


Introduction
People with schizophrenia have performance deficits on tests of executive function at all stages of the illness (Pantelis et al. Reference Pantelis, Barnes, Nelson, Tanner, Weatherley, Owen and Robbins1997; Hutton et al. Reference Hutton, Puri, Duncan, Robbins, Barnes and Joyce1998). An important example of this is impaired cognitive flexibility reflected in increased errors on the Wisconsin Card Sort Test (WCST) (Grant & Berg, Reference Grant and Berg1948; Nelson, Reference Nelson1976; Weinberger et al. Reference Weinberger, Berman and Zec1986; Goldberg & Weinberger, Reference Goldberg and Weinberger1994; Berman et al. Reference Berman, Ostrem, Randolph, Gold, Goldberg, Coppola, Carson, Herscovitch and Weinberger1995; Prentice et al. Reference Prentice, Gold and Buchanan2008) and the CANTAB ID/ED task (Pantelis et al. Reference Pantelis, Barnes, Nelson, Tanner, Weatherley, Owen and Robbins1997; Hutton et al. Reference Hutton and Ettinger1998; Waltz & Gold, Reference Waltz and Gold2007; Murray et al. Reference Murray, Cheng, Clark, Barnett, Blackwell, Fletcher, Robbins, Bullmore and Jones2008; Leeson et al. Reference Leeson, Robbins, Matheson, Hutton, Ron, Barnes and Joyce2009). Successful performance on these tasks depends on a number of control operations, such as maintenance of task goals in working memory, inhibition of pre-potent responses, monitoring of own behaviour and associated feedback. Being able to identify more discrete deficits of this type will allow a greater understanding of the processes contributing to cognitive dysfunction in schizophrenia, their neurobiological basis and how they might impact on clinical outcomes (Carter et al. Reference Carter, Barch, Buchanan, Bullmore, Krystal, Cohen, Geyer, Green, Nuechterlein, Robbins, Silverstein, Smith, Strauss, Wykes and Heinssen2008).
One line of schizophrenia research that has proved fruitful in this regard has focused on more simple paradigms, in which the task demands are clearly defined and, by using eye movements, the responses are accurately measured. For example, schizophrenia patients are consistently found to make increased errors in the anti-saccade task (Fukushima et al. Reference Fukushima, Fukushima, Chiba, Tanaka, Yamashita and Kato1988; Hutton & Ettinger, Reference Hutton and Ettinger2006), possibly due to weak internal representations of task goals in working memory (Reuter et al. Reference Reuter, Jager, Bottlender and Kathmann2007). This has been linked to frontal cortex impairment since patients with lesions in the dorsal and ventrolateral frontal cortex have similar deficits (Guitton et al. Reference Guitton, Buchtel and Douglas1985; Walker et al. Reference Walker, Husain, Hodgson, Harrison and Kennard1998; Hodgson et al. Reference Hodgson, Chamberlain, Parris, James, Gutowski, Husain and Kennard2007).
A number of studies have also investigated how patients with schizophrenia switch between two saccadic tasks, which is considered to reflect cognitive flexibility in the sense of being able to adapt behaviour quickly in response to changing environmental contingencies. Manoach et al. (Reference Manoach, Lindgren, Cherkasova, Goff, Halpern, Intriligator and Barton2002) required subjects to switch between anti-saccades and ‘pro-saccades’ and surprisingly found that schizophrenia patients were normal, showing similar ‘switch costs’ to healthy controls (see also Franke et al. Reference Franke, Reuter, Schulz and Kathmann2007; Greenzang et al. Reference Greenzang, Manoach, Goff and Barton2007). However, this contrasts with work using non-oculomotor switching tasks, in which schizophrenia patients are impaired (Meiran et al. Reference Meiran, Levine, Meiran and Henik2000). Given that a number of healthy volunteer studies report no switch cost or even a small benefit when switching between pro- and anti-saccades (Hallett & Adams, Reference Hallett and Adams1980; Hunt & Klein, Reference Hunt and Klein2002; Hodgson et al. Reference Hodgson, Golding, Molyva, Rosenthal and Kennard2004; Parton et al. Reference Parton, Nachev, Hodgson, Mort, Thomas, Ordidge, Morgan, Jackson, Rees and Husain2007), it is possible that this finding is an artefact of the testing procedure. Hodgson et al. (Reference Hodgson, Golding, Molyva, Rosenthal and Kennard2004) suggest that switching to and from a reflexive response – the pro-saccade – does not require retrieval and reconfiguration of arbitrary stimulus–response mappings and it is arguably these specific operations that constitute the main challenge of task switching in non-oculomotor paradigms, for example, switching versions of the Stroop task (see Monsell, Reference Monsell2003).
