


On 14 April 1911, a team assembled by R.A.S. Macalister (1870–1950), Professor of Celtic Archaeology at University College Dublin, commenced archaeological investigations at Carrowkeel, Co. Sligo, Ireland. Over 18 days, between April and October, they investigated 14 stone cairns (referred to as ‘carns’). They excavated eight of these, with seven revealing internal chambers, containing a combination of cremated and unburnt human and animal bone as well as a range of objects, including bone pins, beads, and pottery (Macalister et al. Reference Macalister, Armstrong and Praeger1912; Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014). The team published their findings in the Proceedings of the Royal Irish Academy the following year (Macalister et al. Reference Macalister, Armstrong and Praeger1912). This publication included a brief report on the human remains by Alexander Macalister (1844–1919), Professor of Anatomy at the University of Cambridge and father of R.A.S. Macalister (Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014; Meehan & Hensey in Reference Meehan and Henseypress). After the 1912 publication, the remains saw no further study until their chance rediscovery in 2001–02 at the Duckworth Laboratory – now the Leverhulme Centre for Human Evolutionary Studies (LCHES) – in Cambridge by Alison Sheridan, Principal Curator at the National Museum of Scotland (Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014; Meehan & Hensey in Reference Meehan and Henseypress). Even thereafter, the collection was not subjected to modern investigative techniques for another decade until the present study was established in 2013 (see Kador et al. Reference Kador, Geber, Hensey, Meehan and Moore2015b).
Given the fact that it had not been worked on throughout the 20th century, the Carrowkeel assemblage presented a unique opportunity to examine the remains using state-of-the-art scientific approaches. More specifically, the present study chose to employ a methodology combining osteological analysis, AMS radiocarbon dating, isotope analysis, and, latterly, also included ancient DNA (aDNA) sequencing to address the following key questions in Irish and European passage tomb research;
1. How does the specific evidence from Carrowkeel relate to population dynamics and burial practices at other passage tombs in Neolithic Ireland and beyond?
2. What is the chronology of activity, and in particular the sequence of depositions, at the complex?
3. What is the demographic profile, genetic ancestry and geographical origin of the people whose remains were placed inside the tombs at Carrowkeel?
After providing a brief background on the Carrowkeel landscape and previous research on the complex, the paper will introduce the analyses conducted on the remains with a view to expanding upon the strategy and methods used for addressing these three questions.
CARROWKEEL IN CONTEXT
Background and location
Carrowkeel comprises a group of megalithic monuments centred on the Bricklieve Mountains, a distinctive karst upland to the west of Lough Arrow in south-east Co. Sligo. There, a closely spaced central agglomeration of cairns (over about 2 km2) is surrounded by a set of outlying monuments, making up a total of 27 passage tomb and probable passage tomb tradition sites within a c. 6 km radius (Table 1; Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014, 11–13, fig. 3), thus covering an area of roughly100 km2 (Fig. 1). Recent literature has considered the most appropriate name for these groups of tombs. Following Bergh (Reference Bergh1995), ‘Carrowkeel–Keshcorran complex’ is most commonly used, as the majority of the cairns are situated around these two elevated zones. The name ‘Lough Arrow complex’ has also been discussed as a way of incorporating other passage tomb tradition sites in the region, including Ardloy, Suigh Lughaidh (Seelewey), and Heapstown (Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014, 7–13). However, for simplicity (and continuity, following the terminology used by the 1911 excavation team) in this publication the terms ‘Carrowkeel’ or ‘Carrowkeel complex’ will be employed.

Fig. 1 Overview map of the Carrowkeel complex and the wider Lough Arrow group of monuments (inset: map of Ireland showing location of Carrowkeel)
Table 1 monuments within the carrowkeel/lough arrow complex
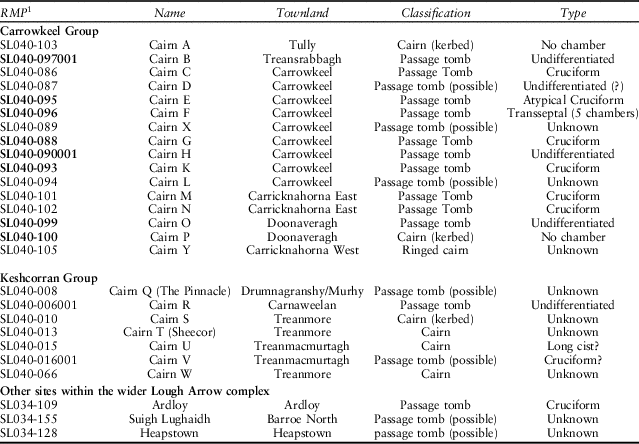
Monuments in bold were excavated by Macalister and colleagues in 1911
1 RMP: Records of Monuments and Places, unique identifier for Irish archaeological monuments
Most of the tombs within the Carrowkeel group have been classed as belonging to the Irish passage tomb tradition. Passage tombs, alongside court and portal tombs, represent one of the three traditionally discussed major Irish Neolithic tomb types (Shee Twohig Reference Shee Twohig1990; Waddell Reference Waddell1998; Cooney Reference Cooney2000). As the morphology of the Carrowkeel tombs is outlined in detail elsewhere (Egan et al. Reference Egan, Byrne, Sleeman, Ronan and Murphy2005), it will not be discussed further here. The core monument cluster, primarily located within the townland of Carrowkeel, contains the 14 sites on which Macalister and colleagues focused, including the eight cairns they excavated. These are perched atop three limestone ridges which grant them a commanding position in the landscape and great visibility from the surrounding lowlands, making Carrowkeel one of Europe’s most striking megalithic landscapes (Fig. 2). Carboniferous Limestone dominates the area surrounding Carrowkeel, however, there are also areas of older (Devonian and Ordovician) geology in the wider north-western region of Ireland (Fig. 3; Harney et al. Reference Harney, Long and MacDermott1996; Holland Reference Holland2003; McAteer & Parkes Reference McAteer and Parkes2004). This is of particular relevance for the interpretation of the results of the strontium isotope analysis.

Fig. 2 Aerial photograph south-west facing of the Carrowkeel passage tomb complex (Courtesy of the Irish National Monuments Service, Department of Arts, Heritage and the Gaeltacht)

