% (i.e. 1–19%) were described as ‘modest’ or ‘small’, effects in the range 20–49% were described as ‘moderate’, and effects ⩾50% were described as ‘large’. The use of these descriptive guidelines in the current study is bolstered by the very high levels of genetic stability seen for AGG and RB over time (van Beijsterveldt et al. Reference van Beijsterveldt, Bartels, Hudziak and Boomsma2003; Tuvblad et al. Reference Tuvblad, Narusyte, Grann, Sarnecki and Lichtenstein2011; Burt, in press). van Beijsterveldt et al. (Reference van Beijsterveldt, Bartels, Hudziak and Boomsma2003), for example, examined the stability of AGG in The Netherlands Twin Register, and found that genetic influences on parental reported AGG were highly stable from ages 10 to 12 years (the age 12 data were examined in Bartels et al. Reference Bartels, Hudziak, van den Oord, van Beijsterveldt, Rietveld and Boomsma2003; see Appendix). The respective genetic correlations for boys and girls were 0.91 and 0.89, indicating that 79–83% of genetic influences on AGG at age 10 were still important at age 12. Burt (in press; see Appendix) similarly examined the stability of self-reported AGG and RB from ages 14 to 17 years, and found genetic correlations of 0.85 for RB and 0.78 for AGG, indicating high levels of genetic overlap (61–72%) across ages 14 and 17. Finally, Tuvblad et al. (Reference Tuvblad, Narusyte, Grann, Sarnecki and Lichtenstein2011) found that 67% of genetic influences on antisocial behavior in general persisted from ages 8 to 20 years (the age 8 data were examined in Eley et al. Reference Eley, Lichtenstein and Stevenson1999; see Appendix). The current study thus made use of the extant informal guidelines, describing effects <20% as ‘modest’, effects between 20% and 49% as ‘moderate’, and effects ⩾50% as ‘large’.
Results
Average genetic and environmental correlations were computed using data from all included studies (see Table 1). The mixed effects estimate of the genetic correlation was 0.62, results that are indicative of only moderate genetic overlap (38.4%) between AGG and RB. In other words, the current results indicate that AGG and RB share some, but certainly not all, of their genetic influences. A high degree of overlap, although still less than unity, was observed for shared environmental influences (64.0%). Non-shared environmental influences overlapped only minimally (10.2%) indicating that the child-specific environmental experiences (and measurement error) that contribute to AGG and RB are largely distinct.
Table 1. Overall meta-analytic estimates of genetic (rA), shared environmental (rC) and non-shared environmental (rE) correlations between aggressive (AGG) and non-aggressive rule-breaking (RB) antisocial behavior, obtained by mixed effects models (n=12 923 sibling pairs)

CI, Confidence interval.
Proportion of etiological overlap indexes the respective proportions of genetic, shared and non-shared environmental variance on AGG that overlaps with that on RB. As such, this row does not sum to 100%. All correlations were significantly greater than zero at p<0.001.
There was evidence of considerable heterogeneity in effects across studies, and particularly so for shared environmental effect sizes [i.e. Q (on 23 df)=582.9, 16,047.3 and 112.6 for genetic, shared environmental and non-shared environmental effect sizes respectively; all p<0.001]. Given these results, several potential moderators of study effect sizes (i.e. age, informant, sex) were examined via two types of mixed effects models: (1) inverse variance weighted one-way ANOVAs (sometimes called Analogs to the ANOVA), which evaluate mean differences in a given effect size across levels of a given moderator; and (2) weighted multiple regression, which analyzes the simultaneous influence of multiple moderators on a given effect size.
Genetic correlations (rA)
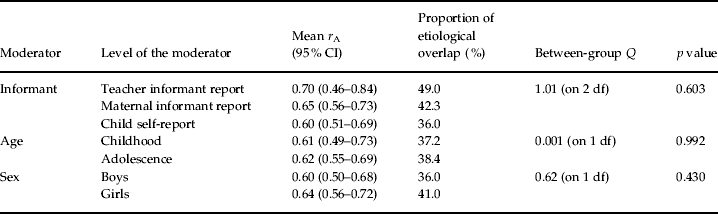
Mean genetic correlations obtained through inverse variance weighted one-way ANOVAs are presented, separately by informant, age and sex, in Table 2. As shown, there was no evidence that informant, age or sex individually moderated the level of genetic overlap between AGG and RB. When all three moderators were entered simultaneously into a mixed effects weighted multiple regression, however, there was evidence of at least some moderation by informant and age [β values were as follows: informant=–0.22 (p=0.033); age=0.28 (p=0.050); sex=0.066 (p=0.49)]. In particular, r A was lower when using child reports and during childhood, although the former seemed to obscure the latter when examining age by itself (probably because child self-reports were available only for adolescents).
