



Introduction
The shore crab Carcinus maenas (L.) is an abundant epibenthic decapod crustacean species within its native range in the northeastern Atlantic hard and soft shallow coastal areas (Roman and Palumbi, Reference Roman and Palumbi2004), and has a worldwide invasive character (Lowe et al., Reference Lowe, Browne, Boudjelas and De Poorter2000; Darling et al., Reference Darling, Bagley, Roman, Tepolt and Geller2008). It is considered an ‘ecosystem engineer’ because of its ability to modify entire ecosystems (Klassen and Locke, Reference Klassen and Locke2007; Garbary et al., Reference Garbary, Miller, Williams and Seymour2014). Therefore, it has been the focus of several studies, recently reviewed by Young and Elliott (Reference Young and Elliott2020). More recently, the seasonality in the body and energetic condition of shore crabs was investigated in the western Dutch Wadden Sea (Campos et al., Reference Campos, Ribas, Bio, Freitas, Souza and van der Veer2021) showing fluctuations related to environmental constraints such as thermal and prey availability patterns, and with the growth and reproduction of the crabs.
The rhizocephalan barnacle Sacculina carcini is a common endoparasite of the shore crab. Parasite prevalence can reach near 80% of the crab population with a great spatio-temporal variation (Lützen, Reference Lützen1984; Mathieson et al., Reference Mathieson, Berry and Kennedy1998; Torchin et al., Reference Torchin, Lafferty and Kuris2001; Werner, Reference Werner2001; Zetlmeisl et al., Reference Zetlmeisl, Hermann, Petney, Glenner, Griffiths and Taraschewski2011). The life cycle of the parasite and its development on the shore crab host has been previously described (Høeg et al., Reference Høeg, Glenner, Shields and Rohde2005). Shortly, infection occurs by female cypris larvae of S. carcini which settle on the carapace of recently moulted crabs and penetrate the exoskeleton into the haemolymph (Glenner and Werner, Reference Glenner and Werner1998; Glenner et al., Reference Glenner, Høeg, O'Brien and Sherman2000). The parasite grows a root-like system (interna) throughout the host tissues, which serves as the nutrient-absorbing organ (O'Brien and Van Wyk, Reference O'Brien, Van Wyk and Wenner1985). The infected host soon becomes a parasite genotype with a crab phenotype (O'Brien and Van Wyk, Reference O'Brien, Van Wyk and Wenner1985). Later, an external reproductive sac-like structure (externa) produced by the parasite emerges out of the abdomen of the crab, in the brood chamber of the host (Glenner and Høeg, Reference Glenner and Høeg1995; Høeg, Reference Høeg1995), and is fertilized by 1–2 cryptic dwarf males (Delage, Reference Delage1884). After releasing the nauplii larva, the externa falls off leaving a scar on the abdomen of the host crab (Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018).
Sacculina infection severely impacts the general condition of the crab host (Mouritsen and Jensen, Reference Mouritsen and Jensen2006; Larsen et al., Reference Larsen, Hoeg and Mouritsen2013), affecting growth, behaviour, morphology and physiology (Thresher et al., Reference Thresher, Werner, Høeg, Svane, Glenner, Murphy and Wittwer2000), feminizes males, and increases (doubles) the mortality rate of infected crabs (Goddard et al., Reference Goddard, Torchin, Kuris and Lafferty2005). Externally infected crabs (often denominated as ‘sacculinized’ crabs) stop moulting (and hence cease growth) (Høeg, Reference Høeg1995), which favours the settlement and growth of fouling organisms, such as epizoic barnacles, on the carapace of crabs (Mouritsen and Jensen, Reference Mouritsen and Jensen2006). Aside from morphological and behavioural feminization of male hosts (Rasmussen, Reference Rasmussen1959; Glenner and Høeg, Reference Glenner and Høeg1995; Høeg, Reference Høeg1995; Werner, Reference Werner2001), S. carcini also causes the degeneration of the gonads (Day, Reference Day1935; Zetlmeisl et al., Reference Zetlmeisl, Hermann, Petney, Glenner, Griffiths and Taraschewski2011), inducing ‘parasitic sterilization’ in both host genders (Høeg and Lützen, Reference Høeg and Lützen1995; Werner, Reference Werner2001; Zetlmeisl et al., Reference Zetlmeisl, Hermann, Petney, Glenner, Griffiths and Taraschewski2011).
