





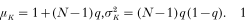
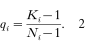
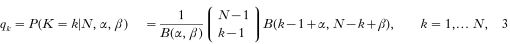

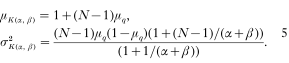















Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Duerr, H.P.
Dietz, K.
Schulz-Key, H.
Büttner, D.W.
and
Eichner, M.
2003.
Density-dependent parasite establishment suggests infection-associated immunosuppression as an important mechanism for parasite density regulation in onchocerciasis.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 97,
Issue. 2,
p.
242.
Brattig, N.W.
2004.
Pathogenesis and host responses in human onchocerciasis: impact of Onchocerca filariae and Wolbachia endobacteria.
Microbes and Infection,
Vol. 6,
Issue. 1,
p.
113.
Duerr, H.P.
Dietz, K.
Schulz-Key, H.
Büttner, D.W.
and
Eichner, M.
2004.
The relationships between the burden of adult parasites, host age and the microfilarial density in human onchocerciasis.
International Journal for Parasitology,
Vol. 34,
Issue. 4,
p.
463.
Duerr, Hans-Peter
Dietz, Klaus
and
Eichner, Martin
2005.
Determinants of the eradicability of filarial infections: a conceptual approach.
Trends in Parasitology,
Vol. 21,
Issue. 2,
p.
88.
Duerr, H.P.
Leary, C.C.
and
Eichner, M.
2006.
High infection rates at low transmission potentials in West African onchocerciasis.
International Journal for Parasitology,
Vol. 36,
Issue. 13,
p.
1367.
Basáñez, María-Gloria
Razali, Karina
Renz, Alfons
and
Kelly, David
2007.
Density-dependent host choice by disease vectors: epidemiological implications of the ideal free distribution.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 101,
Issue. 3,
p.
256.
BOTTOMLEY, CHRISTIAN
ISHAM, VALERIE
COLLINS, RICHARD C.
and
BASÁÑEZ, MARIA-GLORIA
2008.
Rates of microfilarial production by Onchocerca volvulus are not cumulatively reduced by multiple ivermectin treatments.
Parasitology,
Vol. 135,
Issue. 13,
p.
1571.
Duerr, H.P.
Raddatz, G.
and
Eichner, M.
2008.
Diagnostic value of nodule palpation in onchocerciasis.
Transactions of the Royal Society of Tropical Medicine and Hygiene,
Vol. 102,
Issue. 2,
p.
148.
Hoerauf, Achim
Specht, Sabine
Marfo-Debrekyei, Yeboah
Büttner, Marcelle
Debrah, Alexander Yaw
Mand, Sabine
Batsa, Linda
Brattig, Norbert
Konadu, Peter
Bandi, Claudio
Fimmers, Rolf
Adjei, Ohene
and
Büttner, Dietrich W.
2009.
Efficacy of 5-week doxycycline treatment on adult Onchocerca volvulus.
Parasitology Research,
Vol. 104,
Issue. 2,
p.
437.
Bradley, Janette E.
Whitworth, James A.G.
and
Basáñez, Maria-Gloria
2010.
Topley & Wilson's Microbiology and Microbial Infections.
Duerr, Hans P.
and
Eichner, Martin
2010.
Epidemiology and control of onchocerciasis: The threshold biting rate of savannah onchocerciasis in Africa.
International Journal for Parasitology,
Vol. 40,
Issue. 6,
p.
641.
Pion, Sébastien D.S.
Grout, Lise
Kamgno, Joseph
Nana-Djeunga, Hugues
and
Boussinesq, Michel
2011.
Individual host factors associated with Onchocerca volvulus microfilarial densities 15, 80 and 180 days after a first dose of ivermectin.
Acta Tropica,
Vol. 120,
Issue. ,
p.
S91.
Pion, Sébastien D. S.
Nana-Djeunga, Hugues C.
Kamgno, Joseph
Tendongfor, Nicholas
Wanji, Samuel
Njiokou, Flobert
Prichard, Roger K.
Boussinesq, Michel
and
Lammie, Patrick J.
2013.
Dynamics of Onchocerca volvulus Microfilarial Densities after Ivermectin Treatment in an Ivermectin-naïve and a Multiply Treated Population from Cameroon.
PLoS Neglected Tropical Diseases,
Vol. 7,
Issue. 2,
p.
e2084.
Truscott, J.E.
Turner, H.C.
Farrell, S.H.
and
Anderson, R.M.
2016.
Mathematical Models for Neglected Tropical Diseases - Essential Tools for Control and Elimination, Part B.
Vol. 94,
Issue. ,
p.
133.
Bottomley, Christian
Isham, Valerie
Vivas-Martínez, Sarai
Kuesel, Annette C.
Attah, Simon K.
Opoku, Nicholas O.
Lustigman, Sara
Walker, Martin
and
Basáñez, Maria-Gloria
2016.
Modelling Neglected Tropical Diseases diagnostics: the sensitivity of skin snips for Onchocerca volvulus in near elimination and surveillance settings.
Parasites & Vectors,
Vol. 9,
Issue. 1,
Walker, Martin
Hamley, Jonathan I. D.
Milton, Philip
Monnot, Frédéric
Pedrique, Belén
and
Basáñez, Maria-Gloria
2020.
Designing antifilarial drug trials using clinical trial simulators.
Nature Communications,
Vol. 11,
Issue. 1,