Fig. 3 Simplified geological map of County Sligo (Courtesy of the Geological Survey of Ireland). Inset: detail of the Carrowkeel region with the sample locations for the TELLUS Border vegetation samples (V1–4) (graphic: Pádraig Meehan)
Key issues in Irish passage tomb research
The relatively small number of individuals – accounted for archaeologically – from Irish passage tombs, in relation to the size of the communities involved in their construction and the extended duration of activities conducted there, has prompted many discussions regarding their meaning and use (Fleming Reference Fleming1973; Herity Reference Herity1974; O’Kelly Reference O’Kelly1989; Shee Twohig Reference Shee Twohig1990; Bradley Reference Bradley1998; Cooney Reference Cooney2000; Buckley et al. Reference Buckley, Power, O’Sullivan and Thakore2017). In particular, the question concerning who the people chosen for deposition in passage tombs were, and on what basis they were selected, remains a contentious topic. Yet the consensus view seems to be that Neolithic communities were highly selective as to who they interred in their tombs (Cooney & Grogan Reference Cooney and Grogan1994; Cooney Reference Cooney2000; Waterman & Thomas Reference Waterman and Thomas2011).
The paucity of dateable material recovered from Irish passage tombs (Murphy et al. Reference Murphy, O’Donnabhain, Welsh and McGranaghan2010; Buckley et al. Reference Buckley, Power, O’Sullivan and Thakore2017; Cooney Reference Cooney2017), and the historical nature of many investigations, also means that only a limited amount of reliable dating evidence is available. Consequently, the wider chronology of these sites remains a matter of debate (Cooney et al. Reference Cooney, Bayliss, Healy, Whittle, Danaher, Cagney, Mallory, Smyth, Kador and O’Sullivan2011; Bayliss & O’Sullivan Reference Bayliss and O’Sullivan2013; Bergh & Hensey Reference Bergh and Hensey2013; Hensey Reference Hensey2015; Schulting et al. Reference Schulting, Bronk Ramsey, Reimer, Eogan, Cleary, Cooney and Sheridan2017a; Reference Schulting, McClatchie, Sheridan, McLaughlin, Barratt and Whitehouse2017b). In this context, the possibility of obtaining a series of new radiocarbon dates from a major passage tomb complex such as Carrowkeel represents an excellent opportunity to further refine these chronologies.
Given the available burial assemblages from Irish passage tombs, there has been a long-standing perception that cremation was the predominant funerary rite practised at these sites (Herity Reference Herity1974; Cooney & Grogan Reference Cooney and Grogan1994; Cooney Reference Cooney2000). However, this has recently become a matter of debate (Cooney Reference Cooney2014; Reference Cooney2016; Reference Cooney2017); a debate to which the analysis of the Carrowkeel remains can make a significant contribution.
Connecting the bones to the cairns
One cornerstone of this project rested on establishing definitively the relationship between the human remains currently curated at the LCHES, Cambridge and their original context within the Carrowkeel complex, as discussed in the 1912 report (Meehan & Hensey in Reference Meehan and Henseypress). During analysis of the material in 2012 it became apparent that the tomb labelling systems associated with the collection at the LCHES did not match that published by Macalister and colleagues in 1912. Therefore, it was critical to ascertain from which of the Carrowkeel monuments the bones housed at the LCHES originated. A detailed account of the intricate process of unravelling the labelling systems is provided elsewhere (Meehan & Hensey in Reference Meehan and Henseypress) and only the most salient points will be summarised here. It appears that two monument identification systems were employed for the cairns; an alphabetical and a numerical one (Table 2). In the excavation report published in 1912, Macalister and colleagues (1912, 321) describe the alphabetical labelling of 14 individual sites (a system which is still used today). The numerical system, primarily associated with the material curated in Cambridge, applied only to the eight excavated monuments. The initial link between the two systems was provided by a series of photographs made by William A. Green during the team’s second period of excavation at Carrowkeel in June 1911 (Fig. 4). A process of elimination and comparison of the contents of boxes at the LCHES allowed the affinity of the remaining labels to be ascertained to a high degree of probability. They show that the bone material at the LCHES originated from no more than seven (and no less than six) monuments (Table 2). Establishing the relationship between the alphabetic system presented in the 1912 publication and the numeric labels associated with the human remains at the LCHES has been vital to understanding their spatial distribution at the Carrowkeel complex. This in turn has been essential for the analyses and interpretation of the remains described in the following sections.

Fig. 4 Photograph of Cairn F, with R.A.S. Macalister sitting on top, taken by W.A. Green during excavations in June 1911. Note the hand-inked caption reading ‘Carrowkeel No. 4 Cairn’ (Courtesy of the National Museum of Northern Ireland)
Table 2 the numerical & alphabetic labelling systems of the carrowkeel cairns

1 No bone was discovered during excavations of Cairn P in 1911
ANALYSES OF THE REMAINS
Methods and techniques
Four key investigative techniques were brought together to study the remains: osteological analysis; AMS radiocarbon dating; isotope analysis; and aDNA sequencing. The sampling strategies and analytical methodologies employed for each of these methods are outlined in detail in Appendix S1 and will be briefly summarised here. Macroscopic osteological analysis of cremated and unburnt human and animal remains was carried out following standard osteological protocol (Schmid Reference Schmid1972; Buikstra & Ubelaker Reference Buikstra and Ubelaker1994; Brickley & McKinley Reference Brickley and McKinley2004; English Heritage 2014). It focused on demographic (age-at-death and sex), metric (including stature), and taphonomic analysis of both the human and animal bone assemblages (Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2016; Reference Geber, Hensey, Meehan, Moore and Kador2017).
AMS radiocarbon dating was conducted on 38 samples: 29 on human bone (from 27 individuals); eight on cremated red deer antler and one on a canid (dog or wolf) tooth. The non-cremated portion of these samples (n=24) was also analysed for stable carbon and nitrogen isotopes (δ13C and δ15N). The details of these samples, the resulting dates, calibrations, and isotope results are presented in Table 3.
Table 3 summary of samples for dating of human and animal remains

Samples in bold represent double samples (on the same element) for inter- and intra-laboratory comparison and quality control. As the combination of dates for CaK 5_17 showed poor agreement (T=6.5), the sample with the smaller error (SUERC-53151) was used for modelling.
Fifteen human dental enamel samples (from 14 individuals) and one from a canid (see above) were analysed for strontium, oxygen, and carbon isotopes, following published protocols (Kador et al. Reference Kador, Fibiger, Cooney and Fullagar2015a; Chenery et al. Reference Chenery, Pashley, Lamb, Sloane and Evans2012 for oxygen and Triantaphyllou et al. Reference Triantaphyllou, Kador and Nikitae2015 for strontium). To establish a better idea of the (strontium) isotopic background signature in the local biosphere, two samples of crown dentine (Montgomery et al. Reference Montgomery, Evans and Cooper2007; Montgomery Reference Montgomery2010; Ditchfield Reference Ditchfield2014), and four vegetation samples – comprising clippings of twigs – from the Carrowkeel region were analysed for strontium (Fig. 3; see Table 7).
The remains of eight individuals were subjected to ancient DNA (aDNA) extraction and Illumina sequencing, following standard protocols (Yang et al. Reference Yang, Eng, Waye, Dudar and Saunders1998; MacHugh et al. Reference MacHugh, Edwards, Bailey, Bancroft and Bradley2000; Meyer & Kircher Reference Meyer and Kircher2010). With full population genetic analysis still ongoing, the results discussed here are preliminary.
Osteological analysis and human population characteristics
The human bone assemblage from Carrowkeel comprises 9.7 kg unburnt and 5.7 kg cremated commingled remains (Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2016), representing a minimum number (MNI) of 18 unburnt and 22 cremated individuals. Quantified together the cremated and unburnt material give an MNI of 29, an estimate very close to the 31 individuals originally estimated by A. Macalister (Macalister et al. Reference Macalister, Armstrong and Praeger1912). In greater detail they represent a minimum of one neonate (<4 weeks), one infant (1–11 months), two young children (1–4 years), four older children (5–12 years), two adolescents (13–19 years), and 19 adults (≥20 years). This proportion of 35% juveniles/non-adults is higher than that generally found at Irish passage tombs but is virtually identical with the ratio at Knowth (Buckley et al. Reference Buckley, Power, O’Sullivan and Thakore2017; Cooney Reference Cooney2017, 391). Of the adults, nine could be sexed as males and four as females (Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2016, table 2) which is in keeping with a general trend of males seemingly being over-represented at Irish passage tombs (Cooney & Grogan Reference Cooney and Grogan1994; Cooney Reference Cooney2017).
While A. Macalister stated that ‘many of the people buried in the carns [sic] were under twenty-five years of age’ (Macalister et al. Reference Macalister, Armstrong and Praeger1912, 343) the presence of older individuals can be ascertained from progressed stages of attritional tooth wear in at least seven dentitions (three males, one female, and three of indeterminable sex). Additionally, the right half of a male mandible was completely edentulous (9/9), which may indicate an advanced age of that individual with age-related ante-mortem tooth loss. This particular mandible, however, also display a possible healed fracture line located horizontally across the anterior surface of the medial portion of the ramus, just anterior of the right mental foramen, which suggests that some of the tooth-loss was a consequence of direct trauma.
In addition to this case of possible facial trauma, the palaeopathological evidence revealed incidences of joint disease and ossification of ligaments. The spines from a minimum of two individuals (33.33%; 2/6) displayed evidence of degenerative joint disease and osteoarthritis, intervertebral osteochondrosis, vertebral osteophytosis, and Schmorl’s nodes which, in both of these cases, should be regarded as normal wear-and-tear as a consequence of ageing (Gallucci et al. Reference Gallucci, Ongaro, Amici and Regini2009). Among the extra-spinal joints, however, there was a cremated fragment of a scapula from Cairn H that displayed a severe case of joint degeneration of the left shoulder joint. The joint surface of the glenoid cavity was completely worn and displayed patches of eburnation and build-up of marginal osteophytes, which suggests either trauma or excessive repetitive activity as a likely aetiology (Taylor & Laurencin Reference Taylor and Laurencin2009). Other pathological changes relating to activity were indicated from bony enthesophytes on the calcaneal attachment of the Achilles tendon, on a right calcaneus recovered from Cairn K. The Achilles tendon attaches the soleus and gastrocnemius muscles to the heel of the foot, and the pathology suggests that this individual undertook constant pulling on the tendon during plantarflexion of the foot and flexion of the knee joint. This may be as a consequence of repetitive squatting postures, which is further indicated from the primarily platycnemic (laterally flattened) shapes of the measurable tibiae (Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2016). Squatting facets were also present on one left and two right tibiae from the same context, which had already been pointed out by A. Macalister (Macalister et al. Reference Macalister, Armstrong and Praeger1912, 342). No evidence of infectious or metabolic disease was observed in the remains. The only dental pathologies observed were calculus deposits on 81 observable permanent teeth (95.29%; 81/85) in at least 13 dentitions (100.00%; 13/13), ante-mortem loss of ten teeth (2.28%; 10/438) in a minimum of one dentition (8.33%; 1/12), and periodontal disease in five mandibles (41.67%; 5/12).
The people interred in the Carrowkeel monuments appear to conform to a normal variation as expected for an Irish Neolithic population, both in terms of their physical build and skeletal health (Murphy et al. Reference Murphy, O’Donnabhain, Welsh and McGranaghan2010; Roberts & Cox Reference Roberts and Cox2003; Cahill & Sikora Reference Cahill and Sikora2011; O’Donnabhain & Tesorieri Reference O’Donnabhain and Tesorieri2014). Metric analysis indicates that their stature ranged from 154 cm (5 ft ½ in) to 181 cm (5 ft 11 in; Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2016, table 3).
EVIDENCE OF POST-MORTEM BODY PROCESSING
Close macroscopic examination revealed cut-marks on at least 12 bones, deriving from a minimum of one adult individual. The bones were present in Cairns H, K, and from a mixed sample from unknown contexts. They revealed an aspect of the post-mortem funerary rite in the Neolithic which, until this discovery, has rarely been reported from Irish osteoarchaeological evidence, with only two other examples from Carrowmore, Co. Sligo and Millin Bay, Co. Down respectively (Burenhult Reference Burenhult1998; Reference Burenhult2003; Murphy Reference Murphy2003; Murphy et al. Reference Murphy, O’Donnabhain, Welsh and McGranaghan2010). The distribution of the cut-marks suggests that they had been inflicted as part of a dismemberment process rather than defleshing, although the possibility that the latter was also performed cannot be rejected. The joints affected were the shoulder, elbow, hip, knee, and ankle/foot, and the precision and consistency of the cuts indicate that they were undertaken with particular anatomical knowledge and skill that targeted specific tendons and ligaments (Fig. 5; Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2017).