In order to further investigate the processes involved in task switching and how they are affected in schizophrenia, we used a novel oculomotor ‘rule switching’ task, in which participants learn a rule linking a central symbolic cue with a saccade to either the left or the right (Hodgson et al. Reference Hodgson, Golding, Molyva, Rosenthal and Kennard2004). The rule can reverse at different points in the task, as indicated by a change in the feedback presented following the response. This is a closer analogue of non-oculomotor task switching paradigms as it involves coordination of arbitrary stimulus response mappings, requiring suppression of recently correct responses on a subset of trials. Having to suppress particular responses on some trials but not others is of interest because we have found evidence of such an inhibitory impairment in schizophrenia using the stop-signal task (Huddy et al. Reference Huddy, Aron, Harrison, Barnes, Robbins and Joyce2009). We therefore predicted that patients would show increased errors on trials following rule switches.
Two other processes essential for appropriate switching behaviour can be measured with this oculomotor task. One is the ability to monitor responses as reflected in the rate of error correction when a saccade is initially made in the wrong direction (see Husain et al. Reference Husain, Parton, Hodgson, Mort and Rees2003). Although schizophrenia patients have been shown to correct the majority of errors in the anti-saccade task (Polli et al. Reference Polli, Barton, Vangel, Goff, Iguchi and Manoach2006), the rule switching task may be more taxing because the demand to inhibit a response varies from trial to trial.
Another process is the requirement to use positive and negative feedback to guide responses. Studies of the WCST and CANTAB ID/ED found that schizophrenia patients do not use negative feedback effectively (Murray et al. Reference Murray, Cheng, Clark, Barnett, Blackwell, Fletcher, Robbins, Bullmore and Jones2008; Prentice et al. Reference Prentice, Gold and Buchanan2008; Gold et al. Reference Gold, Waltz, Prentice, Morris and Heerey2008; Leeson et al. Reference Leeson, Robbins, Matheson, Hutton, Ron, Barnes and Joyce2009). Using the rule switching task in healthy volunteers, Hodgson et al. (Reference Hodgson, Mort, Chamberlain, Hutton, O'Neill and Kennard2002) found that responses are slower to locations that have just been the site of negative feedback [termed ‘reward-related inhibition of return’ in contradistinction to the ‘inhibition of return’ effect seen in studies of covert attention (Posner et al. Reference Posner, Rafal, Choate and Vaughan1985)]. This task therefore allows a further understanding of negative feedback processing in schizophrenia.
Using this oculomotor task, we investigated rule switching in schizophrenia and how this relates to response inhibition, error monitoring and feedback processing. Findings on this task in patients with focal frontal cortex lesions (Hodgson et al. Reference Hodgson, Chamberlain, Parris, James, Gutowski, Husain and Kennard2007) also allowed us to make inferences about the possible neurobiological substrates of impaired performance.
Method
Participants
Altogether, 50 medicated patients with first-episode psychosis were recruited from the West London longitudinal study (for details, see Huddy et al. Reference Huddy, Hodgson, Kapasi, Mutsatsa, Harrison, Barnes and Joyce2007). Initial diagnoses were ascertained using The Diagnostic Interview for Psychosis (Jablensky et al. Reference Jablensky, Sartorius, Ernberg, Anker, Korten, Cooper, Day and Bertelsen1992) and were reviewed 1 year later. The final DSM-IV diagnoses were schizophrenia (47) and schizoaffective disorder (3). These were compared with 25 healthy volunteers recruited from the same catchment area. Permission to conduct the study was obtained from Merton, Sutton and Wandsworth, Riverside, and Ealing Research Ethics Committees. All participants gave written informed consent and were paid an honorarium for their time.
