Introduction
Tomato is reported to be infected by many microorganisms worldwide, including phytoplasmas (Akhtar et al., Reference Akhtar, Saleem, Yousaf, Ullah, Rasool and Sarwar2018). Phytoplasmas are a group of plant pathogenic wall-less phloem inhabiting prokaryotes (El-Sisi et al., Reference El-Sisi, Omar, Sidaros and ElSharkawy2017) naturally transmitted by phloem-feeding insect species of the order Hemiptera (Akhtar et al., Reference Akhtar, Dickinson, Asghar, Abbas and Sarwar2016). Phytoplasmas can also be transmitted through grafting and by vegetative propagation (Omar and Foissac, Reference Omar and Foissac2012). Phytoplasma diseases of tomato have been reported worldwide (Du et al., Reference Du, Mou, Shi, Xu and Xiang2013). The phytoplasma infected tomato plants show different symptoms such as big bud, proliferation of lateral shoots and phyllody (Xu et al., Reference Xu, Mou, Zhu, Liao and Zhao2013).
Breeding tomato cultivars resistant to phytoplasmas or their insect vectors would be an effective tool to manage phytoplasma diseases (Kumari et al., Reference Kumari, Nagendran, Rai, Singh, Rao and Bertaccini2019). However, to date, no information is available about phytoplasma resistance in tomato. Moreover, finding resistance sources requires an appropriate, efficient and reliable screening technique. Recently, we have described a more resource-effective chip grafts inoculation assay (CGIA) for the screening of tomato germplasm against Tomato leaf curl New Delhi virus (Akhtar et al., Reference Akhtar, Akram, Ullah, Saleem and Saeed2019). In this study, we describe the validation of CGIA and resistance potential of different Solanum species against 16SrII-D phytoplasma that will facilitate faster development of disease-resistant cultivars.
Experimental
The 16SrII-D phytoplasma isolate used in this study was described by Akhtar et al. (Reference Akhtar, Saleem, Yousaf, Ullah, Rasool and Sarwar2018) (GenBank accession: LT671581) maintained on tomato variety ‘Nagina’ in a glasshouse at Nuclear Institute for Agriculture and Biology (NIAB), Faisalabad, Pakistan. Plant material comprised 74 genotypes of different Solanum species (Tables 1 and 2). Seeds of each genotype were sown in three large earthen pots, maintained in an insect-free glasshouse at NIAB and plants were thinned to three plants per pot 4 weeks post-germination. At the age of 6 weeks, test plants were inoculated following the CGIA protocol (Akhtar et al., Reference Akhtar, Akram, Ullah, Saleem and Saeed2019). Firstly, a slit of one inch was made one to two inches below the tip on the main stem of the seedlings using a scalpel (Fig. 1(a)). Small pieces of about 2.54–3.81 cm long tender stem/twig from a 16SrII-D phytoplasma-infected source plant were cut and dipped in sterile distilled water. A thin layer of the cell wall (bark) was removed from two opposite sides of each portion of the small piece to expose the cambium layer (Fig. 1(b)). This chip was then inserted into the slit of the test plant (Fig. 1(c) and (d)) and covered with parafilm (Fig. 1(e)). Parafilm wrapping was degraded/removed after 10 d and success or failure of the graft union was assessed visually. A graft was determined to be successful if the inserted chip retained its green colour and had fused to the rootstock at the graft union edges (Fig. 1(f)). Each grafted plant was observed to record disease severity data. Leaf samples were collected 7, 12 and 15 d after grafting to confirm the transmission/presence of the phytoplasma using nested-PCR (nested-polymerase chain reaction). Total DNA was extracted from symptomatic and healthy plant leaves of tomato test genotypes using the Cetyltrimethyl ammonium bromide (CTAB) method (Doyle and Doyle, Reference Doyle and Doyle1990). Nested-PCR was carried out using primer pairs P1 (5′-AGAGTTTGATCCTGGCTCAGGATT-3′) and P7 (5′-CGTCCTTCATCGGCTCTT-3′), followed by R16F2 (5′-ACGACTGCTGCTAAGACTGG-3′) and R16R2 (5′-TGACGGGCGGTGTGTACAAACCCCG-3′) (Deng and Hiruki, Reference Deng and Hiruki1991; Lee et al., Reference Lee, Hammond, Davis and Gundersen1993).