Fig. 5 Examples of human bones with evidence of cut marks (arrows) (a) the neck of a right femur from Cairn K; (b) the base of a right fourth metatarsal from an unknown context; (c) a left calcaneus from an unknown context; (d) a left lateral cuneiform bone from an unknown context (photograph: Jonny Geber)
ANIMAL REMAINS
The skeletal material from Carrowkeel contains a significant proportion of animal remains (Table 4), including six identified mammal species; cattle, caprovine (sheep/goat), pig, red deer, canid (dog/wolf), and hare, as well as a single bone fragment from a gull. The 1911 excavation report mentions the presence of worked bear bone in Cairn G (Macalister et al. Reference Macalister, Armstrong and Praeger1912, 337). While no evidence for bear bone was found during analysis, this comment may refer to a worked bone pin (E624.2), in the National Museum of Ireland, marked in ink with ‘Fibula, Bear, Cairn I Central Recess’. It is, however, not possible to identify this specimen (from Cairn G) to animal species based on morphology alone. Some of the bone fragments from smaller animals in the unburnt material may represent natural inclusions; the presence of cremated animal remains, however, points to their intentional use as part of the burial rite. The most conclusive evidence for this comes from the 39, mainly cremated, large mammal (possibly red deer) bone and antler fragments (Fig. 6). These include an unburnt antler fragment from Cairn E and seven cremated antler fragments and two cremated cranial bones from (the right-hand recess of) Cairn K, which suggests that a complete or partial red deer cranium with antlers still attached may have been placed on a cremation pyre (Serjeantson Reference Serjeantson.2011, 71). The contents of this monument also included a cremated fragment of the femur of a large mammal, which may also be red deer. Additionally, a total of 28 cremated red deer antler fragments were present among the material that could not be attributed to a particular cairn among the seven monuments from which bone is present at the LCHES. These antler samples provide important evidence for the dating of the earliest activity at Carrowkeel, which will be discussed in the next section.

Fig. 6 Fragments of cremated antler from Cairn K (photograph: Jonny Geber)
Table 4 identified faunal remains (fragment count), by species and monument

CR=cremated; UB=unburnt
Chronology
All the 38 samples obtained for dating from the 1911 excavations returned Neolithic dates (Table 3). More specifically, (at 95.4% confidence) they range between 3641–3381 and 2834–2469 cal bc, thus spanning the Middle and Late Neolithic periods (Cooney et al. Reference Cooney, Bayliss, Healy, Whittle, Danaher, Cagney, Mallory, Smyth, Kador and O’Sullivan2011; Whitehouse et al. Reference Whitehouse, Schulting, McClatchie, Barratt, McLaughlin, Bogaard, Colledge, Marchant, Gaffrey and Bunting2014; McLaughlin et al. Reference McLaughlin, Whitehouse, Schulting, McClatchie, Barratt and Bogaard2016). These dates are in good agreement with two recently dated human skull fragments from Cairn G, discovered some 50 years after the 1911 excavations (Table 5; Lucas Reference Lucas1963, 124; Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014, table 5). Somewhat surprisingly, no Bronze Age or later samples were identified among the dated human and animal remains. This is despite evidence for Early Bronze Age bowl and vase tradition pottery in Cairns B, K, and O from the 1911 excavations, as well as Cairn V, excavated in the 1960s (Rynne Reference Rynne1969). There are also two late 3rd millennium bc radiocarbon dates from Cairns M and V (on human tooth and cremated bone respectively; Table 5; Waddell Reference Waddell1990; Bergh Reference Bergh1995; Brindley Reference Brindley2007; Hensey et al. Reference Hensey, Meehan, Dowd and Moore2014).
Table 5 Previously dated samples from Carrowkeel

Samples in bold were included in the chronological models
As the 1911 excavation team left no stratigraphic records, contextualising the recovered remains is problematic. This in turn makes a detailed chronological analysis of the complex difficult to achieve. However, a simple model that assumes all the burials took place as part of a single phase of depositions can be suggested (Fig. 7; Appendix S2; Table S1); including all the dates obtained on human and animal (largely cremated antler) remains from the 1911 excavation and the two samples from Cairn G obtained by Hensey et al. (Reference Hensey, Meehan, Dowd and Moore2014). This model shows overall good agreement with the exception of the earliest (Cak18_RD1806) and the latest samples (CaK1_68). Removing these two ‘outliers’ the model supports one continuous phase of burial activity from 3454–3354 to 2894–2786 cal bc (at 95.4% confidence).