Procedure
Clinical and neuropsychological assessments
Symptoms were assessed at recruitment using Scales for the Assessment of Positive Symptoms (Andreasen, Reference Andreasen1984) and Negative Symptoms (Andreasen, Reference Andreasen1983) and positive, disorganization and negative syndrome scores were derived (Huddy et al. Reference Huddy, Hodgson, Kapasi, Mutsatsa, Harrison, Barnes and Joyce2007). Cognitive assessments were performed a median of 8 days later as follows: pre-morbid IQ with the Wechsler Test of Adult Reading (Wechsler, Reference Wechsler2001); current IQ with four Wechsler Adult Intelligence Scale (WAIS) III subtests (Wechsler, Reference Wechsler1999) validated for schizophrenia (Blyler et al. Reference Blyler, Gold, Iannone and Buchanan2000); working memory with CANTAB tests of Spatial Span (Owen et al. Reference Owen, Downes, Sahakian, Polkey and Robbins1990), which measures the ability to remember the order of sequences of squares presented on the screen in increasing number; Spatial Working Memory (Owen et al. Reference Owen, Downes, Sahakian, Polkey and Robbins1990), where patients are required to ‘open’ sets of boxes to find tokens and errors are recorded when boxes in which tokens have been found are reopened.
Eye movement recording and analysis
Eye movements were recorded using the Eyelink system (SR Research, Canada), a video-based pupil tracker, with head movement compensation system sampling at 250 Hz. Subjects sat in front of the display monitor approximately 60 cm from the screen. Pupil position was monitored via two miniature infrared CCD video cameras mounted on an adjustable headband. Participants were instructed to keep head movements to a minimum and no active restraint of head movements was required to obtain sufficiently accurate gaze position recordings. Eye movements were visualized offline, saccades were identified and artefacts removed using custom software programs developed within the LabVIEW (National Instruments Corporation, USA) visual programming environment.
Saccadic rule switching task
Three boxes, outlined in black on a dark grey-coloured background, were presented in the centre and 9° to the left and right of the screen. Each box subtended 3° of visual angle. Trial onset was triggered when the subject had been fixating the central box for 800 ms. At this point, a blue or yellow circle was presented in the central box. The subject was instructed to look either to the left or right box whenever a coloured circle appeared. The colour of the cue (yellow/blue) instructed the subject whether to look left or right. The next fixation >800 ms on either the left or the right box was taken as the subject's response on that trial, such that an eye fixation of shorter duration could be made towards the alternate location before the subject made their final decision. Once the viewer had selected one of the boxes by fixating it for >800 ms, feedback was given to indicate if the choice was correct or incorrect in the form of a happy/sad face displayed within the selected box, accompanied by a high- or low-pitched tone. Subjects were made aware that the rule linking the colour of the cue and direction of saccade would reverse at several points during the test. Rule changes were indicated by unexpected errors following runs of between nine and 13 correct response trials. Each subject completed one block of 100 trials, comprising a maximum of eight possible rule reversals. They were instructed to perform the task as quickly and as accurately as possible and to respond on the basis of the rule they know to be correct at that time, without anticipating the occurrence of a rule change.
Results
Patients and controls were matched for age (see Table 1). Patients scored significantly lower on most neuropsychological tests, with a tendency to perform worse on the Spatial Working Memory task.
Table 1. Demographics and neuropsychological performance of the two patient groups and controls

WTAR, Wechsler Test of Adult Reading; WAIS, Wechsler Adult Intelligence Scale; SWM, spatial working memory.
* p<0.05 ** p<0.01, † p<0.1.
Overall latencies and errors
Independent sample t tests revealed significant differences in overall latency [t(73)=2.1, p<0.05], overall errors [t(73)=2.8, p<0.01] and rules completed [t(73)=2.8, p<0.01], patients being slower and more error-prone than controls. Five patients failed to achieve the learning criterion of six consecutive correct responses more than once and were excluded from subsequent analyses.
Interaction between errors and rule switching
Errors that were subsequently corrected with a secondary saccade prior to the feedback, i.e. corrected errors (Fig. 1), were initially separated from errors that were not corrected within the deadline, i.e. uncorrected errors. A mixed three-factor analysis of variance (ANOVA) was applied using group (controls versus patients), error type (corrected versus uncorrected) and trial (first to sixth following a rule change).

