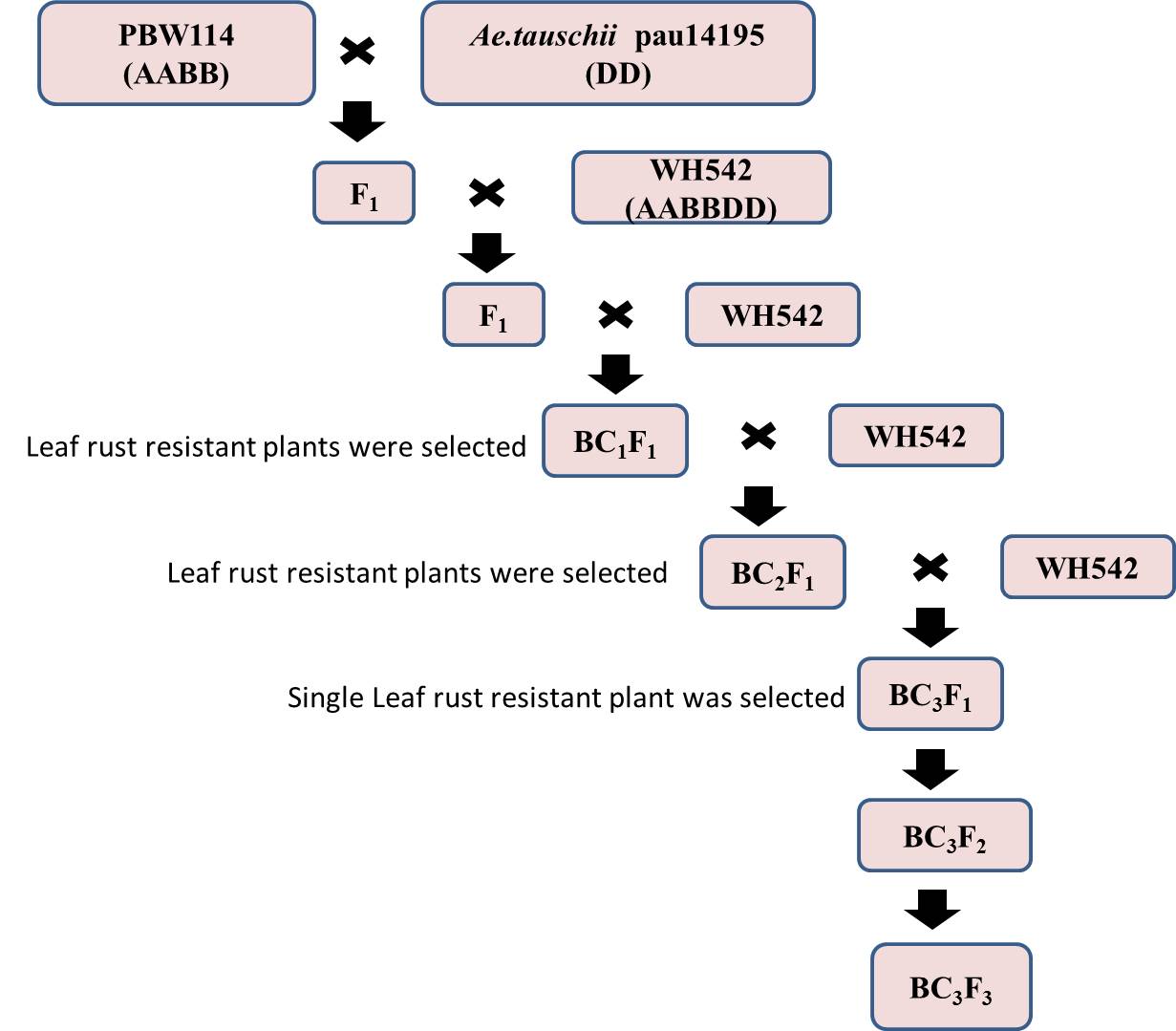
Introduction
Cereal rusts are among the major biotic stresses that pose a substantial threat to wheat production worldwide. Leaf rust caused by Puccinia triticina is one of the most important and devastating foliar diseases of wheat. It causes significant yield losses all over the world due to more frequent and widespread occurrence (Bolton et al., Reference Bolton, Kolmer and Garvin2008). To date, 76 leaf rust resistance genes have been identified and catalogued in wheat (McIntosh et al., Reference McIntosh, Yamazaki, Dubcovsky, Rogers, Morris, Somers, Appels and Devos2013; Bansal et al., Reference Bansal, Kaur, Dhaliwal, Bains, Bariana, Chhuneja and Bansal2017). Continuous emergence of new virulent races and monoculture of few improved cultivars result in increased vulnerability of wheat to this disease. Therefore, it is imperative to stay ahead of the pathogen and search for new resistance genes, not only in the cultivated gene-pool, but also in the wild relatives of wheat (Friebe et al., Reference Friebe, Jiang, Raupp, McIntosh and Gill1996; Valkoun, Reference Valkoun2001).
