












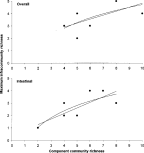
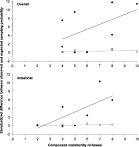


Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
González, M.T.
and
Poulin, R.
2005.
Spatial and temporal predictability of the parasite community structure of a benthic marine fish along its distributional range.
International Journal for Parasitology,
Vol. 35,
Issue. 13,
p.
1369.
Krasnov, Boris R.
Shenbrot, Georgy I.
Mouillot, David
Khokhlova, Irina S.
and
Poulin, Robert
2005.
Spatial variation in species diversity and composition of flea assemblages in small mammalian hosts: geographical distance or faunal similarity?.
Journal of Biogeography,
Vol. 32,
Issue. 4,
p.
633.
Krasnov, Boris R.
Shenbrot, Georgy I.
Khokhlova, Irina S.
and
Poulin, Robert
2005.
Nested pattern in flea assemblages across the host's geographic range.
Ecography,
Vol. 28,
Issue. 4,
p.
475.
GONZÁLEZ, M. T.
and
POULIN, R.
2005.
Nested patterns in parasite component communities of a marine fish along its latitudinal range on the Pacific coast of South America.
Parasitology,
Vol. 131,
Issue. 04,
p.
569.
Krasnov, Boris R.
Stanko, Michal
Miklisova, Dana
and
Morand, Serge
2006.
Habitat variation in species composition of flea assemblages on small mammals in central Europe.
Ecological Research,
Vol. 21,
Issue. 3,
p.
460.
Krasnov, Boris R.
Stanko, Michal
Khokhlova, Irina S.
Miklisova, Dana
Morand, Serge
Shenbrot, Georgy I.
and
Poulin, Robert
2006.
Relationships between local and regional species richness in flea communities of small mammalian hosts: saturation and spatial scale.
Parasitology Research,
Vol. 98,
Issue. 5,
p.
403.
Krasnov, Boris R.
Poulin, Robert
and
Morand, Serge
2006.
Micromammals and Macroparasites.
p.
197.
KRASNOV, BORIS R.
STANKO, MICHAL
and
MORAND, SERGE
2006.
Are ectoparasite communities structured? Species co‐occurrence, temporal variation and null models.
Journal of Animal Ecology,
Vol. 75,
Issue. 6,
p.
1330.
POULIN, R.
2007.
Are there general laws in parasite ecology?.
Parasitology,
Vol. 134,
Issue. 6,
p.
763.
ORTEGO, JOAQUÍN
and
ESPADA, FRANCISCO
2007.
Ecological factors influencing disease risk in Eagle OwlsBubo bubo.
Ibis,
Vol. 149,
Issue. 2,
p.
386.
Krasnov, Boris R.
Shenbrot, Georgy I.
Khokhlova, Irina S.
Mouillot, David
and
Poulin, Robert
2008.
Latitudinal gradients in niche breadth: empirical evidence from haematophagous ectoparasites.
Journal of Biogeography,
Vol. 35,
Issue. 4,
p.
592.
BEHNKE, J. M.
BAJER, A.
HARRIS, P. D.
NEWINGTON, L.
PIDGEON, E.
ROWLANDS, G.
SHERIFF, C.
KULIŚ-MALKOWSKA, K.
SIŃSKI, E.
GILBERT, F. S.
and
BARNARD, C. J.
2008.
Temporal and between-site variation in helminth communities of bank voles (Myodes glareolus) from N.E. Poland. 1. Regional fauna and component community levels.
Parasitology,
Vol. 135,
Issue. 8,
p.
985.
Yang, Kun
Wang, Xian-Hong
Yang, Guo-Jing
Wu, Xiao-Hua
Qi, Yun-Liang
Li, Hong-Jun
and
Zhou, Xiao-Nong
2008.
An integrated approach to identify distribution of Oncomelania hupensis, the intermediate host of Schistosoma japonicum, in a mountainous region in China.
International Journal for Parasitology,
Vol. 38,
Issue. 8-9,
p.
1007.
BEHNKE, J. M.
BAJER, A.
HARRIS, P. D.
NEWINGTON, L.
PIDGEON, E.
ROWLANDS, G.
SHERIFF, C.
KULIŚ-MALKOWSKA, K.
SIńSKI, E.
GILBERT, F. S.
and
BARNARD, C. J.
2008.
Temporal and between-site variation in helminth communities of bank voles (Myodes glareolus) from N.E. Poland. 2. The infracommunity level.
Parasitology,
Vol. 135,
Issue. 8,
p.
999.
TIMI, J. T.
and
POULIN, R.
2008.
Different methods, different results: temporal trends in the study of nested subset patterns in parasite communities.
Parasitology,
Vol. 135,
Issue. 1,
p.
131.
BORDES, F.
and
MORAND, S.
2008.
Helminth species diversity of mammals: parasite species richness is a host species attribute.
Parasitology,
Vol. 135,
Issue. 14,
p.
1701.
BUENESTADO, FRANCISCO J.
FERRERAS, PABLO
BLANCO‐AGUIAR, JOSÉ A.
TORTOSA, FRANCISCO S.
and
VILLAFUERTE, RAFAEL
2009.
Survival and causes of mortality among wild Red‐legged Partridges Alectoris rufa in southern Spain: implications for conservation.
Ibis,
Vol. 151,
Issue. 4,
p.
720.
LAUDISOIT, Anne
LEIRS, Herwig
MAKUNDI, Rhodes
and
KRASNOV, Boris Ronald
2009.
Seasonal and habitat dependence of fleas parasitic on small mammals in Tanzania.
Integrative Zoology,
Vol. 4,
Issue. 2,
p.
196.
Millán, Javier
2009.
Diseases of the Red-Legged Partridge (Alectoris rufa L.): a review.
Wildlife Biology in Practice,
Vol. 5,
Issue. 1,
Krasnov, Boris R.
Matthee, Sonja
Lareschi, Marcela
Korallo-Vinarskaya, Natalia P.
and
Vinarski, Maxim V.
2010.
Co-occurrence of ectoparasites on rodent hosts: null model analyses of data from three continents.
Oikos,
Vol. 119,
Issue. 1,
p.
120.