Table 2. Genetic correlations (rA) between aggressive (AGG) and non-aggressive rule-breaking (RB) antisocial behavior across a series of moderators
CI, Confidence interval; df, degrees of freedom.
Proportion of etiological overlap indexes the proportion of genetic overlap across AGG and RB. As such, this column does not sum to 100%. All correlations with CIs that do not overlap with zero are significantly greater than zero.
r A did not vary significantly across levels of any moderator (as evidenced by statistically non-significant between-group Q values).
Shared environmental correlations (rC)
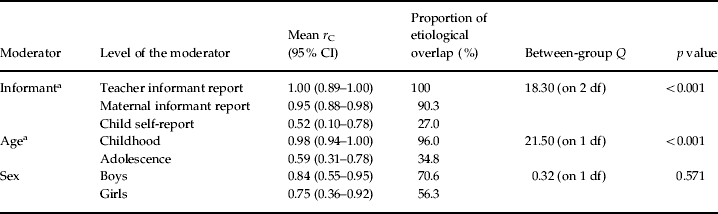
Mean shared environmental correlations are presented separately by informant, age and sex in Table 3. As shown, there were pronounced differences in shared environmental correlations across both informant and age. Shared environmental overlap was at or near unity for teacher and parent informant reports (100% and 90.3% respectively) and was considerably lower for child self-reports (27.0%; pairwise comparisons: teacher versus child, p=0.002; mother versus child, p=0.0004). Shared environmental overlap also decreased significantly with age (96.0% in childhood versus 34.8% in adolescence). Moreover, these effects generally persisted even when all three moderators were entered simultaneously into a multiple regression [β values were as follows: informant=–0.67 (p=0.059); age=–0.98 (p=0.04); sex=–0.21 (p=0.51)].
Table 3. Shared environmental correlations (rC) between aggressive (AGG) and non-aggressive rule-breaking (RB) antisocial behavior across a series of moderators
CI, Confidence interval; df, degrees of freedom.
Proportion of etiological overlap indexes the proportion of shared environmental overlap across AGG and RB. As such, this column does not sum to 100%. All correlations with confidence intervals that do not overlap with zero are significantly greater than zero.
a r C varied significantly across levels of that moderator (as evidenced by a statistically significant between-groups Q).
Non-shared environmental correlations (rE)
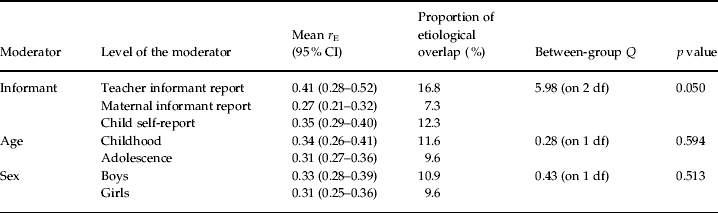
Mean non-shared environmental correlations are presented separately by informant, age and sex in Table 4. As shown, r E did not vary across either age or sex (the range of overlap was 9.6% to 11.6%). There was some evidence of differential overlap across informant reports, however, such that non-shared environmental correlations were slightly smaller, if modestly so, for samples assessed using maternal reports (7.3%) compared to teacher reports or child self-reports (16.8% and 12.3% respectively; pairwise comparisons: teacher versus mother, p=0.042; child versus mother, p=0.067). When analyzed alongside the other moderators, informant effects continued to emerge as statistically significant [β values were as follows: informant=−0.09 (p=0.01); age=–0.03 (p=0.45); sex=–0.02 (p=0.57)].
Table 4. Non-shared environmental correlations (rE) between aggressive (AGG) and non-aggressive rule-breaking (RB) antisocial behavior across a series of moderators
CI, Confidence interval; df, degrees of freedom.