Cuticular wax deposits are known to reduce the loss of water due to transpiration and hence increasing drought tolerance in plants (Jenks and Ashworth, Reference Jenks and Ashworth1999; Riederer and Schreiber Reference Riederer and Schreiber2001). Glaucousness is a visual trait, which is related to greyish or whitish appearance on the leaves, sheaths, glumes and stems in wheat. This appearance is due to epicuticular wax exudates produced by the plant parts resulting in a waxy bloom on the surface of plant parts. Genetically, glaucousness is controlled by a single dominant gene (W1) mapped to wheat chromosome 2BS. Another single dominant gene, Iw2 located on chromosome 2DS, acts as an epistatic inhibitor to glaucousness resulting in non-glaucous (NG) appearance of plant organs. W1 has a duplicated gene W2 and Iw2 has duplicated gene Iw1, located on chromosome 2DS and 2BS, respectively (McIntosh et al., Reference McIntosh, Yamazaki, Dubcovsky, Rogers, Morris, Somers, Appels and Devos2013), demonstrating that the genes for wax production and suppression are non-allelic (Tsunewaki and Ebona, Reference Tsunewaki and Ebona1999). Iw3, another glaucousness inhibitor gene has been located on chromosome 1BL (Dubcovsky et al., Reference Dubcovsky, Echaide, Giancola, Rousset, Luo, Joppa and Dvorak1997).
Evaluation of wild Triticum and Aegilops species at the Punjab Agricultural University, Ludhiana, India has led to the identification of several Aegilops species including Ae. tauschii as very good sources of resistance to many diseases including leaf rust and stripe rust (Chhuneja et al., Reference Chhuneja, Garg, Kumar, Kaur, Sharma, Bains, Ahuja, Dhaliwal and Singh2010). Present study was conducted to understand the genetic basis of leaf rust resistance and associated glaucousness trait introgressed into hexaploid wheat from Ae. tauschii and to determine the chromosomal location of genes for both traits.
Materials and methods
Development of the mapping population
Ae. tauschii pau14195 was crossed with tetraploid Triticum turgidum ssp. durum cv. PBW114 and F1 was crossed with leaf rust susceptible hexaploid T. aestivum cv. WH542. The complex F1s were further backcrossed with WH542 along with screening for rust resistance at the seedling and adult plant stages (APSs) in each backcross generations. Only the leaf and/or stripe rust resistant BC1 and BC2 plants were further backcrossed. Resistant BC3F1 plants were selfed to generate homozygous introgression lines as well as developing mapping populations. Mapping of a leaf rust resistance gene using BC3F2 and BC3F3 (PBW114/Ae. tauschii acc. pau14195//4*WH542) mapping populations derived from a single resistant BC3F1 plant is being described in the present investigation. Schematic representation of the development of mapping population is given in Figure S1. Populations of 305 BC3F2 plants and 293 BC3F3 families were tested against leaf rust at the seedling and APSs and 160 BC3F2 plants were selected for genotyping.
Evaluation for leaf rust resistance
Screening for leaf rust resistance was done at the seedling stage (SS) and the APS. Parental lines and six randomly selected homozygous resistant BC3F2:3 progenies were tested against 10 different leaf rust isolates at SS. Whole BC3F2 and BC3F3 populations were screened against isolate 77–5 at SS and mixture of races 77–1, 77–5, 104–2 and 77–2 at the APS. At the SS, 305 BC3F2 plants and 10–15 plants of each BC3F2:3 progeny along with parental lines WH542, Ae. tauschii pau14195 and PBW114 were screened following the method of Nayar et al. (Reference Nayar, Prashar and Bhardwaj1997). The landrace ‘Agra Local’ was used as susceptible check. Infection types (ITs) were recorded 14 days after inoculation on a 0–4 scale given by Stakman et al. (Reference Stakman, Stewart and Loegering1962) with slight modifications as given in Roelfs et al. (Reference Roelfs, Singh and Saari1992).