Fig. 7a Chronological model for depositions at Carrowkeel in one continuous sequence (with two outliers)

Fig. 7b
Over this period (460–680 years) one can observe a number of changes in burial practice at the complex. The earliest four dates (including RD1086 which has been excluded from the model) are on cremated antler and, in fact, seven of the eight cremated antler samples are among the ten earliest dates from the complex. Therefore, it seems very likely that the deposition of cremated antlers represents the earliest activity at the complex (see below). Some of the earliest human remains within the Carrowkeel sequence are those samples that show evidence for post-mortem body processing in the form of cut-marks (CaK6_258, CaK15_63, CaK12_730, & CaK14_1270). This suggests that dismemberment was practised during the primary use of the Carrowkeel tombs, between 3403–3026 and 3354–3014 cal bc, with no evidence for bones with cut-marks after 3000 cal bc. Cremated human remains occur broadly contemporaneously with the cut-marked ones, continuing from 3346–3097 to 3012–2887 cal bc. A plateau in the calibration curve at this point (Ashmore Reference Ashmore2004; Schulting et al. Reference Schulting, McClatchie, Sheridan, McLaughlin, Barratt and Whitehouse2017b) prevents further separation of these dates. While deposition of cremated remains may have continued during the 1st century of the 3rd millennium bc, after c. 3000 cal bc unburnt bones and, in particular, skull fragments, appear to predominate. The last 20 samples in the sequence (with the exception of one canid tooth, CaK5_21) are all skull fragments and only two of them show evidence for cremation. It must be added as a caveat that, in order to facilitate isotope and aDNA analyses and to prevent duplicate samples (for details see Appendix S1), the sampling strategy for this project focused to a significant degree on unburnt cranial (mandible and temporal) bones. It therefore cannot be ruled out that interment of complete bodies was undertaken within the monuments and/or that there may be earlier cremated human remains in the assemblage than those identified here.
As an alternative to the above, a purely hypothetical model (ie, not based on archaeological constraints) can be proposed, which sees all the antler depositions – dating from 3641–3382 to 3366–3165 cal bc – preceding any of the human remains (Fig. 8; Table S2). This model – once the outlying earliest (also a cremated antler fragment) and latest dates in the sequence are again excluded – shows equally good agreement with the single sequence of depositions proposed above. While purely hypothetical, the model would present the cremated antlers at Carrowkeel as the earliest phase of activity. It also shows that the cut-marked bones date to the beginning of the sequence of human bone depositions, thus, like the first model, it suggests that the earliest identified human remains deposited at Carrowkeel were subject to dismemberment. There are various reasons, however, why we may not have human bone at dates as early as antler, including the collection strategy of the excavators and the sampling strategy of the current project. As noted above, the majority of the 29 dated human bone samples are on unburnt bone (with approximately 50% on human cranial elements), with only five samples (representing four individuals) of cremated human remains subjected to radiocarbon dating. Thus it cannot be excluded that further dating on cremated human bone may produce results as early as those already obtained from cremated antler.

Fig. 8a Chronological model for depositions at Carrowkeel in two phases (with antlers first and two outliers)

Fig. 8b
While it appears that depositions at Carrowkeel continued throughout most of the second half of the 4th and the first half of the 3rd millennia bc, it is possible to suggest a rough sequence. It commences with the deposition of cremated antler remains (with or without human remains). Thereafter, a diverse range of burial practices can be observed, however, initially focused on the deposition of dismembered human body parts (until c. 3000 cal bc) and of cremated human remains (until c. 2880 cal bc). This appears to be broadly comparable, both chronologically and in terms of mixing of cremated and unburnt remains, with other elaborate Irish passage tombs (Cooney Reference Cooney2017). From the beginning of the 3rd millennium bc there seems to be a shift towards a focus on the deposition of unburnt skull bone, a practice that may present similarities to the evidence from the Mound of the Hostages, Tara, Co. Meath (O’Sullivan Reference O’Sullivan2005; Bayliss & O’Sullivan Reference Bayliss and O’Sullivan2013). An unburnt human cranium, possibly placed in Cairn H, dating to 2834–2469 cal bc (91.5% probability: 2636–2469 cal bc; UBA-30808), represents the latest sample identified as part of this project.
Isotope analyses
The principles of archaeological isotope analysis (of carbon, nitrogen, oxygen, and strontium) have seen considerable discussion in the recent archaeological literature (Bentley Reference Bentley2006; Montgomery Reference Montgomery2010; Schulting Reference Schulting2011; Chenery et al. Reference Chenery, Pashley, Lamb, Sloane and Evans2012; Evans et al. Reference Evans, Chenery and Montgomery2012; Klein Reference Klein2013; Ditchfield Reference Ditchfield2014; Snoek et al. Reference Snoeck, Pouncett, Ramsey, Meighan, Mattielli, Goderis and Schulting2016; Neil et al. Reference Neil, Montgomery, Evans, Cook and Scarre2017). In brief and simplified terms, the ratio between the strontium isotopes 87Sr and 86Sr found in human and animal remains relates to the composition and, especially, the age, of the underlying bedrock geology and the resulting strontium composition in the biosphere (Bentley Reference Bentley2006; Montgomery Reference Montgomery2010). The oxygen ratio (between 16O and 18O expressed as δ18O) on the other hand relates to climate and, in particular, rainfall that is passed on into the cell metabolism through drinking water (Evans et al. Reference Evans, Montgomery, Wildman and Boulton2010). The carbon 13 ratio (expressed as δ13C) relates to diet. Measured on dental enamel carbonate (presented in Table 6), the δ13CC value reflects the blood bicarbonate reservoir and thus the total diet of an individual (Klein Reference Klein2013; Ditchfield Reference Ditchfield2014) whereas when measured on bone collagen (presented in Table 3), δ 13C is more sensitive towards dietary protein (Hedges Reference Hedges2003a; Reference Hedges2003b; Richards & Schulting Reference Richards and Schulting2006). The nitrogen isotope ratio (expressed as δ15N), measured on collagen, is linked to the trophic level (position in the food chain) of the dietary proteins of foods consumed. As human dental enamel is cut off from circulation once the tooth is fully formed, all isotopic values measured on enamel represent the early years of an individual’s life, while the teeth were forming. Therefore they are generally assumed to relate to the location where the individual was born and/or spent their childhood as well as the food and water they consumed during the early years of their lives (Bentley Reference Bentley2006; Montgomery Reference Montgomery2010). In contrast, isotope measurements on bone collagen, which remains connected to circulation until the individual’s death, represent their diet over a number of years prior to this point. However, the turnover rates vary between different bones, with a faster rate in less dense bones such as ribs compared to a slower remodelling rate of the long bones in the arms and legs (Sealy et al. Reference Sealy, Armstrong and Schrire1995).
Table 6 summary of isotope analysis results