Rhizocephalans can deplete energy stores of the host by utilizing the energy for their development or by causing the host to divert energy into a defence response against infection. Therefore, one would expect that infected crabs have lower energy content than uninfected crabs of similar size. However, it is not clear how C. maenas handles the extra energy requirements of infection. Although some morphological changes induced by Sacculina, such as the size reduction of walking legs (Kristensen et al., Reference Kristensen, Nielsen, Jørgensen, Mouritsen, Glenner, Christensen, Lützen and Høeg2012) and chelipeds of feminized males (Høeg, Reference Høeg1995; Mouritsen and Jensen, Reference Mouritsen and Jensen2006; Lafferty and Kuris, Reference Lafferty and Kuris2009; Kristensen et al., Reference Kristensen, Nielsen, Jørgensen, Mouritsen, Glenner, Christensen, Lützen and Høeg2012) can impact the crabs’ feeding behaviour, host food consumption appears to be unaffected by infection (Larsen et al., Reference Larsen, Hoeg and Mouritsen2013). The energy requirements of the parasite should then match the energy which the crab would allocate for its own growth and reproduction (Larsen et al., Reference Larsen, Hoeg and Mouritsen2013). If so, the nutritional status, including the energy content of the host (i.e. parasite included) would be identical to that of uninfected crabs of similar size.
This is the second of a series of 3 papers dedicated to a comprehensive investigation of the shore crab's condition in Europe's largest coastal wetland system, the western Dutch Wadden Sea. The aim of the present study was to evaluate the impact of Sacculina infection on the general condition of the shore crab C. maenas, more precisely on the condition of the pair ‘crab-parasite’ (sacculinized crab) as they cannot be physically separated. For that, we compared the nutritional status between sacculinized (i.e. parasite rootlets included) and non-sacculinized crabs, using both morphometric indices – the Fulton's K condition index and the percentage of dry weight – and biochemical data – the body energy content.
Material and methods
Sampling and laboratory procedures
A monthly sampling programme was carried out from August 2012 to March 2014 (except in September 2012 and 2013, October 2013, January and February 2014, due to adverse weather conditions) at 3 sites in the Marsdiep tidal basin in the western Dutch Wadden Sea (Fig. 1) as described in Campos et al. (Reference Campos, Ribas, Bio, Freitas, Souza and van der Veer2021). Shortly, a 2 m beam trawl with a single tickler chain and a mesh size of 1 × 1 cm was towed by a scientific vessel (RV Stern) at a speed of approximately 60 m min−1. Each haul covered a surface area of at least 900 m2. All sites were 5–12 m deep and had soft bottom sediments. In addition, water temperature and salinity were recorded at each site before the hauls. All catches were sorted immediately and the shore crabs were transported alive to the lab. In the 2012–2014 dataset, most parasite externa samples were below the minimum required weight for the calorimetry. Therefore, later, in May 2019, an additional sampling focused on obtaining the missing information on the energy content of the parasite externa was carried out to complement the available dataset.

Fig. 1. Map of the sampling locations in the western Dutch Wadden Sea. (1) Gat van der Stier (N 52°57.27′ E 4°55.73′); (2) Texelstroom (N 53°02.03′ E 5°03.37′); and (3) Kornwerderzand (N 53°04.52′ E 5°16.55′); top left: sampling area in the Netherlands. (Adapted from Katwijk and Hermus, Reference Katwijk and Hermus2000).
All crabs were sexed according to the sexual dimorphism traits described in Squires (Reference Squires1990) (male with a triangular abdomen and 3rd–5th somites fused; female with a subtriangular rounded lateral abdomen and somites not fused), and measured to the nearest mm carapace width (CW) with a digital calliper. Each individual was classified into colour morphotype (green or red) based on the predominant colour of the thoracic sternum (McGaw and Naylor, Reference McGaw and Naylor1992), and inspected for the presence of eggs in females, epibiont fouling, and parasites. Fouled crabs were excluded from the study to avoid extra confounding factors affecting energy determinations. Parasites were detected by lifting the abdomen of each crab from the thorax to check for the presence of Sacculina carcini externa (gonad mass sac). Secondary features including scars of lost externa and broad abdomen in males were used to determine past infection and modified male (i.e. feminized) condition, respectively. Prevalence of infection was defined as the percentage of crabs that were externally parasitized (i.e. sacculinized crabs); exclusively internal (i.e. immature) phase of infection was not determined, nor were scarred nor modified male crabs without externa included in the calculation of prevalence, following previous studies (e.g. Zetlmeisl et al., Reference Zetlmeisl, Hermann, Petney, Glenner, Griffiths and Taraschewski2011).