After recording leaf rust data, BC3F2 seedlings were transplanted in the field in 1.0 m rows with row to row distance of 20 cm and plant to plant distance of 10 cm to record adult plant leaf rust response on the same F2 plants. BC3F3 progenies of single F2 plants, however, were simultaneously tested at SS and APS using 10–15 seeds for SS testing and 15–20 seeds for APS testing. In field evaluations, susceptible cultivar Agra Local and WL711 were used as spreader and planted after every 20 rows and all around the experimental plot. Artificial rust epidemic was created by spraying the mixture of uredinospores of leaf rust pathotypes 77–1, 77–5, 104–2, 77–2 every alternate day and keeping infected pots in between the rows. Irrigation was done at regular intervals to maintain high humidity in the field. Terminal disease severity was recorded according to the modified Cobb scale (Peterson et al., Reference Peterson, Campbell and Hannah1948).
Phenotypic characterization of glaucousness
The population also segregated for glaucousness (a waxy layer covering the leaves, stem and spike) as the Ae. tauschii pau14195 was NG, while recipient parent WH542 and bridging species PBW114 were glaucous (G). Glaucousness trait was scored at the APS when the spikes were fully visible. Glaucous plants were greyish green in appearance due to the presence of waxy coat on leaf, stem and ear, while NG plants were totally green. BC3F2 plants and BC3F2:3 progenies were scored for glaucousness (G) and non-glaucousness (NG).
Molecular analysis
Genomic DNA Extraction and bulk preparation
Genomic DNA of WH542, Ae. tauschii pau14195, PBW114 and BC3F2 plants was isolated from the young leaves using the CTAB method (Saghai-Maroof et al., Reference Saghai-Maroof, Biyashev, Yang, Zhang and Allard1994) with some modifications. Equimolar concentration of DNA of 10 phenotypically similar BC3F2 plants, i.e. leaf rust resistant-NG and leaf rust susceptible-glaucous plants were pooled separately to form ‘Resistant-non-glaucous bulk’ and ‘Susceptible-glaucous bulk’, respectively, for BSA (bulked segregant analysis).
SSR analysis
Sixty-eight SSR markers previously mapped on seven D genome chromosomes were selected at a distance of 8–10 cM based on the assumption that leaf rust resistance has been transferred from Ae. tauschii. Initial marker analysis was done at Punjab Agricultural University, Ludhiana, Punjab, though final linkage map generation was done at Plant Breeding Institute, The University of Sydney. PCR was performed in Applied Biosystems master cyclers in 20 µl reaction volumes containing 1× buffer, 60 ng of template DNA, 0.375 µM of each primer, 1.5 mM of MgCl2, 0.15 mM of dNTP and 1 U of Taq DNA polymerase. PCR products were separated on 6% non-denaturing polyacrylamide gels and visualized under UV light in gel documentation system. SSR markers polymorphic between parents and bulks were genotyped on 160 BC3F2 plants. Chromosomal location of the markers was ascertained by genotyping on a set of nulli-tetrasomics Chinese spring lines.
Linkage and statistical analysis
Chi-squared (χ 2) test was used to check goodness of fit of the observed and expected ratios for leaf rust resistance and non-glaucousness in BC3F2 population and BC3F2:3 progenies. Linkage analysis was performed using MapDisto version 7.1 (Lorieux, Reference Lorieux2012) using Kosambi mapping function (Kosambi, Reference Kosambi1943) with a LOD threshold of 3.0.
Results
Characterization of leaf rust and glaucousness
The donor Ae. tauschii accession pau14195 showed high level of resistance against all 10 leaf rust isolates, T. durum cv. PBW114 showed susceptible reaction to six isolates and T. aestivum cv. WH542 showed susceptible infection type to four of the isolates (Table 1). At APS, Ae. tauschii pau14195 and PBW114 displayed resistant leaf rust responses (TR), while WH542 was susceptible with rust severity of 40S-60S. WH542 and PBW114 were glaucous, while Ae. tauschii pau14195 was NG (Figure 1).