Proportion of etiological overlap indexes the proportion of non-shared environmental overlap across AGG and RB. As such, columns do not sum to 100%. All correlations with confidence intervals that do not overlap with zero are significantly greater than zero.
r E did not vary significantly across levels of any moderator (as evidenced by statistically non-significant between-group Q values).
Discussion
The primary aim of the current meta-analysis was to clarify the extent to which genetic and environmental influences on AGG overlapped with those on RB. Thirteen twin/sibling studies examining the covariation between AGG and RB were collected, of which 11 (examining 12 samples) were included in the analyses. The results strongly supported prior suggestions that AGG and RB constitute meaningfully distinct dimensions within the broader construct of antisocial behavior. Mixed effect models revealed that genetic influences on AGG were largely, although not entirely, distinct from those on RB: only 38.4% of the genetic influences on AGG overlapped with those on RB. Similarly, only 10.2% of non-shared environmental influences on AGG overlapped with those on RB. Shared environmental influences overlapped considerably more, albeit inconsistently so.
Importantly, the conclusion that genetic and non-shared environmental influences on AGG are partially distinct from those on RB was found to persist across several potential moderators. Despite this consistency, however, the moderator analyses also indicated that the extent of etiological overlap often varied across childhood versus adolescence and/or across the specific informant report used. Shared environmental overlap was observed to be significantly stronger during childhood than during adolescence: indeed, it was essentially unity during childhood and was more moderate during adolescence (96.0% and 34.8% respectively). These findings suggest that, during childhood, the family-wide environmental experiences contributing to the presence of AGG are nearly identical to those contributing to RB. By adolescence, however, the shared environmental factors influencing AGG are more distinct from those influencing RB. Such findings are likely to at least partially reflect the fact that shared environmental influences on AGG are small, both in absolute terms and when compared to those on RB. This is particularly the case during adolescence, when shared environmental influences on AGG are estimated to be near-zero (Burt, Reference Burt2009a). In this light, the proportion of shared environmental overlap may not be all that meaningful. To the extent that it is meaningful, however, it is noteworthy that parental divorce and parent–child conflict have consistently emerged as sources of shared environmental variance in antisocial behavior during adolescence (Burt et al. Reference Burt, McGue, Krueger and Iacono2007, Reference Burt, Barnes, McGue and Iacono2008; Klahr et al. Reference Klahr, McGue, Iacono and Burt2011). The current results could thus imply that these risk factors may be differentially related to AGG and RB. Future work should explore this possibility.
Etiological influences on AGG also overlapped more with those on RB depending on the informant reports used in the analyses. The use of child self-reports seemed to minimize genetic and shared environmental overlap whereas non-shared environmental overlap was smallest when using maternal informant reports. Although somewhat troubling, these results are not particularly surprising, as it is well known that heritability estimates vary by informant (e.g. genetic influences on child and adolescent conduct problems in Burt, Reference Burt2009b were estimated to be 59% for maternal informant reports, 68% for teacher informant reports, and 37% for child self-reports). Although the reasons behind this variation remain unclear, it seems likely that they at least partially reflect the shared method variance inherent in maternal informant reports and most teacher informant reports (i.e. one adult reports on both siblings and those reports are correlated), a methodological limitation that is entirely circumvented by the use of child self-reports (i.e. each sibling reports on themselves and these reports are correlated). To the extent that shared method variance is an issue in the maternal and teacher data examined here, it would have key consequences for the conclusions of the current meta-analysis, in that there was relatively less genetic and shared environmental overlap between AGG and RB when using child self-report. It may thus be the case that AGG and RB are even more etiologically distinct than would be suggested by our overall results.
Shared method variance is not the only possible explanation, however. Informant effects may also reflect the different patterns of phenotypic correlation observed across various informant reports. In particular, across several nationally and internationally representative samples of children and adolescents (as reviewed in Burt, Reference Burt2012), phenotypic correlations were lower when examining youth self-reports of AGG and RB (average r=0.43) as compared to parental informant reports (average r=0.65). Although such results may mean that parents have trouble distinguishing between AGG and RB when they report on their children's behavior, another interpretation is that children are successfully concealing at least some of their antisocial behaviors from their parents, making it more difficult for parents to accurately report on their children's behavior. The latter idea is bolstered by the finding that adolescents report roughly twice as many symptoms of conduct disorder as do their parents (Hewitt et al. Reference Hewitt, Silberg, Rutter, Simonoff, Meyer, Maes, Pickles, Neale, Loeber, Erickson, Kendler, Heath, Truett, Reynolds and Eaves1997). Alternately, it may mean that child self-reports are more prone to measurement error, a bias that would serve to suppress phenotypic correlations between AGG and RB, and perhaps genetic and shared environmental contributions to these correlations as well. Future work should attempt to evaluate these different possibilities.