Every month, for each sex, subsets of about 10 uninfected and 10 infected crabs (randomly chosen) per 10 mm size classes were selected for subsequent morphometric and calorimetric analyses. All animals were weighted (WW, wet weight), eviscerated (i.e. dissected to remove the stomach) and weighted again right after, to avoid mixing of stomach content in the calorimetric analyses. The externa of S. carcini was removed from infected crabs before weighing. Since S. carcini creates an internal root system in the whole body of the host, it was impossible to completely remove the parasite and exclusively analyse the host; thus the parasite roots had to be included. This means that any quantitative difference between non-sacculinized and sacculinized crabs is a conservative estimate. Therefore, comparative analyses were made between ‘sacculinized’ (i.e. crabs infected with S. carcini and presenting an externa sac) and ‘non-sacculinized’ (i.e. crabs presenting no externa, which can be uninfected or exclusively internally infected crabs) animals. For the 2019 samples, the externa of sacculinized crabs and the egg mass of ovigerous females were dried and analysed separately. Samples (crab whole body excluding stomach, eggs and S. carcini externa) were individually dried for 10 days at 60°C till constant dry weight (DW). The crabs Fulton's condition index (K) was determined by dividing the WW by the cubic CW.
Calorimetry
The energy content was determined using an IKA C2000 basic Calorimeter the same way as in Campos et al. (Reference Campos, Ribas, Bio, Freitas, Souza and van der Veer2021). After maceration of each dried crab, the powder sample was pressed in a mortar to build a pellet and taken into the calorimeter bomb, where it was combusted and analysed for caloric content (cal g−1 DW). Subsequently, values were corrected for minerals and water remains that did not account for the energy content of the animal. Sample ash contents higher than 30% make it difficult to obtain a complete combustion in the calorimeter (Cummins and Wuycheck, Reference Cummins and Wuycheck1971). In C. maenas, the inorganic material consists largely of calcium (Adelung, Reference Adelung1971), and makes up a great part (about 40%) of the dry weight, affecting the calorimetric result since calcium reaction in the calorimeter is endothermic (Topley et al., Reference Topley, Hume and Hartley1928). Therefore, the ash content was first determined by weighting the remaining ash after burning in the calorimeter to determine the calcium concentration in the samples. Then the ash was re-burned at 900°C in a muffle to remove the minerals that could not be removed during the calorimetric burning process and the remaining was weighed again to determine the ash-free dry weight (AFDW). The difference was used to calculate the percentage of calcium, which was then used to correct the energy content as follows:

where 1.4 cal g−1 is the caloric value of carbonate calcium in calorimetric reactions (Paine, Reference Paine1964). Caloric content was also determined separately for the externa and the egg mass of crabs from the 2019 samples. Finally, caloric values were converted to kJoules per gram of DW (kJ g DW−1), hereafter E DW, per gram of AFDW (kJ g AFDW−1), hereafter E AF, and total energy content (E tot) of each crab was obtained multiplying E DW by the respective DW.
A total of 617 crabs were analysed for energy, of which 114 crabs were externally infected with S. carcini (Table 1). No scarred crab was analysed for energy.
Table 1. Number of sacculinized Carcinus maenas analysed for energy condition per sex (F: females, M: males, MM: modified males) and colour and percentage (%) of total sacculinized sample

Data analysis
The crab condition was analysed for non-sacculinized and sacculinized crabs, considering crab sex and size, and comparing their mean values using Welch's t-test, a modification of the t-test. This test adjusts the number of degrees of freedom when the variances are thought not to be equal to each other and performs better than Student's t-test whenever sample sizes and variances are unequal between groups, as is the case here, providing the same result when sample sizes and variances are equal (Delacre et al., Reference Delacre, Lakens and Leys2017). The significance level was set at α = 0.05.
The effect of Sacculina external infection was also assessed through regression modelling. A previous study, with C. maenas from the Dutch Wadden Sea, carried out on apparently uninfected crabs (not bearing the parasite externa), showed that crab sex and size, and sampling season were the most relevant predictors for crab condition, whereas crab colour was a negligible, as well confounding variable (being related to the animal's size), and sampling site was only relevant for %DW (Campos et al., Reference Campos, Ribas, Bio, Freitas, Souza and van der Veer2021). Therefore, these variables (crab colour and sampling site) were not considered in the models of the present study.
Here, the objective was to obtain insight into external infection effects on condition, independent of the sampling season or site, knowing that samples were not spatially and temporally independent. Furthermore, in the available data set, sampling season is a non-continuous variable (data collected in 2012–2014 and in 2019) with gaps, and sampling sites were sampled in an unbalanced way, containing data from different seasons, crab development stages and, thus, sizes. Therefore, linear mixed effect models were applied, with Sacculina external infection, crab size and sex as fixed factors, and sampling season and site as random factors. This way, the model accounted for temporal and spatial variability, producing better estimates for the fixed effects parameters. Crossed LMER with random intercept and slope were applied (using the R lme4 package; Bates et al., Reference Bates, Mächler, Bolker and Walker2015), with CW standardized to mean zero (i.e. centred) and a standard deviation of 1 (i.e. scaled), to ensure that the estimated coefficients are all on the same scale, to allow comparison of effects. Model explained variance was obtained calculating the marginal R 2, which represents the variance explained by the fixed effects, and the conditional R 2, which is interpreted as the variance explained by the entire model, including both fixed and random effects (according to Nakagawa et al., Reference Nakagawa, Johnson and Schielzeth2017), using the R package stargazer (Hlavac, Reference Hlavac2018).