1 Sample CaK5_21 was taken from a tooth (right mandibular canine) of a canid (dog or wolf)
2 CaK8_10&11d are crown dentine samples
3 Insufficient sample available of CaK8_12 to measure Sr concentration
Standard errors are as follows: δ13C±0.6; δ18O PDB±0.5; δ18O SMOW±0.6 (all at 1σ)
Tooth samples from 14 humans and one canid were analysed for carbon, oxygen, and strontium (Table 6). These remains, together with an additional ten unburnt human bone samples, were also (dated and) analysed for carbon and nitrogen isotopes on the collagen (Table 3). The carbon analysis of the 14 human enamel samples resulted in an average of –16.9‰ (±0.6) for δ13CC PDB (ranging from –17.6 to –15.2‰). All the 14 values are consistent with a largely terrestrial-based diet, as is generally the case among Neolithic remains from north-west Europe (Richards & Schulting Reference Richards and Schulting2006; Schulting et al. Reference Schulting, Murphy, Jones and Warren2012; Ditchfield Reference Ditchfield2014; Schulting & Boric Reference Schulting and Boric2017) and as could be expected from the inland position of Carrowkeel (located c. 20 km from the coast at Sligo Bay). This is further supported by the collagen δ13C values, which average at –22.1 (±0.9) and range from –23.8 to –21.4‰ and collagen δ15N values, which average at 10.9 (±0.5), ranging from 9.9 to 11.7‰ (Table 3). These values relate to all the 24 dated unburnt bone and tooth samples while, if taken only from the 14 mandibles used for strontium and oxygen analysis, the range for δ13C is narrower (–22.6 to –21.4 ‰) but is unchanged for δ15N. Carbon analysis of the canid tooth enamel (carbonate) returned a value of –11.5‰ (±0.6) δ13CC PDB, which suggests the possibility of a significant marine contribution to the animal’s diet (Laffoon et al. Reference Laffoon, Hoogland, Davies and Hofman2017). However, the stable carbon and nitrogen values (–22.5 and 10.8‰ respectively) on dentine from the same tooth are broadly comparable with those of the human samples and thus do not support this conclusion (Table 3). Additional evidence suggests that this higher δ13CC value may relate to diagenetic alteration (see below).
Strontium analysis of the 14 human tooth enamel samples resulted in an average ratio of 0.7095 (±0.0008), ranging from 0.70863 to 0.71129, while the canid tooth provided a ratio of 0.70860. The four vegetation samples from the surrounding region range from 0.70973 to 0.71194 (Table 7). The two crown dentine samples taken from CaK8_10 and CaK8_11 measured 0.70889 and 0.70842 respectively. The human dentine and canid enamel samples agree well with what could be expected for a Carboniferous environment (Evans et al. Reference Evans, Montgomery, Wildman and Boulton2010; Ryan et al. Reference Ryan, Snoeck, Crowley and Babechuk2018). The higher strontium concentrations of all three of these samples (Table 6) suggests that they have been subject to diagenesis since their deposition at Carrowkeel and are thus more reflective of the burial environment than of lifetime biosphere values of the respective individuals (Fig. 9; Montgomery et al. Reference Montgomery, Evans and Cooper2007). The plant samples, on the other hand, all have strontium ratios above 0.7097 (which is the upper limit for Carboniferous limestone suggested by Ryan et al. Reference Ryan, Snoeck, Crowley and Babechuk2018) and thus highlight the presence of older geological formations in the wider region, in particular to the south of the monuments, where these samples were taken (Fig. 3).

Fig 9 Strontium concentrations & 87Sr/86Sr ratios, showing the diagenetic vectors between enamel & dentine samples, a possible mixing line between 87Sr/86Sr 0.7105 and 0.7086, as well as the two outliers (CaK 8_03 & 09). The dashed horizontal lines demarcate the likely Carboniferous Limestone range. 2σ errors are within the symbols
Table 7 strontium analysis of tellus border vegetation samples from the carrowkeel area

The Easting and Northings are according to the Irish National Grid
The V sample numbers relate to sample locations as shown on Figure 3
All the enamel samples fall between the upper range marked by the plant remains and the lower limit of the dentine samples (Fig. 9). It is thus possible that all the sampled individuals came from within the north-western region of Ireland. However, this represents a relatively broad spectrum of strontium ratios and, while it is difficult to provide a more specific interpretation of the individual samples, it seems probable that a number of different geological backgrounds are represented. This in turn is in line with the relatively diverse geology of north-western Ireland. The majority of samples (n=9) returned ratios between 0.7086 and 0.7097, which can most probably be linked to an area of Carboniferous Limestone (Evans et al. Reference Evans, Montgomery, Wildman and Boulton2010; Ryan et al. Reference Ryan, Snoeck, Crowley and Babechuk2018), similar to the burial environment at Carrowkeel itself. The remaining five individuals with more radiogenic strontium ratios point to origins in areas of older geology, such as the Old Red Sandstone of the Curlew or the metamorphic (largely Granodiorite) geology of the Ox Mountains, c. 10 km south and 25 km north-west respectively (Fig. 3; Harney et al. Reference Harney, Long and MacDermott1996). In particular one sample (CaK8_09) seems considerably higher (2.3σ) than the other 13 and thus suggests a different source of biosphere strontium.
Oxygen analysis of the 14 human tooth enamel samples resulted in averages of: –4.0‰ (±0.5) for δ18O carbonate PDB, ranging from –5.5 to –3.4‰; and 26.8‰ (±0.6) for δ18O SMOW, ranging from 25.2 to 27.4‰. The values for the canid tooth were –5.8‰ (±0.5) δ18O PDB and 25.0‰ (±0.6) SMOW, hence, like the δ13CC, clearly below those measured on the human enamel samples. Given the discrepancies between the enamel carbonate and the collagen δ13C results they need to be treated with caution. Moreover, based on the substantially higher strontium concentration found in the canid tooth compared to the human dental enamel samples (see above and Table 6) it seems probable that diagenetic changes have acted upon the dental enamel of this tooth. While δ18O values measured on the carbonate cannot be directly related to rainwater values (Kohn Reference Kohn1996; Chenery et al. Reference Chenery, Pashley, Lamb, Sloane and Evans2012), based on their oxygen isotope results, an origin within the north-west of Ireland seems probable for most of the samples. All but one of the 14 cluster between 26.2 and 27.4‰ δ18O SMOW (–4.6 to –3.4‰ PDB) and are therefore in line with the expected range of 27.1±1‰ for central Ireland proposed by Cahill Wilson and Standish (Reference Cahill Wilson and Standish2016; Darling et al. Reference Darling, Bath and Talbot2003; Diefendorf & Patterson Reference Diefendorf and Patterson2005). The remaining human enamel sample (CaK8_03) reporting a δ18O value of 25.2‰ (±0.6) δ18O SMOW (–5.5±0.5‰ PDB) is significantly below this range (2.8σ). Cahill Wilson and Standish characterise burials with δ18OC values below 26.0‰ SMOW (–4.7‰ PDB) as ‘favouring an origin outside Ireland, for example in a cooler climate, at a higher latitude or altitude, or further from the Atlantic coast’ (2016, 237). This sample is also one of the five with a strontium ratio above the likely range for Carboniferous limestone, although similar to the average of the vegetation samples.
Thus, as with the strontium ratios, the δ18O values suggest 13 of the 14 individuals from Carrowkeel fit well within what would be expected for the north-west of Ireland region but each of these analyses produced one outlier. Plotting the oxygen against the strontium results for the 12 individuals, excluding these two outliers (CaK8_03 & 8_09), the values appear to be on a diagonal array (Fig. 10). This suggests a mixing line between two different sources (end users); one around δ18O=26.6‰, 87Sr/86Sr=0.7086 and the other around δ18O=27.5‰, 87Sr/86Sr=0.7105 (Montgomery et al. Reference Montgomery, Evans and Cooper2007). At the lower end, the strontium values are consistent with the Carboniferous environment dominating the immediate vicinity of the Bricklieve Mountains, but are also in line with rainwater which, in Atlantic north-west Europe, is similar to seawater (Evans et al. Reference Evans, Montgomery, Wildman and Boulton2010; Montgomery Reference Montgomery2010). The strontium ratios at the upper end appear to reflect the potentially older geology suggested by the plant remains which, for example, could relate to the Boyle, Moygara, and Keadew sandstone formations that make up the Curlew Mountains (Fig. 3; Harney et al. Reference Harney, Long and MacDermott1996).