Fig. 1. (a) Rule reversal task. Subjects learn a rule linking a coloured shape with a movement to either the left or the right. After a random number of trials the rule can reverse. The task is self-paced with at least 1500 ms elapsing between each trial; (b) Corrective saccades. Feedback is only given following a fixation >800 ms on one of the response boxes. On a proportion of trials, participants make saccade errors followed by a corrective movement towards the correct response box. Both examples are corrected errors.
There was a main effect of trial [F(5, 340)=18.7, p<0.001] and a trend towards a main effect of group [F(1, 68)=3.1, p<0.1] (see Fig. 2). This finding was qualified by significant trial×group [F(5, 340)=3.5, p<0.01] and error type×group [F(5, 340)=5.4, p<0.05] interactions. There was no three way interaction of trial×error type×group [F(5, 340) <1].

Fig. 2. Latencies and error rates for patients and control groups in the rule switching task, showing proportion of total errors which were corrected, plotted against trial after rule change.
The error type×group interaction was due to patients making proportionally more uncorrected errors than controls overall. Inspection of the data suggested that the patients made more errors of both types on trial 1 only. To investigate this group×trial effect, the ANOVA was repeated for the second to sixth trials inclusive, excluding trial 1, and this revealed that the group×trial interaction was no longer significant [F(4, 272)<1]; there was also no group main effect [F(1, 68)<1]. Thus, the significant group×trial interaction found in the first analysis was driven by higher errors in patients on trial 1. This was confirmed by an ANOVA, using group and errors as factors, carried out for trial 1, which showed that patients made more errors than controls [F(1, 68)=10.7, p<0.01] of both types.
Interaction between response latencies and rule switching
A two-factor ANOVA was applied to mean response latencies for the first saccade following cue onset on correct response trials, i.e. when the first saccade was to the correct location; the factors were group and trial (first to sixth following a rule change). There was a main effect of trial [F(5, 335)=7.9, p<0.001] but no group difference or trial×group interaction [F(5, 335) <1]. Fig. 2 indicates that both groups showed a slowing of response times immediately following a rule change. This interpretation was confirmed by an analysis limited to trials 2–6, inclusive, which demonstrated no main effect of trial [F(4, 268)<1].
In summary, patients made more errors on the first trial following a rule change than the control group. On trials performed entirely correctly, patients showed equivalent slowing of response latencies on trial 1 following a rule change.
Reward-related inhibition of return effect on latencies
Hodgson et al. (Reference Hodgson, Mort, Chamberlain, Hutton, O'Neill and Kennard2002, Reference Hodgson, Golding, Molyva, Rosenthal and Kennard2004) demonstrated a location specific ‘inhibition of return’ effect of feedback on subsequent response latencies so that responses are slower to locations that have just been the site of negative feedback. A three-factor ANOVA with group, previous trial feedback (error versus correct) and previous feedback location (same versus different) was performed on latency for entirely correct responses (see Fig. 3). This allowed us to examine whether there was a bias to make slower responses when the correct saccade was to the side where negative feedback had just been received. There was a significant interaction of location×feedback [F(1, 68)=48.7 p<0.001], indicating slower responses to the location of negative feedback on the previous trial compared with the opposite location. The absence of a feedback×direction×group interaction [F(1, 68)=0.29] indicates that patients and controls showed the same location specific effect of negative feedback in terms of their latencies as the control group.