All data processing and statistical analyses were performed in R (R Core Team, 2020).
Results
General patterns of crab infection
A total of 11 068 shore crabs were collected and analysed for general biometry in the 2012/2014 sampling, from which 3.3% presented a visible externa and therefore were considered as sacculinized crabs (externally infected by Sacculina carcini). Additionally, 0.7% of the non-sacculinized crabs had a visible scar from lost externa on the abdomen. Distribution patterns of prevalence per sex, colour type, size class and season are presented in Fig. 2. Most sacculinized crabs were males (61.7%), of which more than half (60.0%) were modified males. No egg-bearing female was found externally infected or scarred, and non-sacculinized modified males represented only 0.24% of the total sample. In relation to the colour morphotype, despite the majority of the sacculinized crabs were green (57.1%), externally infected green morphs represented only 2.3% of all sampled green crabs, while for red morphs, sacculinized crabs represented 7.9% of all sampled red morphs. The size of externally infected crabs ranged from 14.8 to 88.6 mm, with an average of 40.9 ± 14.0 mm CW, but external infection was higher in adult crabs >25 mm CW, with almost half of the sacculinized crabs ranging from 25 to 40 mm CW (49.5% of the sacculinized crabs) and lowest in juveniles (<25 mm; 7.6%) and in crabs larger than 55 mm (10.3%) (Figs 2 and 3). Distribution of sacculinized crabs followed a seasonal pattern; abundance was highest in spring (38.0%) and autumn (32.6%), and lowest in summer (13.9%).

Fig. 2. Percentage distribution of the abundance of sacculinized crabs according to sex (F: females, M: males, MM: modified males), colour morphotype (G: green, R: red), size class (Juv, juveniles: ⩽25 mm CW; A1: 25–40 mm CW; A2: 40–55 mm CW; A3: >55 mm CW) and seasons.

Fig. 3. Size distribution of sacculinized and non-sacculinized crabs per sex (F: females, upper panel; and M: males and MM: modified males, lower panel). Juv, juveniles: ⩽25 mm CW; A1: 25–40 mm CW; A2: 40–55 mm CW; A3: >55 mm CW. Note that MM non-sacculinized crabs represented only 0.24% of the sampled population.
Model results
Table 2 presents the linear mixed effects model (GLMM) results. The models fitted particularly well for the E tot condition (R 2 > 80%), followed by Fulton's K (R 2 up to 69%), while for E DW, E AF and %DW the R 2 of the model was relatively low (R 2 < 30%). The results suggested non-significant impacts of Sacculina external infection on the morphological condition (K and %DW) and on total energy (E tot), and significant impact of external infection on the energy density (E DW and E AF). Other relevant factors affecting condition were sex and size. According to the GLMM models no predictor variable was significant for the variance found in %DW, only CW for K and E tot, and only Sacculina infection for E DW and E AF.
Table 2. Linear mixed effects model results, with predictor estimate and estimate error, t-value, significance (n.s. non significant, *P < 0.05, **P < 0.01, ***P < 0.001) and the proportion of variance explained by the fixed effects (marginal R 2) and by the entire model (conditional R 2)

Impact of Sacculina external infection on crab's condition
Although models did not reveal a significant impact of external infection on the morphological condition of crabs, sacculinized animals had slightly higher mean Fulton's K (1.42 ± 0.52 and 1.60 ± 0.64, respectively in non-sacculinized and sacculinized crabs; P value <0.01, Welsh's t-test), especially females (1.53 ± 0.56 and 1.82 ± 0.89, respectively in non-sacculinized and sacculinized crabs; P value = 0.051) and modified males (1.39 and 1.63 ± 0.41, respectively for the single non-sacculinized and sacculinized crabs) (Fig. 4). A general decreasing trend of K with size (CW) was also observed, as also identified as significant by the GLMM (Fig. 4, Table 2). In contrast, the %DW was unaffected by external infection (32.84 ± 4.58% and 32.09 ± 4.16%, respectively, in non-sacculinized and sacculinized crabs; P value = 0.099), though slightly lower in modified crabs (30.42 ± 5.44%), with no significant trend with size (Table 2).