Fig. 1. Spikes of parental lines (from left to right) PBW114, WH542, Aegilops tauschii and BC3F2 plants showing segregation for glaucousness trait.
Table 1. Infection types of Ae. tauschii, PBW114 and WH542 and six randomly selected BC3F3 families against 10 different Pt pathotypes

0; = no visible uredinia, ; = hypersensitive flecks, 2 = medium uredinia with necrosis, 3+ = large sporulating uredinia without chlorosis.
In BC3F2, 215 plants were resistant, while 78 plants were susceptible (χ 2 (3:1) = 0.41 non-significant at P = 0.05 and df = 1) (Table 2). Seedling and adult plant reaction of the parental lines and representative BC3F2 plants is shown in Figure 2. Plants which were resistant at SS were also resistant at APS and those susceptible at SS were susceptible at APS. For glaucousness trait the BC3F2 population segregated in a ratio of 216NG: 77 G (χ 2 (3:1) = 0.25 non-significant at P = 0.05 and df = 1) indicating that the trait was controlled by single dominant gene. Two-way segregation indicated that out of the 215 leaf rust resistant plants, 209 were NG and six were glaucous. Out of the 78 leaf rust susceptible plants, 71 were glaucous and seven were NG (Table 2). The single gene control of both the traits was further confirmed in BC3F3 generation as F3 families segregated for single gene for leaf rust resistance in 70HR: 143Seg: 75HS (χ 2 (1:2:1) = 0.18, P > 0.05) at SS and APS and 80 Homozygous Non-Glaucous (HNG): 134Seg: 74 Homozygous Glaucous (HG) (χ 2 (1:2:1) = 1.65, P > 0.05) for non-glaucousness (Table 3). Of the 70 homozygous resistant (against 77–5) BC3F3 families, six randomly selected families were tested against nine additional leaf rust races and found to be homozygous resistant against eight of these races (Table 1). ITs against race 12–5 could not be generated due to unavailability of race. These observations confirmed that resistance gene is from Ae. tauschii as PBW114 give susceptible reaction to races 11, 12–2, 16–1, 104–2 and 106. Predominance of the parental types in F2 and F3 populations indicated genetic linkage between genes controlling leaf rust resistance and non-glaucousness. These genes are being temporarily designated as LrT and IwT, respectively.

Fig. 2. Leaf rust reaction of the parental lines and BC3F2 population at seedling stage (A) and adult plant stage (B) with Agra local as susceptible check. R, Resistant; S, Susceptible.
Table 2. Segregation of leaf rust resistance and glaucousness in the BC3F2 population derived from the cross BC3F1 PBW114/Ae. tauschii acc. pau14195//4*WH542

a Expected genotypes are given in parenthesis.
Table 3. The χ 2 analysis of leaf rust resistance and non-glaucousness segregation in BC3F3 families

χ 2 (1:2:1) (LrT versus Lrt) = 0.18, P > 0.05, χ 2 (1:2:1) (IwT versus Iwt) = 1.65, P > 0.05, χ 2 (1:2:1:2:4:2:1:2:1) (LrT versus IwT) = 431.9, P < 0.0001.
Molecular mapping of LrT and IwT
A total of 56 SSR markers and two EST derived markers were polymorphic between parental genotypes Ae. tauschii pau14195 and WH542 and 15 of these polymorphic markers amplified Ae. tauschii specific introgressions in the bulks. Of the 15 introgressed markers, six markers Xbarc124, Xgdm5, Xgdm35, Xcfd51, Xcau96 and Xte6 from chromosome 2D (Liu et al., Reference Liu, Ni, Peng, Song, Liu and Sun2007; Röder et al., Reference Röder, Korzun, Wendehake, Plaschke, Tixier, Leroy and Ganal1998; Somers et al., Reference Somers, Isaac and Edwards2004) amplified diagnostic polymorphic DNA fragments between the two contrasting bulks. None of the SSR markers selected from chromosome 2A and 2B showed polymorphism between the bulks. These six diagnostic markers were then genotyped on 160 BC3F2 plants.