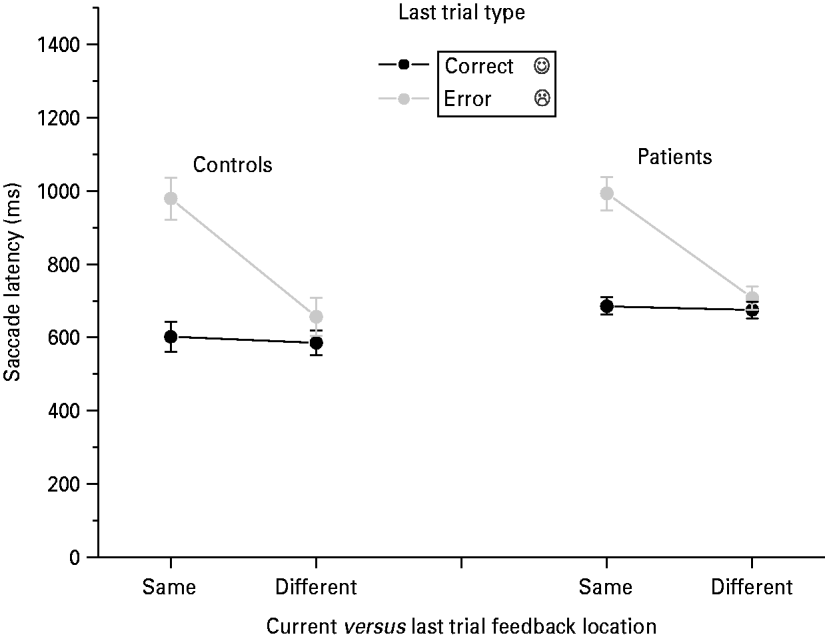
Fig. 3. Effect of relative location of previous feedback on correct trial latencies for low and average pre-morbid IQ patients and control groups.
Reward-related inhibition of return effect on errors
An analysis was also conducted to determine if the bias to avoid making saccades to the location of previous error feedback impacted on the nature of the errors. A three-factor ANOVA was conducted only on the first trial following rule switches to examine the effect of the previous location of feedback (same versus different), error type (corrected versus uncorrected) and group. There were significant main effects of location [F(1, 68)=30.1, p<0.001] and group [F(1, 68)=10.2, p<0.01] but no three-way interaction of feedback, location and group [F(1, 68)<1]. The location×group interaction was significant [F(1, 68)=8.2, p<0.01], whereas the group×error type interaction was not [F(1, 68)=2.6, p=0.11]. Fig. 4 indicates that both groups made more errors of both types by directing their gaze away from the previous location of error feedback but patients showed a significant exacerbation of this effect.

Fig. 4. Effect of relative direction on error rates on rule switch trials for patients and control groups.
In summary, both patients and controls were equally slow to make saccades in the direction of previous negative feedback. Both groups also tended to make errors in this condition by incorrectly looking away from the location of previous negative feedback but patients made significantly more. This latter effect explains the finding of increased trial 1 errors in the patients, which is reported above.
Correlation between rule switching and neuropsychology
To reduce the number of comparisons, corrected and uncorrected errors were collapsed into a total errors score and this was compared only on trial 1, where group differences emerged (see Table 2). There were no strong relationships between errors, spatial working memory or spatial span in patients with schizophrenia. However, the control group showed a strong correlation between spatial span and switching errors. Pre-morbid and current IQ was moderately associated with rule switch errors in patients.
Table 2. Correlations between switch task errors (Sw ers) (trial 1), working memory [spatial span and spatial working memory errors and IQ (WTAR and WAIS)]
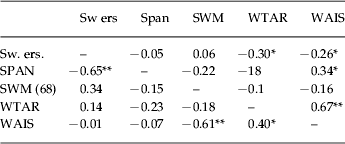
SWM, Spatial working memory; WTAR, Wechsler Test of Adult Reading; WAIS, Wechsler Adult Intelligence Scale.
Correlations above the diagonal are patients with schizophrenia (n=45) and below are controls (n=25).
* p<0.05, ** p<0.01.
The influence of IQ on switching
As patients had a significantly lower IQ than controls, we examined the effect of IQ by extracting a subgroup of patients with an average IQ using a WAIS cut-off score of 90 (n=17); these were matched to the controls on current IQ [patients: mean=100.2, s.d.=4.8; controls: mean=97.7, s.d.=10.4; t(39)=−0.9]. A three-factor ANOVA (group, error type and trial) revealed a significant group×trial interaction [F(5, 200)=3.0, p<0.05], indicating that patients with schizophrenia who have IQ in the average range and equivalent to controls make more errors immediately following rule shifts.