Fig. 4. Mean Fulton's K and error bars (as standard deviation) for sacculinized (dark grey) and non-sacculinized (light grey) crabs per sex (F: females; M: males) and modified males (MM) together with number of crabs between brackets (upper panel); and Fulton's K in relation to size (CW, mm) (lower panel). Sacculinized crabs in dark grey circles; non-sacculinized crabs in dark grey squares.
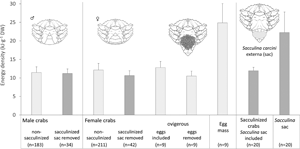
Contrasting with the morphological condition, the energy density of crabs was significantly affected by external infection (model results, Table 2), and size was only significant for E tot. The mean E DW was 4.3% lower in sacculinized crabs (excluding the externa sac), overall, and up to 5.8% lower in crabs <40 mm CW (though size was not a significant factor affecting E DW), with females more heavily affected (mean E DW of F 12.14 ± 1.77 and 10.65 ± 1.28, P value <0.001; M 11.46 ± 1.54 and 11.20 ± 1.22, P value = 0.293, respectively, for non-sacculinized and sacculinized females) (Fig. 5). The 2019 dataset provided extra information on the externa sac energy content (Fig. 5) allowing determining the mean E DW of the pair ‘sacculinized crab-parasite including the externa sac’, which was found to be very close to the mean E DW of non-sacculinized crabs (only 1.2% lower), even though the sac itself had about the double of the energy density – mean E DW of the sac 22.27 ± 5.52 vs 11.57 ± 1.49 kJ of non-sacculinized crabs (all 2019 crabs were <40 mm CW). The 2019 dataset further provided extra information on mean E DW of the egg mass (Fig. 5). Mean E DW of the egg mass (24.56 ± 5.21 kJ) was even higher than the E DW of externa sac resulting in a mean E DW of ‘ovigerous females including the egg mass’ (12.79 ± 1.67 kJ), 10.8% higher than mean E DW of (non-sacculinized) crabs of similar size.

Fig. 5. Mean E DW (kJ.g−1) and error bars (as standard deviation) for sacculinized and non-sacculinized crabs per sex (upper panel); and mean E DW (kJ.g−1) and error bars (as standard deviation) for sacculinized, for non-sacculinized crabs, for the externa sac, the pair crab host-externa, the egg mass, and for the pair ovigerous females-egg mass (lower panel). Sacculinized crabs in dark grey; non-sacculinized crabs in light grey. Number of crabs between brackets.
The E AF condition provided confounding results. Externally infected females reduced in 5.8% the E AF in relation to non-sacculinized ones (mean E AF 18.14 ± 1.66 and 17.08 ± 1.10, respectively, for in non-sacculinized and sacculinized females; P value <0.001), but infection did not affect males’ E AF (mean E AF 17.55 ± 1.27 and 17.78 ± 0.91, respectively, for in non-sacculinized and sacculinized males; P value = 0.224) (Fig. 6).

Fig. 6. Mean E AF (kJ.g AFDW−1) and error bars (as standard deviation) for sacculinized and non-sacculinized crabs per sex. Sacculinized crabs in dark grey; non-sacculinized crabs in light grey. Number of crabs between brackets.
In relation to E tot, the energy of crabs increased with size (Fig. 6) and, considering similar sized crabs, E tot was slightly lower for sacculinized crabs, externa sac excluded. The 2019 dataset, however, included data on E tot of the parasites’ externa sac (mean 2.93 ± 1.86 kJ, Fig. 7) and enabled to determine the total energy of the pair (sacculinized crab-parasite including externa). The same way, the E tot of the egg mass measured with the 2019 dataset (mean 12.30 ± 7.39 kJ), enabled to determine the total energy of the pair (ovigerous female-egg mass) (Fig. 7). Though mean E tot of the pair (sacculinized crab-parasite including externa) (19.49 ± 8.48 kJ) was much higher (about 30.8% higher) than the E tot of non-sacculinized crabs of similar size (14.90 ± 6.87 kJ), mean E tot of the pair [ovigerous females-egg mass] was even higher (48.5% higher; 41.93 ± 12.48 kJ and 28.23 ± 11.86 kJ, respectively for ovigerous and for non-sacculinized crabs of similar size).

Fig. 7. Total energy (E tot, kJ) in relation to size (CW, mm) for sacculinized and non-sacculinized crabs, for the externa sac, for the egg mass, and for the pairs [sacculinized crab-parasite including externa] and [ovigerous females-egg mass].