Genotypic and phenotypic data were combined for mapping leaf rust resistance and non-glaucousness locus of Ae. tauschii. LrT was linked to IwT at a recombination distance of 3.4 cM. LrT was flanked by Xbarc124 and Xcau96 at 0.6 and 1.6 cM, respectively, and IwT was flanked by Xgdm35 and Xte6 at 4.1 and 2.5 cM, respectively with gene marker order as Xgdm5-Xgdm35-IwT-Xte6-Xbarc124-LrT-Xcau96-Xcfd51 generating a partial linkage map of 18.4 cM (Figure 3).

Fig. 3. Partial linkage map of chromosome 2D based on BC3F2 population showing leaf rust resistance gene (LrT) and non-glaucousness gene (IwT) transferred from Aegilops tauschii on the short arm of chromosome 2D. Comparison with the maps published by Liu et al. (Reference Liu, Ni, Peng, Song, Liu and Sun2007) and Sun et al. (Reference Sun, Bai and Carver2009) is also depicted.
Comparative analysis of markers mapped on present map with already published maps (Singh et al., Reference Singh, Franks, Huang, Brown-Guedira, Marshall, Gill and Fritz2004; Somers et al., Reference Somers, Isaac and Edwards2004; Sourdille et al., Reference Sourdille, Singh, Cadalen, Brown-Guedira, Gay, Qi, Gill, Dufour, Murigneux and Bernard2004; Liu et al., Reference Liu, Ni, Peng, Song, Liu and Sun2007; Sun et al., Reference Sun, Bai and Carver2009) showed similarity in positions of markers though some rearrangements were also found. This indicated LrT is introgressed on chromosome 2DS. Linked SSR markers Xgdm5, Xgdm35, Xbarc124 and Xcfd51 were verified for their chromosomal location using Chinese Spring nullisomic-tetrasomic lines for homoeologous group two chromosome (N2AT2D, N2BT2A, N2BT2D, N2DT2A and N2DT2B). SSR marker Xgdm35 found to be present on chromosome 2D only with no allele amplifying on chromosome 2A and 2B. Markers Xcfd51, Xbarc124 and Xgdm5 amplified alleles from all the three homeologous chromosomes, i.e. 2A, 2B and 2D (Figure S2). Segregating alleles mapped in the population, were D-genome specific supporting the evidence that the genes in question are located on wheat chromosome 2D. Moreover, Xcfd51 and Xbarc124 have been physically mapped in the distal region of short arm of chromosome 2D (deletion bin 0.47–1.00) (http://wheat.pw.usda.gov/ggpages/SSRclub/GeneticPhysical/groupe2v2.xls) (Sourdille et al., Reference Sourdille, Singh, Cadalen, Brown-Guedira, Gay, Qi, Gill, Dufour, Murigneux and Bernard2004). These results confirmed the location of LrT and IwT on chromosome 2DS.
Discussion
This study identified and characterized a leaf rust resistance gene that showed linkage with a morphological marker non-glaucousness introgressed from Ae. tauschii into cultivated wheat background. Genetic studies conducted on BC3F2 plants and BC3F2:3 progenies revealed monogenic dominant inheritance for leaf rust resistance and non-glaucousness. T. durum PBW114 used as bridging species in crosses of Ae. tauschii with hexaploid wheat WH542. Since PBW114 was also showed resistance to four leaf rust isolates (77–5, 77–7, 77–8 and 77–10) at SS and mixture of races at APS and but resistance gene identified and mapped in the present study was contributed by Ae. tauschii acc. pau14195. PBW114 is susceptible to six of the 10 leaf rust races tested but selected six BC3F3 families were homozygous resistant against the nine leaf rust isolates. Ae. tauschii accession pau14195 also showed complete resistance against all the races. Difference in infection types of donor Ae. tauschii and derived BC3F3 families against different isolates as shown in Table 1 could be attributed to their complex background generated during three way crosses. Resistance gene from PBW114 might have been lost inadvertently while selecting for resistance plants from backcrosses. Based on molecular mapping and nullisomic-tetrasomic analysis LrT and IwT were located on the short arm of chromosome 2D. The linked markers/marker alleles to these two genes were also D-genome specific. One of the linked marker Xgdm35 did not amplify any allele in lines nullisomic for chromosome 2D, confirming its position on chromosome 2D. Though other markers amplified alleles from all the three homeologous chromosomes but alleles segregating in mapping population were chromosome 2D specific as evident from phenotypic data of six selected BC3F3 families against nine leaf rust isolates. Linkage of LrT with NG phenotype acted as a morphological marker in the present study further supports the conclusion that leaf rust resistance and non-glaucousness is the result of introgression from Ae. tauschii as PBW114 and WH542 are NG and only Ae. tauschii has glaucous phenotype. Thus gene mapped in present study on chromosome 2DS is neither a contribution of A or B genome of durum wheat PBW114 used as bridging species.