Fig. 10 Oxygen (on dental enamel carbonate) & strontium isotope results showing a possible mixing line between 87Sr/86Sr 0.7105 and 0.7086, as well as the two outliers (CaK 8_03 & 09). The dashed horizontal lines demarcate the likely Carboniferous Limestone range. Errors indicated for δ18O, 2σ errors for 87Sr/86Sr are within the symbols
Such a mixing line among these 12 individuals is associated with a Pearson correlation of r=0.446. While this is not significant (p=0.127) in itself, this hypothesis can be further substantiated through assessing the correlation between δ13CC and 87Sr/86Sr among these same 12 individuals (Fig. 11). Here the Pearson correlation of r=–0.612 is significant (p=0.026) and even if the two outliers are included the Pearson correlation of r=–0.573 is still significant (p=0.04). Consequently, it seems plausible that the range of strontium ratios from Carrowkeel represents a spread of values between two sources of strontium that are reflected to different degrees in the diets of the 12 individuals. While this is difficult to substantiate further, at the lower end this could reflect the amount of animal produce (meat and dairy) people consumed which are low in strontium, and a more diverse and plant rich diet (higher in strontium) – partially sourced from an area of older geology – at the upper end. This appears to be further supported by Figure 9, which charts strontium concentrations against ratios, showing that the samples with lower strontium ratios also tend to have lower concentrations and vice versa. It is even more striking when comparing strontium and δ13CC values (Fig. 11), showing a clear (and significant) trend from lower δ13C values, among individuals with more radiogenic strontium ratios, towards higher δ13C values, among those with lower strontium ratios (closer to the values for Carboniferous limestone and/or rainwater).

Fig. 11 Carbon (on dental enamel carbonate) & strontium isotope results showing a possible mixing line between 87Sr/86Sr 0.7105 and 0.7086, as well as the two outliers (CaK 8_03 & 09). The dashed horizontal lines demarcate the likely Carboniferous Limestone range. Errors indicated for δ13C, 2σ errors for 87Sr/86Sr are within the symbols
Ancient DNA analysis
Six of the eight samples selected for aDNA analysis yielded usable levels of endogenous human DNA content (8.11–74.87%). The two failed samples (CaK5_16 & CaK17_375) both showed evidence for cremation, which is known to be detrimental to the survival of aDNA (Hansen et al. Reference Hansen, Damgaard, Margaryan, Stenderup, Lynnerup, Willerslev and Allentoft2017). Of the six samples that were successfully shotgun sequenced to higher genomic coverage (Table 8), two were determined to be female and four to be male, which compares relatively well with the 9:4 ratio between males and females identified through osteological analysis. All six samples – five of which came from the same monument (Cairn K) – exhibit genetic affinities similar to those of the Neolithic adult female (3343–3020 cal bc; 4465±38 bp; UB-7059) excavated at Ballynahatty, Co. Down (Cassidy et al. Reference Cassidy, Martiniano, Murphy, Teasdale, Mallory, Hartwell and Bradley2016). Preliminary results suggest their ancestry is derived predominantly from a source closely related to Neolithic Anatolians, as has been observed for their continental counterparts (Hofmanova et al. Reference Hofmanová, Kreutzer, Hellenthal, Sell, Diekmann, Díez-del-Molino, van Dorp, López, Kousathanas, Link and Kirsanow2016), and expectedly they show high similarity to present-day Sardinians. The individuals also exhibit higher levels of allelic sharing with Early Neolithic Iberians relative to Early Neolithic central Europeans. This is consistent with genetic affinities observed for both the Ballynahatty genome and British Neolithic populations, which are argued to be the result of south–north migration and connectivity along the Atlantic seaboard (Cassidy et al. Reference Cassidy, Martiniano, Murphy, Teasdale, Mallory, Hartwell and Bradley2016; Olalde et al. Reference Olalde, Brace, Allentoft, Armit, Kristiansen, Booth, Rohland, Mallick, Szécsényi-Nagy, Mittnik and Altena2018). European Mesolithic ancestry is also detectable in their genomes, at levels similar to the Ballynahatty individual (Cassidy et al. Reference Cassidy, Martiniano, Murphy, Teasdale, Mallory, Hartwell and Bradley2016).
Table 8 preliminary ancient dna results for individuals from the carrowkeel complex