Discussion
The present study described patterns in the external infection prevalence of the rhizocephalan parasite Sacculina carcini in a shore crab population from the western Dutch Wadden Sea. The results show a slight (up to 5.8%), but significant decrease in the energy density of sacculinized crabs (externa excluded), although the morphometric body condition was not compromised by external infection. The reduction in energy density was more evident in crabs up to 40 mm size, and in females. However, when the parasite sac was considered, the difference in energy condition between sacculinized and non-sacculinized crabs was attenuated because, despite small-sized (and hence with low amount of energy in total), the sac had almost the double of the energy density of the non-sacculinized crabs. In addition, the estimation of the energy content of the crab egg mass showed that both the energy density and total energy of the egg mass was even higher than that of the externa sac, and consequently, the pair ovigerous female-egg mass had the highest energy condition.
Patterns of Sacculina carcini external infection prevalence
A relatively small fraction of the crab population from the western Dutch Wadden Sea (only 3.3%) was externally infected by S. carcini, which is in accordance with a contemporaneous study in the area, reporting up to 3% prevalence of Sacculina infection (Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016). Prevalence of sacculinized crabs has a great spatial variation (Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018). It is even lower (1.9–2.9%) in the Isfjord, Denmark (Lützen, Reference Lützen1984) and similarly low (2.9%) in the west coast of Sweden (Werner, Reference Werner2001). Yet, while in the Wadden Sea, scarred crabs and modified males with no sac nor scar were nearly absent (respectively 0.7% and 0.2% of the total population), in Sweden scarred crabs accounted for an extra 2.6%, and uninfected modified males for an extra 2.7% of the population (Werner, Reference Werner2001). Such a small fraction of scarred crabs did not allow evaluating the potential of recovery from infection. Elsewhere, the prevalence of S. carcini can range up to 20%, and can locally even reach 40–80% (Lützen, Reference Lützen1984; Mathieson et al., Reference Mathieson, Berry and Kennedy1998; Torchin et al., Reference Torchin, Lafferty and Kuris2001; Werner, Reference Werner2001; Zetlmeisl et al., Reference Zetlmeisl, Hermann, Petney, Glenner, Griffiths and Taraschewski2011). All these estimates (including present data) are conservative values, as the fraction of the immature phase of infection (interna, without externa) was not quantified. Quantification of internal infection requires the dissection of the crabs (Belgrad and Griffen, Reference Belgrad and Griffen2015; Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018), which is time consuming and out of the scope of the present studyi. Histological examination of shore crabs’ hepatopancreas resulted in a mean of 24% prevalence of infected shore crabs in a population from South Wales (UK), though only 6.3% crabs were externally infected and 1.8% had scars (Rowley et al., Reference Rowley, Davies, Malkin, Bryan, Thomas, Batista and Coates2020). Assuming a similar fraction, infected crabs in the Dutch Wadden Sea could be in the order of 12–13% of the population, internally infected crabs accounted.
A seasonal pattern in Sacculina prevalence has been described in previous studies. In general, prevalence peaks in winter and minimums are observed in summer (Costa et al., Reference Costa, Bessa and Pardal2013; Jensen et al., Reference Jensen, Schneider, Høeg, Glenner and Lützen2019), as described here for the Dutch Wadden Sea population. Therefore, increased mortality of externa has been related to the temperature rise during summer (Heath, Reference Heath1971). Besides temperature, also salinity conditions influence the prevalence of infected crabs. The lower salinity limit for maintaining a permanent population of Sacculina seems to be around 20 (Kashenko and Korn, Reference Kashenko and Korn2002; Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018), which fits the range (18–29) observed in the present (Campos et al., Reference Campos, Ribas, Bio, Freitas, Souza and van der Veer2021) and in the contemporaneous work in the area (Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016). Yet, as in other sacculinid rhizocephalan species (Walker et al., Reference Walker, Clare, Rittschof and Mensching1992), S. carcini eggs and larvae are more sensitive (Tolley et al., Reference Tolley, Winstead, Haynes and Volety2006), requiring salinity >26 (Ramult, Reference Ramult1935), and condition the distribution of infected crabs in relation to salinity (Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018).