A dominant inhibitor of glaucousness derived from Ae. tauschii, Iw2 has been previously mapped on the distal region of chromosome 2DS (Tsunewaki and Ebona, Reference Tsunewaki and Ebona1999). Liu et al. (Reference Liu, Ni, Peng, Song, Liu and Sun2007) mapped a dominant non-glaucousness gene Iw3672 from a synthetic hexaploid wheat in the same region as Iw2 that showed linkage with SSR markers Xbarc124 and Xte6 (0.9 cM) and Xwe6 (1.4 cM) on chromosome 2DS and concluded that Iw3672 and Iw2 were the same. In the present study, IwT mapping at a similar location with Xte6 as the closest marker, indicates that IwT and Iw2 could be allelic or the same genes.
Chromosome 2DS of wheat has many other catalogued leaf rust resistance genes such as Lr2, Lr22a and Lr39. Lr2 is close to the centromere (McIntosh, Reference McIntosh1998) and is not effective against Indian races. LrT is different from Lr2 as it has been mapped at the distal region of chromosome 2DS and is resistant to prevalent races in the Indian Subcontinent. There are two alleles of Lr22 (Lr22a and Lr22b) and both of these confer adult plant resistance. In contrast, LrT exhibited leaf rust resistance at both seedling and APS. Lr39 a seedling resistance gene has been mapped on 2DS and is closely linked to Xbarc124 (Singh et al., Reference Singh, Franks, Huang, Brown-Guedira, Marshall, Gill and Fritz2004; Sun et al., Reference Sun, Bai and Carver2009). LrT being mapped in the same region can be an allele of Lr39.
Marker order Xcfd51-Xcau96-LrT-Xbarc124-Xte6-IwT-Xgdm35-Xgdm5 on 2DS, however, showed an inversion in the present map in comparison to the map of Liu et al. (Reference Liu, Ni, Peng, Song, Liu and Sun2007) used for mapping of Iw3672, Sun et al. (Reference Sun, Bai and Carver2009) for Lr41 (Figure 3) and consensus map of Somers et al. (Reference Somers, Isaac and Edwards2004). LrT was distal to IwT at a recombination distance of 3.4 cM. Marker Xbarc124 mapped at 0.6 cM from LrT, can be used for marker assisted mobilization of this gene to other wheat backgrounds.
Because of evolution of virulent leaf rust races, single disease resistance genes become ineffective rapidly. Therefore, pyramiding of various all-stage resistance genes with APR genes can be a more effective breeding strategy to achieve durable resistance. As there is no reported virulence against LrT in India, it is a good candidate gene for use in breeding programmes. It can be pyramided with other leaf rust resistance genes to achieve durable control of leaf rust. We have identified closely linked flanking markers for LrT that can aid in transfer of this gene to other wheat backgrounds through marker assisted selection for wheat germplasm improvement. Owing to ease of selection, linkage with an easily scorable morphological marker can improve selection efficiency in the breeding programmes. Leaf tip necrosis has been utilized as phenotypic marker for selection of progenies carrying Lr34/Yr18 and Lr46/Yr29 (Singh, Reference Singh1992; Rosewarne et al., Reference Rosewarne, Singh, Huerta-Espino, William, Bouchet, Cloutier, McFadden and Dah2006; Shah et al., Reference Shah, Hussain, Ahmad, Farhatullah, Ali and Ibrahim2011). Lines carrying LrT with and without non-glaucousness can be used in the breeding programmes to transfer this gene to elite wheat backgrounds using selection based on molecular and/or morphological markers.
Supplementary Material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262116000460
Acknowledgements
Financial assistance provided by the University Grant Commission, Government of India, New Delhi, is gratefully acknowledged.
Conflict of Interests
None.