In contrast, there was no error type×group interaction [F(5, 200)<1]. This indicates that the group×error type interaction reported in the full group of patients above may be due to IQ differences between the groups. To determine which factors predicted error correction, a hierarchical multiple regression was conducted with proportion of errors (proportion of all errors that were followed by a corrective saccade within the deadline) as the dependent variable and group and current IQ as predictors. IQ was entered first and group second. IQ explained a significant amount of the variance (R 2=0.20, p<0.01) in the first step but group failed to predict further variance in the second step (R 2 change=0.00, n.s.). This finding suggested that error correction is related to IQ rather than group membership.
In summary, the ability to correct erroneous responses within the time limit was a function of IQ. Patients with a normal IQ, equivalent to that of the control group, made more errors of any type than controls immediately following the rule change.
Correlation between error rates and symptoms
Correlations were conducted between trial 1 errors and the three symptom syndromes. There was a significant moderate correlation between the severity of the negative syndrome and trial 1 errors (r=0.36, p<0.05) but no correlations with positive or disorganization syndromes.
Discussion
In this study, we examined rule switching performance in patients with first episode schizophrenia using an oculomotor task. Patients made more errors and had longer response latencies overall relative to control participants. The increase in errors was particularly marked immediately following a change in the rule. Further examination revealed that this effect was almost completely the result of saccades executed away from the location of a previous negative feedback, i.e. rather than responding on the basis of the new rule, patients with schizophrenia made a saccade to the location opposite to the last response. Further, when patients responded correctly, they showed the same slower response latencies as controls to the location of a recently presented negative feedback (i.e. the so called ‘reward-dependent’ inhibition of return effect described by Hodgson et al. Reference Hodgson, Mort, Chamberlain, Hutton, O'Neill and Kennard2002). Thus, patients with schizophrenia demonstrated abnormalities on a task that required oculomotor switching between rules triggered by symbolic cues. This is in contrast to findings of unimpaired oculomotor switching between reflexive pro-saccades and anti-saccades. We suggest that this difference occurs because the current task involves repeated reconfiguration and retrieval of stimulus response mappings that do not involve a reflexive response.
The finding that errors on this task were particularly related to impaired negative feedback processing has broader implications for understanding the nature of rule and attention switching deficits in other contexts. It is important to emphasize differences in the structure of superficially similar paradigms when comparing studies that measure rule and attention switching in schizophrenia patients. The rule switching tests described, WCST and CANTAB attentional set-shifting tasks, all require participants to update behavioural rules on the basis of response contingent feedback. However, the cognitive operations resulting from feedback may lead to variations from task to task. As with the present task, in the WCST, biasing responses away from the card, which had just been associated with a negative feedback on the last trial, would be a maladaptive strategy. Indeed, a meta-analysis of WCST errors profile in schizophrenia has shown that non-perseverative errors of this type constitute a large proportion of the total errors on the WCST (Li & Park, Reference Li and Park2004), so a similar ‘avoid negative feedback’ response strategy may contribute to the deficit on this task. However, aversive responses away from specific stimuli that have been subject to negative feedback would be adaptive for mediating straightforward response switches between trials or intra-dimensional (ID) attentional switches, explaining why patients show a clearer deficit at the extra-dimensional (ED) shift stage of the ID/ED task (Hutton et al. Reference Hutton, Puri, Duncan, Robbins, Barnes and Joyce1998). Also consistent with this common explanation of deficit in schizophrenia, Waltz & Gold (Reference Waltz and Gold2007) report marked impairments in patients using a probabilistic version of a reversal learning task (Swainson et al. Reference Swainson, Rogers, Sahakian, Summers, Polkey and Robbins2000). In their study, the standard parameters of reversal learning were modified by introducing a probabilistic component, so that erroneous feedback was given on a minority (20%) of ‘correct’ trials. Under these conditions, participants must avoid being influenced by the location of recent negative feedbacks and instead attend to the feedback likelihood over a series of trials to determine the current rule. The marked impairment that patients with schizophrenia demonstrate is again indicative of negative feedback processing deficits in this group.