The prevalence of sacculinized crabs also varies with water depth (Rainbow et al., Reference Rainbow, Ford and Heppelwhite1979; Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016). The deeper subtidal waters house older crabs with the highest levels of S. carcini (Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016), while near none is found in the intertidal flats (Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016). Water depth itself may not be a forcing reason but the depth distribution of crabs of different sizes, as older and larger crabs are typically more abundant in the subtidal (Atkinson and Parsons, Reference Atkinson and Parsons1973; Hunter and Naylor, Reference Hunter and Naylor1993). Yet, preference for host size has not been documented (Werner, Reference Werner2001; Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016; Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018). In the present study, despite sampling took place always at the subtidal (5–12 m deep), Sacculina externally infected crabs mainly between 25 and 40 mm width, while only 10% of the largest crabs (>55 mm) were infected. This was also the size range with higher prevalence of infection previously reported for the area (Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016) and for the Mondego estuary, in Portugal (Costa et al., Reference Costa, Bessa and Pardal2013). In the present work, the largest sacculinized crab was a male 88.6 mm CW and the smallest was a modified male 14.8 mm. This is smaller than previously reported (20 mm, Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018) and contradicts the assumption that crabs 3–16 mm do not become infected, as revealed by molecular analyses (Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018). When feminized males were accounted, prevalence of male external infection doubled the prevalence of sacculinized female crabs. Yet, preference for host gender has not been reported (Werner, Reference Werner2001; Costa et al., Reference Costa, Bessa and Pardal2013; Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016), with some populations revealing more sacculinized males (e.g. Isefjord, Denmark, Lützen, Reference Lützen1984) and others more females (e.g. Mondego estuary, Portugal, Costa et al., Reference Costa, Bessa and Pardal2013; Danish coast, Mouritsen et al., Reference Mouritsen, Geyti, Lützen, Høeg and Glenner2018; Scotland, Mathieson et al., Reference Mathieson, Berry and Kennedy1998).
The developmental stages of S. carcini externa seem correlated with the crab host colour cycle such that the youngest externa stages are predominant when the crabs are green, the intermediate stages when they are orange and the oldest when they are red (Lützen et al., Reference Lützen, Jensen and Glenner2018). In the present study, the externa development stages were not evaluated, but external infection was more often found in green morphs, suggesting then that most parasites’ externa would be on an early stage. Literature, however, refers that red crabs are more commonly infected than green ones (Zetlmeisl et al., Reference Zetlmeisl, Hermann, Petney, Glenner, Griffiths and Taraschewski2011; Costa et al., Reference Costa, Bessa and Pardal2013; Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016), as infected animals remain longer in the intermoult stage which is prolonged in the red morphotype (Høeg, Reference Høeg1995).
Impact of external infection on crabs’ condition
Infection by S. carcini causes castration, moulting arrest, behavioural and even body shape changes (Thresher et al., Reference Thresher, Werner, Høeg, Svane, Glenner, Murphy and Wittwer2000; Mouritsen and Jensen, Reference Mouritsen and Jensen2006; Kristensen et al., Reference Kristensen, Nielsen, Jørgensen, Mouritsen, Glenner, Christensen, Lützen and Høeg2012; Belgrad and Griffen, Reference Belgrad and Griffen2015; Coates and Söderhäll, Reference Coates and Söderhäll2021), including the broadening of the abdomen in male crabs (Reinhard, Reference Reinhard1956), a feminized attribute. All these effects stress the animals, suppress their defences, including a reduction in burrowing capacity, interrupt moulting and may favour increased colonization by epibionts (Mouritsen and Jensen, Reference Mouritsen and Jensen2006). This would imply that infected crabs face extra energy costs due to the presence of the parasites, resulting in a lower general condition and a reduction in energy content. While the present findings revealed in fact a slight decrease in energy density in sacculinized crabs (externa excluded, and slightly less when sac was included), condition was evaluated for the pair host–parasite, because Sacculina internal roots cannot be dissected without losing relevant biological material. The main body of the parasite is a root-like system of branches that extends into most of the host's tissue, whereas the gonads of the parasites emerge externally under the abdomen of the crab (Høeg and Lützen, Reference Høeg and Lützen1995). Therefore, the internal part of the parasite body was impossible to remove prior to calorimetric analysis resulting in a biased, i.e. apparently smaller, reduction in the host's energetic condition: part of the measured organic weight and energetic content corresponded in fact to the parasite roots. This is probably the reason why the morphological condition of infected crabs was not affected by infection, as the extra weight of the parasite itself – corresponding to the internal roots – was included. It also means that the reduction in energy density of infected crabs accounts not only for the energy density of the host but also for the energy content of the parasite itself. And again this could explain why sacculinized crabs plus externa sac had higher total energy (about 30%) than non-sacculinized crabs of similar size, as an undetermined part of the energy belonged to the parasite. In addition, the extent of S. carcini rootlets within the crab internal tissues was not evaluated and might not be equivalent between sacculinized crabs (Powell and Rowley, Reference Powell and Rowley2008).
The arrest of moult and reproduction in sacculinized crabs, releases the energy pool which would be allocated for growth and gonad maturation of the host crab. In laboratory feeding trials with mussels, infection by Sacculina had no effect on food consumption by male crabs (Larsen et al., Reference Larsen, Hoeg and Mouritsen2013), which means that the energy released from crabs’ growth is available for the parasite to spend on its own growth and reproductive requirements. Therefore, expectations of similar nutritional condition between non-sacculinized and sacculinized crabs including the parasite were partly confirmed in the present findings, as only a very slight decrease in energy density (1.2%) was found when the energy of the parasite sac was accounted for. However, the combined energy of host and parasite including externa in total was higher, suggesting high efficiency of the parasite allocating host energy. Yet, the energy content of the host, if isolated from the parasite, must be further reduced to an unknown extent in comparison to uninfected crabs.