There are several limitations to the current meta-analysis. First, although the current study is useful for clarifying sources of etiological overlap between AGG and RB during childhood and adolescence, it remains unclear how these results may generalize to adulthood. Adult samples were excluded from these analyses, primarily because available twin studies on this topic have centered on younger samples. Future work should explore sources of etiological overlap between AGG and RB during adulthood, particularly as other work has indicated that the behavioral dimensions of AGG and RB persist beyond adolescence (Burt & Donnellan, Reference Burt and Donnellan2009; Kendler et al. Reference Kendler, Aggen and Patrick2012). Second, it is unclear to what extent positive skew in the distributions of AGG and RB might have influenced our results, particularly in light of the higher estimates of non-shared environment obtained in severely skewed data (see Burt, Reference Burt2009b). Although future work should clarify the extent to which non-normal distributions might influence genetic and environmental correlation estimates, it is worth noting that the genetic correlations obtained in Burt (in press; r A=0.53 and 0.64) were on a par with the meta-analytic estimates even though the data in those samples were not skewed. Third, the current study did not directly examine how age of onset impacted the results. As noted, however, the aggressive/non-aggressive distinction examined here seems to map closely on to Moffitt's age-of-onset taxonomy (Moffitt, Reference Moffitt1993, Reference Moffitt, Lahey, Moffitt and Caspi2003). The present finding of prominent etiological distinctions between AGG and RB thus offers indirect support for Moffitt's hypothesis of etiological distinctions between life-course persistent and adolescence-limited antisocial behavior.
Fourth, only a few available studies (13 out of 116) were appropriate for the current meta-analysis. This is potentially problematic, as it may mean that the current results are skewed in some unknown way. Fortunately, most of the excluded studies (∼70%) were redundant with those examined here, primarily because of multiple publications using the same data. In other cases, however, the studies were simply excluded. Some twin samples (e.g. VTSABD) have examined the etiologies of AGG and RB but have not examined the sources of their covariation, and thus could not be included here. Finn Twin, the Quebec Newborn Twin Register and the Wisconsin Twin Panel, by contrast, have not examined RB (to my knowledge), and were thus omitted here. Their exclusive focus on AGG may reflect the ages of their sample (e.g. some examine toddlers), an absence of factor analytic support for separable AGG and RB dimensions in a given sample and/or a primary focus on other aspects of human behavior. Nevertheless, at least some of the excluded studies may well have data that would allow for additional examinations of AGG and RB [e.g. DSM symptom counts, the Child Behavior Checklist (CBCL), etc.]. Future studies should seek to further clarify the origins of the covariation between AGG and RB.
Conclusions
The results of the current meta-analysis strongly support prior suggestions that AGG and RB constitute meaningfully distinct dimensions within the broader construct of antisocial behavior. Only 38.4% of the genetic influences on AGG overlapped with those on RB. Although shared environmental influences evidenced high (if inconsistently so) levels of overlap, only 10.2% of non-shared environmental influences on AGG overlapped with those on RB. In short, AGG and RB seem to be largely, although not entirely, phenotypes at the etiological level. Such findings serve to not only underscore the presence of important etiological differences between AGG and RB but also highlight the need for future research to distinguish between the behavioral dimensions when studying the causal processes that underlie antisocial behavior. For example, the well-known association between antisocial behavior and executive dysfunction has been shown to vary across AGG and RB, such that executive functioning is negatively related to AGG but is either unrelated or positively related to RB (Barker et al. Reference Barker, Seguin, White, Bates, Lacourse, Carbonneau and Tremblay2007, Reference Barker, van Lier, Vitaro, Nagin, Assaad, Tremblay and Ségiun2011). When combined with the personological distinctions between AGG and RB noted in the introduction, the above findings begin to resolve the different nomological nets surrounding AGG and RB. Future conceptualizations of antisocial behavior should be organized (at least in part) around the dimensions of AGG and RB.
Declaration of Interest
None.
Appendix Effect sizes by study