Our findings therefore suggest that schizophrenia patients have rule switching deficits and that this is mainly due to impaired negative feedback processing. Previous studies of more complex rule switching tasks, such as the WCST schizophrenia, have been unable to clarify whether the impairments are secondary to an abnormally blunted impact of negative feedback or occur because patients fail to use negative feedback correctly to guide behaviour despite appreciating the affective valence (Gold et al. Reference Gold, Waltz, Prentice, Morris and Heerey2008; Prentice et al. Reference Prentice, Gold and Buchanan2008; Leeson et al. Reference Leeson, Robbins, Matheson, Hutton, Ron, Barnes and Joyce2009; Murray et al. Reference Murray, Cheng, Clark, Barnett, Blackwell, Fletcher, Robbins, Bullmore and Jones2008). Our results are more unequivocal in this matter as patients were ultra-sensitive to negative feedbacks, as evidenced by their pattern of errors following a rule change, but failed to update conditional stimulus–response rules as a result of the error. This conclusion agrees with that of Heerey et al. (Reference Heerey, Robinson, McMahon and Gold2007), who showed that patients with schizophrenia have a general difficulty in ‘translating experience into action’.
In this regard, it is important to understand whether our findings can be explained by the IQ difference between our patients and controls. Generalized cognitive impairment is probably an intrinsic feature of schizophrenia (Woodberry et al. Reference Woodberry, Giuliano and Seidman2008) and can explain many of the deficits found on neuropsychological tests (Dickinson & Harvey, Reference Dickinson and Harvey2009). We examined this possibility in a subset of patients matched for current IQ and found they too showed increased errors immediately after negative feedback, explained by the reward-related inhibition of return effect. The main difference was that the average IQ patient corrected errors to the same degree as controls and regression analysis showed that error correction was related to IQ and not to diagnostic group. Thus, in the full group of schizophrenia patients, it can be concluded that lower IQ is related to the inability to self-monitor and correct errors, whereas impaired negative feedback processing is independent of IQ effects.
Rather than reflecting failures to update rules, an alternative explanation for the present results would be that patients' have correct knowledge of task rules but have a weak representation of motoric goals (Hunt et al. Reference Hunt, Olk, von Muhlenen and Kingstone2004; Reuter & Kathmann, Reference Reuter and Kathmann2004), such that pre-potent saccadic responses dominate. In the current context, the aversion to negative feedback is the analogous pre-potent response that dominates saccadic responding. A previous study of the oculomotor rule switching task in patients with frontal lobe lesions (Hodgson et al. Reference Hodgson, Chamberlain, Parris, James, Gutowski, Husain and Kennard2007) reports that increased errors after a rule switch is associated with lesions of the right ventrolateral prefrontal cortex, an area thought to be part of a network involved in inhibitory control of responding, including task switching (Aron et al. Reference Aron, Robbins and Poldrack2004) and response inhibition, particularly stopping (Aron et al. Reference Aron, Fletcher, Bullmore, Sahakian and Robbins2003). We have also previously demonstrated impaired stopping in the same group of patients with schizophrenia (Huddy et al. Reference Huddy, Aron, Harrison, Barnes, Robbins and Joyce2009) and other researchers have shown an attenuation of inferior frontal cortex activity in patients (Kaladjian et al. Reference Kaladjian, Jeanningros, Azorin, Grimault, Anton and Mazzola-Pomietto2007). However, if inhibitory failure was solely responsible for patients' impairment on the task, it would be expected that patients would correct a substantial proportion of their errors (Polli et al. Reference Polli, Barton, Vangel, Goff, Iguchi and Manoach2006) – and the fact that they did not may support the previous suggestion that the abnormality lies in the use of negative feedback in rule abstraction. Another possibility is that the deficit on this task represents both rule updating and inhibitory impairment in schizophrenia.