Fig. 1. Different steps involved in CGIA: (a) slit below the tip on the main stem; (b) phytoplasma infected small piece (chip) of stem cut from source plant; (c & d) insertion of chip into the slit of the test plant; (e) slit covered with parafilm; (f) inserted chip fused to the rootstock.
Table 1. Response of different Solanum species against phyllody disease after chip grafting

+++ = Severe disease symptoms; LP = latent period; DS = disease severity; D = determinate; ID = indeterminate; SD = semi-determinate; TGRC = Tomato Genetic Resources Centre, USA; AVRDC = Asian Vegetable Research and Development Centre, Taiwan; ? = not known.
Table 2. Response of different S. lycopersicum genotypes against phyllody disease after chip grafting

+++ = severe disease symptoms; LP = latent period; DS = disease severity; D = determinate; ID = indeterminate; SD = semi-determinate; NIAB = Nuclear Institute for Agriculture and Biology, Pakistan; AARI = Ayub Agricultural Research Institute, Faisalabad, Pakistan; NARC = National Agricultural Research Council, Pakistan; TGRC = Tomato Genetic Resources Centre, USA; AVRDC = Asian Vegetable Research and Development Centre, Taiwan; EFUP = Establishment of facilitation unit for participatory vegetable seed and nursery production programme, Pakistan; GWP = Gujranwala Pakistan; ? = not known.
Discussion
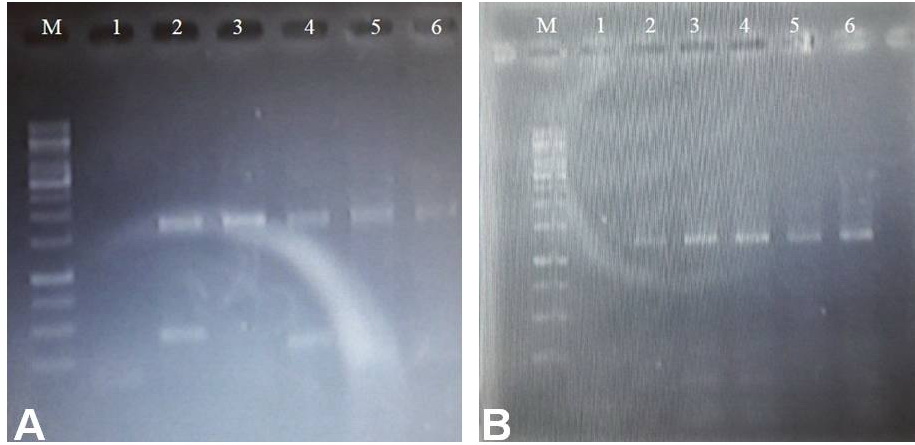
The CGIA successfully transmitted 16SrII-D phytoplasma to all the grafted plants of different Solanum species. Chip grafting and disease transmission were 100%, as all the inserted chips retained their green colour and fused to the graft union edges of rootstock as earlier reported by Akhtar et al. (Reference Akhtar, Akram, Ullah, Saleem and Saeed2019). Symptoms developed from 17 to 30 d post-inoculation (DPI) depending upon the tested genotypes. Parafilm wrapped around the graft was removed 10 DPI. Phytoplasma infection was confirmed in the samples collected 12 and 15 DPI and the nested-PCR product of 1.2 kbp was amplified (online Supplementary Fig. S1). However, phytoplasma was not detected in the samples collected after 7 DPI from the grafted and healthy control plants. None of the grafted genotypes were highly resistant, resistant or tolerant (Tables 1 and 2). After 25–40 DPI, severe phyllody symptoms similar to the source plant were observed in grafted plants (Fig. 1(g)).
Knowledge of the resistance level of tomato germplasm is important in the resistance breeding programmes. However, resistant germplasm identification requires an efficient and reliable screening method to clearly identify their resistance level. Grafting is considered a more standardized and reliable method to transmit viruses and phytoplasmas from infected to healthy plants (Akhtar et al., Reference Akhtar, Akram, Ullah, Saleem and Saeed2019). The CGIA validated under the present study has several potential advantages as it is fast, easy, needs no special expertise, requires much less space, and is sensitive enough to screen tomato germplasm for phytoplasma resistance. The major advantage of the CGIA over other grafting methods is that our developed method allows one infected plant to provide a lot of chips (scions) as compared to a 1:1 ratio of the scion to rootstock in the case of side and cleft grafting techniques. Moreover, a chip used in this study does not require a leaf or bud which is the principle of previously used methods (Akhtar et al., Reference Akhtar, Akram, Ullah, Saleem and Saeed2019). The CGIA, therefore, offers a tool for efficient assessment of resistance to this disease across a range of Solanum germplasm to develop high-yielding and tolerant genotypes/hybrids to limit phytoplasma spread in the field. Also, the CGIA can be used to graft multiple stems or branches of the rootstock plants and even they can be grafted using two different scions. Multiple grafts on single rootstock plants can increase the ‘dose’ of phytoplasma.
Here, for the first time, we have documented the response of a high number of genotypes of different Solanum species against phytoplasma. The results clearly show that no sources of resistance to 16SrII-D phytoplasma are available in the tested genotypes. In future, more genotypes of Solanum species will be screened using the CGIA to find resistant sources, with a view to developing resistant tomato varieties. This assay also has the potential to be extended to better understand the plant–phytoplasma interaction.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262121000137.
Acknowledgements
The authors are thankful to Mr Muhammad Tanvir Elahi, RA, NIAB, Hamna Sadaf SA-I and Mr Zubair Ahmad SA-II for their useful technical assistance.
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical standards
This article does not contain any studies with human participants or animals performed by any of the authors.