Besides the impact on crabs’ growth, Sacculina tend to feminize male hosts both morphological and behaviourally (Høeg, Reference Høeg1995; Mouritsen and Jensen, Reference Mouritsen and Jensen2006; Lafferty and Kuris, Reference Lafferty and Kuris2009; Kristensen et al., Reference Kristensen, Nielsen, Jørgensen, Mouritsen, Glenner, Christensen, Lützen and Høeg2012). In shore crabs, sexual dimorphism is largely confined to a relatively broader abdomen and smaller master chelae in females (Shen, Reference Shen1935; Elner, Reference Elner1980). Several other female attributes can also be impacted by S. carcini infection. The externa sac is lodged in the same location of the crab's body where females carry the egg mass – no ovigerous female has ever been found sacculinized. Infected crabs (male or female) then develop a nurturing behaviour typical of an ovigerous female crab towards the parasite externa: the crab cares for the sac as if it was their own eggs (Høeg and Lützen, Reference Høeg and Lützen1995; Thresher et al., Reference Thresher, Werner, Høeg, Svane, Glenner, Murphy and Wittwer2000). Further, the parasite takes advantage of the natural hatching process of the crab to release its larvae into the water when the externa is matured (Øksnebjerg, Reference Øksnebjerg2000). Infected crabs also tend to migrate to more saline and deeper areas (Rasmussen, Reference Rasmussen1959; Rainbow et al., Reference Rainbow, Ford and Heppelwhite1979; Lützen, Reference Lützen1984; Waser et al., Reference Waser, Goedknegt, Dekker, McSweeney, Witte, van der Meer and Thieltges2016), which favour Sacculina development (Kashenko and Korn, Reference Kashenko and Korn2002), a migration pattern typical of ovigerous females (Baeta et al., Reference Baeta, Cabral, Neto, Marques and Pardal2005).
As described above, Sacculina seems to use efficiently the energy left from the crabs’ growth and maintenance. However, the comparison with the energy female crabs invest in reproduction is much more complicated. In fact, the energy density of the egg mass is much higher than the energy density of sacculinized crabs, even when the energy of the sac is accounted for, resulting in 4 times more energy in total of the egg mass in relation to the externa sac, though this may vary with externa stage of development and available data are restricted to a single month. Yet, while female crabs produce a single brood per year (Younge and Elliott, Reference Young and Elliott2020) and hence the investment in reproduction is well represented in the energy density of the egg mass, S. carcini can produce several successive broods (Thresher et al., Reference Thresher, Werner, Høeg, Svane, Glenner, Murphy and Wittwer2000), and thus the snapshot information of the sac energy density is not enough to inform on the parasites’ energy investment in reproduction, taken from the host.
Besides arresting the host growth and reproduction, and doubling the chances of mortality of crabs (Goddard et al., Reference Goddard, Torchin, Kuris and Lafferty2005), the present findings describe the effects of S. carcini external infection on crabs’ energetic condition, although conservatively, requiring that in future studies the extent of internal infection is disentangled. As for the implications of such energy reduction due to parasitism for the C. maenas population dynamics in the Dutch Wadden Sea further investigations are required.
Acknowledgements
The authors acknowledge Sander Holthuijsen from the Royal Netherlands Institute for Sea Research (NIOZ) for his valuable assistance with bomb calorimetry. We thank the 2 anonymous reviewers whose comments and suggestions helped improve and clarify this manuscript.
Author contributions
H. W. V., J. C. and A. T. S. conceived and designed the study; F. R. conducted the data gathering; A. B. performed statistical analyses and graphical work; J. C. and V. F. wrote the article; H. W. V., A. T. S. and F. R. reviewed and edited the manuscript.
Financial support
This research was supported by national funds by FCT – Foundation for Science and Technology – within the scope of UIDB/04423/2020 and UIDP/04423/2020, results from the project ATLANTIDA (ref. NORTE-01-0145-FEDER-000040), and was funded by the project Ocean3R (NORTE-01-0145-FEDER-000064), supported by the North Portugal Regional Operational Programme (NORTE2020), under the PORTUGAL 2020 Partnership Agreement and through the European Regional Development Fund (ERDF).
Conflict of interest
The authors declare there are no conflicts of interest.
Ethical standards
Not applicable.