Another brain region that may be important in attention and rule switching is the orbitofrontal cortex. This region is activated in healthy people and non-human primates during reversal learning tasks. It has been suggested that it serves to maintain a representation of the negative value of stimuli for action selection and in detecting breaches in expected positive outcomes and learning from them (Nobre et al. Reference Nobre, Coull, Frith and Mesulam1999; Tremblay & Schultz, Reference Tremblay and Schultz2000; Takahashi et al. Reference Takahashi, Roesch, Stalnaker, Haney, Calu, Taylor, Burke and Schoenbaum2009). A single case study of a patient with a circumscribed bilateral orbitofrontal cortex excision reported their performance on the same oculomotor rule switching task described here (Hodgson et al. Reference Hodgson, Mort, Chamberlain, Hutton, O'Neill and Kennard2002). It was found that this patient showed a reduction in the magnitude of the reward-based inhibition of return effect, i.e. the opposite effect to that found in patients with schizophrenia in the present study, who made increased errors and whose behaviour was dominated by an overt inhibition of return bias. This would seem to indicate that the orbitofrontal cortex is overactive in patients with schizophrenia, leading to an apparent oversensitivity to negative feedback. However, heightened orbitofrontal activity conflicts with other research indicating underactive orbitofrontal function in schizophrenia (Waltz & Gold, Reference Waltz and Gold2007; Murray et al. Reference Murray, Cheng, Clark, Barnett, Blackwell, Fletcher, Robbins, Bullmore and Jones2008; Leeson et al. Reference Leeson, Robbins, Matheson, Hutton, Ron, Barnes and Joyce2009). Given that rule representation is likely to be mediated by the dorsal frontal cortex and outcome value is an aspect of orbitofrontal function, it may be that dysfunction in schizophrenia is best explained by abnormal interactions between dorsal and orbital frontal cortex (see also Gold et al. Reference Gold, Waltz, Prentice, Morris and Heerey2008), rather than abnormality of the orbitofrontal cortex per se.
These findings can also be viewed as being consistent with putative neurotransmitter dysfunction in schizophrenia. A wealth of evidence points towards abnormal dopamine function in psychosis (Anden et al. Reference Anden, Butcher, Corrodi, Fuxe and Ungerstedt1970; Creese et al. Reference Creese, Burt and Snyder1976; Seeman et al. Reference Seeman, Chau-Wong, Tedesco and Wong1976; Abi-Dargham et al. Reference Abi-Dargham, Rodenhiser, Printz, Zea-Ponce, Gil, Kegeles, Weiss, Cooper, Mann, Van Heertum, Gorman and Laruelle2000). The mesolimbic dopamine system may be recruited to signal breaches in behaviour-outcome predictions, which demand updating of cognitive representations. This dopaminergic ‘prediction error’ signal (Schultz & Dickinson, Reference Schultz and Dickinson2000; Waelti et al. Reference Waelti, Dickinson and Schultz2001) may be disrupted in schizophrenia compared with healthy individuals, contributing to the elaboration of delusional beliefs (Kapur, Reference Kapur2003; Corlett et al. Reference Corlett, Murray, Honey, Aitken, Shanks, Robbins, Bullmore, Dickinson and Fletcher2007). This would fit with the present findings, in which patients show an enhanced behavioural response to negative feedbacks (i.e. outcome prediction breaches).
In summary, the current study demonstrated that patients in the early course of schizophrenia were more sensitive than healthy controls to negative feedbacks in the context of a simple rule switching task. The response profile suggested that while patients had intact appreciation of the negative valence of punishing events, they more often reacted instinctively rather than using the information to update rules that adaptively guide responses. This finding builds on previous reports of impaired negative feedback processing in schizophrenia by further specifying this deficit at the level of integration of negative feedback into a rule set, with intact basic responsiveness to feedback in the immediate context. As well as being consistent with hypothesized neuroanatomical and pharmacological abnormalities in the condition, the findings also support the use of cognitive rehabilitation packages that particularly focus on strategies such as an emphasis on errorless learning and positive feedback (e.g. Wykes et al. Reference Wykes, Reeder, Landau, Everitt, Knapp, Patel and Romeo2007).
Acknowledgements
This paper describes work conducted at the Division of Neuroscience and Mental Health, Imperial College London, London, UK.
This study was supported by Wellcome Trust programme grant 064607 to E.M.J., M.R. and T.R.E.B. and a Wellcome Trust Sabbatical Award to T.L.H. We are grateful to the consultants and nurses of West London and South West London and St George's Mental Health National Health Service (NHS) Trusts for greatly facilitating the study and to Isobel Harrison and Stan Mutsatsa for patient assessments.
Declaration of Interest
T.R.E.B. has acted as a consultant for Servier, Johnson & Johnson and Bristol–Myers Squibb.