Mitochondrial haplogroup analysis (Table 8) supports these precursory findings, with both haplogroups H and X previously identified as two of the dominant maternal lineages present in early farming populations across Europe. Haplogroup J1c has also been observed in these groups, with J1c3 seen in an Early Neolithic individual from Spain (Haak et al. Reference Haak, Lazaridis, Patterson, Rohland, Mallick, Llamas, Brandt, Nordenfelt, Harney, Stewardson and Fu2015). The mtDNA haplogroup of CaK10_530, W5, appears to be much rarer, both in Neolithic and modern-day European populations (Olivieri et al. Reference Olivieri, Pala, Gandini, Kashani, Perego, Woodward, Grugni, Battaglia, Semino, Achilli, Richards and Torroni2013) and requires further investigation.
It was possible to reconstruct approximately the pigmentation profile of the Carrowkeel population. The presence of the A111T haplotype at the SLC24A5 gene, associated with Western Eurasian light skin colour and fixed in modern European populations, was seen across samples. In addition, both ancestral and derived alleles were observed at the SNP rs16891982 (SLC45A2), the derived allele of which is associated with increased risk of melanoma, lighter skin, hair, and eye colour, and decreases in frequency on a north–south gradient in modern Europe (Norton et al. Reference Norton, Kittles, Parra, McKeigue, Mao, Cheng, Canfield, Bradley, McEvoy and Shriver2007). Estimation of hair and eye colour on a population level (Walsh et al. Reference Walsh, Liu, Wollstein, Kovatsi, Ralf, Kosiniak-Kamysz, Branicki and Kayser2013), based on overall imputed allele frequencies, suggested the most common hair colour in the group was dark brown or black, with brown eye colour dominating, though variation associated with blue eyes was also observed.
Finally, assessment of potential kinship between pairs of samples, with robust inference of up to 4th-degree relatives (Lipatov et al. Reference Lipatov, Sanjeev, Patro and Veeramah2015), detected no very recent relatedness between any of the Carrowkeel individuals, suggesting they represent an outbreeding population. This is supported by preliminary runs of homozygosity analysis, based on imputed genotype data, which reveals no long segments (>8Mb) of homozygosity present in any genome, suggesting none of these individuals is the result of a recent inbreeding event.
Discussion
To make a meaningful investigation of human population dynamics based on the material from Carrowkeel possible it was essential to provide a chronological context for the human remains buried at the complex and link them, to the greatest possible extent, to the monuments from which they were excavated. The first stage in the process was connecting the material at the LCHES to the respective Carrowkeel tombs (Table 2; Meehan & Hensey in Reference Meehan and Henseypress) and providing 38 new AMS radiocarbon determinations mainly on human, but also animal, remains (Table 3). Through these it has been possible to suggest two, somewhat different, chronological models for burial depositions at the complex. The first assumes that all the depositions – with the exception of an outlying early and late sample respectively – took place over one continuous phase (Fig. 7; Table S1). As the earliest dates in the sequence are on cremated antler, an alternative hypothetical model might suggest that the cremated antler remains represent an initial phase predating the (to date identified) human bone remains at the complex (Fig. 8; Table S2). The fact that the earliest dates at Carrowkeel come from cremated antler, whether this constitutes a distinct depositional phase or not, is intriguing and requires further investigation. In particular, it needs to be established whether this could reflect specific burial practices at the tombs (Worley & Serjeantson Reference Worley and Serjeantson2014), or if this apparent phase of discrete deposition of antler without similarly dated human bone may be an artefact of the collection strategy of the 1911 excavations, or of the necessity to primarily sample unburnt bone in this project. The records of Macalister et al. (Reference Macalister, Armstrong and Praeger1912) suggest that antler was found mixed with human bone.
More generally, cremated antler, often in the form of pins, is a relatively common and significant find at Irish passage tombs, suggesting that red deer held important symbolic significance for Neolithic communities. The deposition of antler pins can be directly related to human burials (Herity Reference Herity1974; Eogan Reference Eogan1986; Cooney & Grogan Reference Cooney and Grogan1994; O’Sullivan Reference O’Sullivan2005; Bergh & Hensey Reference Bergh and Hensey2013) and several such pins have been identified at Carrowkeel (Macalister et al. Reference Macalister, Armstrong and Praeger1912). A recent dating project of the Carrowmore passage tomb complex (c. 22 km north of Carrowkeel) was focused on the dating of cremated antler pins (Bergh & Hensey Reference Bergh and Hensey2013). While most of the antler fragments from Carrowkeel are too small to ascertain their role definitively, it is probable that at least some of them once formed parts of pins (Fig. 5) and their dates are virtually synchronous with the date range that seemed to represent the most intensive phase of depositions at Carrowmore (c. 3600–3200 cal bc). Another potentially significant discovery is the identification of possible red deer bone from Cairn K. If this can be further substantiated, it would support the presence of red deer in Neolithic Ireland, rather than the antlers being imported, thus contributing to a long-running debate regarding this question (Woodman & McCarthy Reference Woodman and McCarthy2003; Carden et al. Reference Carden, McDevitt, Zachos, Woodman, Rose, Monaghan, O’Toole, Campana, Bradley and Edwards2012).
The earliest identified human remains from Carrowkeel – though near contemporaneous with cremated human bone samples – are unburnt bones that show evidence for dismemberment. There are no samples with cut-marks post-dating 3000 cal bc and no cremated bone after 2880 cal bc. Consequently, these remains are broadly in line with the recognised Irish Middle Neolithic II (Whitehouse et al. Reference Whitehouse, Schulting, McClatchie, Barratt, McLaughlin, Bogaard, Colledge, Marchant, Gaffrey and Bunting2014; McLaughlin et al. Reference McLaughlin, Whitehouse, Schulting, McClatchie, Barratt and Bogaard2016) and correspond to the period (3300–3000/2900 cal bc) traditionally considered the main phase for ‘developed Irish passage tombs’ (Schulting et al. Reference Schulting, McClatchie, Sheridan, McLaughlin, Barratt and Whitehouse2017b, 14). From around 3000 cal bc, unburnt human cranial bone fragments dominate the assemblage. The dates for these remains are broadly in line with the onset of the recognised Irish Late Neolithic (Cooney Reference Cooney2000; Cooney et al. Reference Cooney, Bayliss, Healy, Whittle, Danaher, Cagney, Mallory, Smyth, Kador and O’Sullivan2011; Whitehouse et al. Reference Whitehouse, Schulting, McClatchie, Barratt, McLaughlin, Bogaard, Colledge, Marchant, Gaffrey and Bunting2014; McLaughlin et al. Reference McLaughlin, Whitehouse, Schulting, McClatchie, Barratt and Bogaard2016). Hence the continued period of depositions after c. 2900 bc, focusing on unburnt crania, marks Carrowkeel out as different from other passage tomb complexes, though the focus on crania shows some potential similarities to the Mound of the Hostages (O’Sullivan Reference O’Sullivan2005; Bayliss & O’Sullivan Reference Bayliss and O’Sullivan2013). The relatively limited amount of Bronze Age finds notwithstanding, they seem to mark the end of prehistoric funerary activity at the complex.
A number of recent high-resolution studies combining palaeolimnological investigation with pollen and geochemical analyses of sediments in two nearby lakes – Templevanny Lough, c. 1 km south-west, and Loughmeenaghan, c. 3 km north of Carrowkeel – have identified evidence for Neolithic farming activity in the catchment areas of both lakes in several key phases (Stolze et al. Reference Stolze, Dörfler, Monecke and Nelle2012; Reference Stolze, Muscheler, Dörfler and Nelle2013; Taylor et al. Reference Taylor, Stolze, Beilman and Potito2016). The studies reveal that the most intensive evidence for mixed arable and pastoral farming dates to the Early Neolithic between c. 3750 and 3500 bc and wanes after this date. This pattern of low levels of farming activity, accompanied by partial woodland recovery, persists throughout both the Middle and Late Neolithic periods, albeit with a brief period of increased small-scale pastoral farming identified at Templevanny c. 2980–2910 bc (Taylor et al. Reference Taylor, Stolze, Beilman and Potito2016, 9). Stolze and colleagues (Reference Stolze, Dörfler, Monecke and Nelle2012; Reference Stolze, Muscheler, Dörfler and Nelle2013) suggest that the changes in agricultural practice may relate to climatic oscillations. At Templevanny they detected a period of climatic downturn and increased rainfall c. 3600–3290 bc, leading to ‘an almost complete woodland recovery between c. 3080 and 3070 bc’ (Stolze et al. Reference Stolze, Muscheler, Dörfler and Nelle2013, 148). While at Loughmeenaghan they date the climatic downturn and recurring periods of increased precipitation to 3520–3170 bc (Stolze et al. Reference Stolze, Dörfler, Monecke and Nelle2012, 400).
The earliest depositions at Carrowkeel, commencing between c. 3600 and 3350 cal bc, appear to coincide with the onset of these worsening weather conditions and a near-cessation of farming activity in the area. Moreover, the brief period of increased pastoral farming in the 30th century bc seems to correspond with the change in burial tradition from a diverse range of practices, including both dismemberment and cremation, to a focus on the deposition of unburnt cranial bones. Thus the evidence from the complex may support the suggestion that changing land-use practices (Taylor et al. Reference Taylor, Stolze, Beilman and Potito2016), declining human activity, and climatic deterioration may have been factors in the expansion of the Irish passage tomb tradition (see Hensey Reference Hensey2015 for discussion). This mirrors significant developments in monument construction elsewhere in Neolithic north-west Europe (Gronenborn Reference Gronenborn2009; Lechterbeck et al. Reference Lechterbeck, Edinborough, Kerig, Fyfe, Roberts and Shennan2014; Downey et al. Reference Downey, Bocaege, Kerig, Edinborough and Shennan2014; Reference Downey, Haas and Shennan2016).
The opportunity to reassess the Carrowkeel remains has provided several additional important insights. Arguably the most significant finding has been the identification of a series of cut-marks on 12 human bones that indicate the post-mortem dismemberment of the dead as part of the burial rite. In their analysis of the assemblage from Coldrum Long Barrow, Kent, England, Wysocki and colleagues suggested that dismemberment of (unburnt) human corpses may point to a variety of meanings, associated with the availability of portable bone and possibly practices such as ‘[n]ecrophagy, shamanism [or] witchcraft’ (2013, 69). Cremation, it could be contended, is a very different process carrying different ritual meanings (Larsson & Stutz Reference Larsson and Stutz2014; Williams Reference Williams2014). At Carrowkeel, however, the dismemberment of unburnt bodies was contemporaneous with the use of cremation. This combination of practices may have represented rites of passage to ‘aid the deceased … in the transition to a state of post-mortem existence’ (Geber et al. Reference Geber, Hensey, Meehan, Moore and Kador2017, 44). Cooney advocates that in an Irish passage tomb context cremation and unburnt burial ‘should not be seen as opposed rites’ (2017, 294). Indeed, the practice of skilled dismemberment on corpses may be relevant to Cooney’s (Reference Cooney2017, 395, 400) proposal that at Knowth, cremated and unburnt bone may, in some cases, derive from a single individual. Notably, A. Macalister appears to have considered the ‘apparent contemporaneity’ of cremated and unburnt bone in his analysis of the remains from the 1911 excavations. In one of his notes, preserved (in box CaK 2) at the LCHES, he observed that burnt and unburnt bone were often encountered mixed together.
More broadly, the significant proportion of unburnt human bone (c. 45% based on MNI) within the Carrowkeel assemblage, runs contrary to the general view of cremation being the dominant funerary rite at Irish passage tombs (Herity Reference Herity1974; Cooney & Grogan Reference Cooney and Grogan1994; Cooney Reference Cooney2000; Buckley et al. Reference Buckley, Power, O’Sullivan and Thakore2017). It is, however, unclear whether this means that the funerary practices at Carrowkeel were significantly different from other Irish passage tombs (Cooney Reference Cooney2017) or whether the higher proportion of unburnt material is largely the result of collection bias during the 1911 excavations and/or favourable preservation of unburnt bone in the karstic environs of Carrowkeel.
Contrastingly, in line with the general view of passage tombs, with a minimum number of 29 individuals (or 40 if counting cremated and unburnt material separately) from seven tombs – in interments that span a period of between five and 11 centuries – it appears that Neolithic communities were selective in choosing who to bury at the complex. This corresponds to a wider trend among megalithic burials in Ireland (Cooney Reference Cooney2017) and north-western Europe more generally (Midgley Reference Midgley2008; Laporte & Scarre Reference Laporte and Scarre2016) and raises the question on what basis people were selected for burial within megalithic tombs. From the evidence at Carrowkeel, it is unlikely that selection was based on sex or age, as the assemblage represents a broad demographic range, with all age groups from infants to older adults and both males and females (albeit at a skewed ratio) being accounted for. The substantial presence (c. 35%) of non-adults in the assemblage, points to all age groups receiving ‘equal treatment’ in death and suggests that children and adolescents were valued members of their communities (Waterman & Thomas Reference Waterman and Thomas2011, 178). The fact that aDNA analysis found no evidence for direct kinship among the individuals whose bones were deposited in the tombs – even though five of the six successful samples came from the same monument – highlights that family groupings were unlikely to have been the basis of selection for placement in the tombs. As an alternative explanation for the low number of identifiable individuals at Irish passage tombs, it could be proposed that tombs may have been regularly cleared of bone (for deposition elsewhere) (Scarre Reference Scarre2011; 185–8; Cooney 2018; 406–7; Schulting et al. 2018, 378).
On a population level, aDNA evidence revealed a genetic profile in keeping with other Neolithic population samples from north-west Europe (Cassidy et al. Reference Cassidy, Martiniano, Murphy, Teasdale, Mallory, Hartwell and Bradley2016; Olalde et al. Reference Olalde, Brace, Allentoft, Armit, Kristiansen, Booth, Rohland, Mallick, Szécsényi-Nagy, Mittnik and Altena2018), with ultimate origins in western Anatolia but also more immediate affinities with remains from the Iberian Neolithic. This seems to reflect the importance of Atlantic seaways in the spread of agriculture and megalithic burial traditions to north-west Europe (Sheridan Reference Sheridan2003; Reference Sheridan2010).
Isotope evidence suggests that the majority of individuals buried at the Carrowkeel complex are most likely local to Ireland’s north-western region. However, two samples differ markedly in their strontium and oxygen ratios respectively, which may point to these individuals having originated from outside the region. Among the remaining 12 individuals a linear relationship between strontium and both carbon and oxygen ratios as well as strontium concentrations suggests that the people buried at Carrowkeel may have sourced their food from two different regions within the north-west of Ireland, highlighting the possibility that this could point to social differentiation, as reflected in their diets.
Conclusions
The multidisciplinary approach employed in the analysis of the burial population at Carrowkeel has served to enhance our current understanding of life and death during an important phase of the Irish Neolithic.
First, the results emphasise the highly complex and varied character of mortuary practices in the Irish passage tomb tradition, which evolved over a sustained period. The study identified the practice of dismembering human remains and showed this to have been conducted contemporaneously with that of cremation. In quantifying the remains it could be established that near equal proportions of the assemblage are represented in the cremated and unburnt material; that male and female adults as well as non-adults are accounted for; and that animals, both wild and domesticated, played important roles in people’s lives and accompanied them in death.
Secondly, through 38 new radiocarbon determinations, the project has provided a chronology for the complex suggesting that its initial use occurs around the middle of the 4th millennium bc. This succeeds relatively closely the earliest phases at Carrowmore (Bergh & Hensey Reference Bergh and Hensey2013) and pre-dates most other passage tomb evidence from Ireland, such as the Boyne Valley tombs (Cooney et al. Reference Cooney, Bayliss, Healy, Whittle, Danaher, Cagney, Mallory, Smyth, Kador and O’Sullivan2011; Bayliss & O’Sullivan Reference Bayliss and O’Sullivan2013; Schulting et al. Reference Schulting, Bronk Ramsey, Reimer, Eogan, Cleary, Cooney and Sheridan2017a). The extended duration of usage of Carrowkeel for the deposition of bone is notable, covering more than half a millennium. The model also indicates a number of changes in burial practice over time, potentially in line with apparent periods of climatic oscillation and changes in Neolithic farming practice, identified more broadly across Ireland (Whitehouse et al. Reference Whitehouse, Schulting, McClatchie, Barratt, McLaughlin, Bogaard, Colledge, Marchant, Gaffrey and Bunting2014; McLaughlin et al. Reference McLaughlin, Whitehouse, Schulting, McClatchie, Barratt and Bogaard2016) and north-west Europe (Whittle et al. Reference Whittle, Healy and Bayliss2011; Lechterbeck et al. Reference Lechterbeck, Edinborough, Kerig, Fyfe, Roberts and Shennan2014; Downey et al. Reference Downey, Bocaege, Kerig, Edinborough and Shennan2014; Reference Downey, Haas and Shennan2016).
Thirdly, a combination of multi-isotope analysis and aDNA sequencing has demonstrated that a relatively diverse population accessed the monuments for ritual and burial purposes over several centuries. They placed within the chambers of Carrowkeel multiple individuals whose deep ancestry was in Anatolia, but who had grown up and lived their lives in the environs of Lough Arrow, the Bricklieve Mountains or regions nearby, as well as a small number of individuals possibly from outside the region. Finally, the isotope research also hints at the possibility that the majority of the individuals identified at Carrowkeel sourced their food from two different areas within the region. This could reflect a level of social differentiation within the community, expressed through access to different food sources, or the presence of distinct populations, originating from several areas within the north-west of Ireland region, among the deposited remains.
Acknowledgements: We would like to acknowledge the kind support from the Leverhulme Centre for Human Evolutionary Studies (LCHES), Cambridge and especially Marta Mirazón Lahr and Maggie Belatti, for granting and facilitating access to the Carrowkeel collection. We also wish to thank the Antiquities Division of the National Museum of Ireland for providing access to the material from Carrowkeel in that collection and the Geological Survey of Ireland for providing vegetation samples from the TELLUS border project. AMS radiocarbon dating was supported through grants from the Prehistoric Society (SUERC), the National Radiocarbon Facility (Oxford), and the Royal Irish Academy (14Chrono Centre). We thus wish to thank all these agencies for their very kind support. Ancient DNA analysis carried out by Lara M. Cassidy was funded by the Irish Research Council Government of Ireland Scholarship Scheme (GOIPG/2013/1219). Additional research support was provided by the Bristol Isotope Group (University of Bristol), the Department of Earth Sciences (Durham University), the British Geological Survey, UCL Earth Sciences as well as the Human Population Genetics Laboratory (Trinity College Dublin). In this context we wish to especially acknowledge the support from Alex Bentley, Colin MacPherson, Daniel Bradley, and John McArthur. We also wish to thank Alison Sheridan for having drawn attention to the existence of the Carrowkeel material at Cambridge; Kevan Edinborough for his advice on chronological modelling; Kerri Cleary for helping with access to materials; Charlotte King and Vesna Lukic for commenting on various drafts of this paper; and the three anonymous reviewers, as well as Julie Gardiner for her editorial advice. Finally, the research formed part of a Marie Skłodowska Curie Research Fellowship (Thomas Kador), funded under the seventh Framework for European Research Innovation Funding (FP7) and we thus wish to acknowledge the support received from the European Research Executive Agency.





















