



Introduction
Cutaneous leishmaniasis (CL) comprises a broad spectrum of clinical manifestations caused by over 20 different kinetoplastid Leishmania species transmitted by female sandflies (WHO, 2020). In Asia and Africa, most CL cases are caused by Leishmania major and Leishmania tropica, leading to self-limiting ulcers. In the Americas, the disease is caused by several species, including Leishmania braziliensis, Leishmania mexicana, Leishmania guyanensis, Leishmania naiffi and Leishmania amazonensis. The clinical presentations range from self-healing localized skin lesions to disfiguring mucocutaneous ulcerations (Martins et al., Reference Martins, Barreto, Lauris and Martins2014; de Vries et al., Reference de Vries, Reedijk and Schallig2015). Annually, about 1.2 million new CL cases are reported, and a significant number of patients develop severely disfiguring permanent scars, which results in social stigma and loss of economic productivity (Alvar et al., Reference Alvar, Vélez, Bern, Herrero, Desjeux, Cano, Jannin and de Boer2012; Baileyid et al., Reference Baileyid, Mondragon-Shem, Haines, Olabi, Alorfi, Ruiz-Postigo, Alvar, Hotez, Adams, Vélez, Al-Salem, Eaton, Acosta-Serrano and Molyneux2019). CL is one of the most mistreated illnesses among neglected tropical diseases (NTDs). The large variation of drug susceptibility among the diverse parasite species makes the drug discovery process even more difficult (de Vries et al., Reference de Vries, Reedijk and Schallig2015; Van Bocxlaer et al., Reference Van Bocxlaer, Caridha, Black, Vesely, Leed, Sciotti, Wijnant, Yardley, Braillard, Mowbray, Ioset and Croft2019).
The first-line treatment for CL includes pentavalent antimonials: sodium stibogluconate and meglumine antimoniate. Antimonials present several drawbacks in terms of efficacy, safety (cardiotoxicity and hepatotoxicity) and protracted periods of drug administration (Weisz, Reference Weisz2006). Alternative systemic agents include miltefosine, amphotericin B, pentamidine isethionate, paromomycin and granulocyte-macrophage colony-stimulating factor, limited by severe adverse effects (de Vries et al., Reference de Vries, Reedijk and Schallig2015). There is a need for safer, more selective and easily accessible therapies for CL.
A few new small molecules are currently in preclinical and clinical development against Leishmania. One example is the first-in-class kinetoplastid-selective proteasome inhibitor compound LXE408. LXE408 was discovered by Novartis with financial support from the Wellcome Trust. LXE408 is efficacious in the murine model of CL, and it is under clinical trial for the treatment of visceral leishmaniasis (Nagle et al., Reference Nagle, Biggart, Be, Srinivas, Hein, Caridha, Sciotti, Pybus, Kreishman-Deitrick, Bursulaya, Lai, Gao, Liang, Mathison, Liu, Yeh, Smith, Lerario, Xie, Chianelli, Gibney, Berman, Chen, Jiricek, Davis, Liu, Ballard, Khare, Eggimann, Luneau, Groessl, Shapiro, Richmond, Johnson, Rudewicz, Rao, Thompson, Tuntland, Spraggon, Glynne, Supek, Wiesmann and Molteni2020). Partnerships between not-for-profit research and private pharmaceutical companies for the research and development of new treatments for NTDs such as leishmaniasis are increasing (Katsuno et al., Reference Katsuno, Burrows, Duncan, van Huijsduijnen, Kaneko, Kita, Mowbray, Schmatz, Warner and Slingsby2015).
Some of the compounds in development include nitroaromatic compounds, which are historically used to treat several diseases caused by protozoans, for example, DNDI-8219, DNI-VL2098 and DNDI-0690 (Van Bocxlaer et al., Reference Van Bocxlaer, Caridha, Black, Vesely, Leed, Sciotti, Wijnant, Yardley, Braillard, Mowbray, Ioset and Croft2019). In this study, we envision that scaffold-driven and easily synthesizable nitroaromatic compounds based on prior art can facilitate rapid phenotypic screenings using in vitro and in vivo experimental models. The scaffolds were selected based on our ongoing research on the use of natural product-derived motifs in the design and development of antiprotozoal agents (Zhang et al., Reference Zhang, Collins, Nyamwihura, Ware, Kaiser and Ogungbe2018). The compounds were evaluated against the free amastigote forms (AFs) of L. amazonensis (strain LTB0016), purified from paw lesion, followed by evaluation against the intracellular forms as well as on peritoneal macrophages (PMMs). The most selective were evaluated in L. amazonensis-infected BALB/c mice. The methods and results are presented below.
Materials and methods
Compounds: Dimethyl sulphoxide (DMSO, Merck) solutions (30 mm) of the compounds (Fig. 1) and miltefosine ⩾98% (Sigma-Aldrich, St. Louis, USA) were diluted just before the in vitro experiments into RPMI-1640 medium (pH: 7.2–7.4) without phenol red (Sigma-Aldrich (St. Louis, USA), R7509) but supplemented with 1% l-alanyl-l-glutamine (GLUTAMAX I, Gilbco™), 1% penicillin–streptomycin 5000 U mL−1 (Gibco™, PEN-STR) and 10% heat-inactivated sterile-filtered fetal bovine serum (FBS), from Cultilab, Brazil. The concentration of DMSO was <0.6% for all in vitro experiments to prevent non-specific toxicity to host cells (Romanha et al., Reference Romanha, de Castro, Soeiro, Lannes-Vieira, Ribeiro, Talvani, Bourdin, Blum, Olivieri, Zani, Spadafora, Chiari, Chatelain, Chaves, Calzada, Bustamante, Freitas-Junior, Romero, Bahia, Lotrowska, Soares, Andrade, Armstrong, Degrave and Andrade2010). For in vivo assays, compounds 6 and 7 were prepared daily in a solution composed of 5% Gum Arabic from acacia tree (Sigma-Aldrich, St. Louis, USA), 6% DMSO and 3% Tween™ 80 Surfact-Amps™ detergent solution (Thermo Scientific™). The reference drug, Milteforan™ (Mt™), was dissolved in sterile deionized water.

Fig. 1. Structures of the tested compounds.
Parasite strain and mammalian host cell cultures
Leishmania amazonensis (strain LTB0016) was used throughout the study. AFs were purified from male BALB/c mice. Briefly, the foot paws were inoculated (subcutaneously) with 20 μL containing 106 amastigotes, and the animal skin lesions were aseptically removed 30 days post-infection (dpi). The parasites in the lesions were mechanically dispersed by pipetting and used for the assays. The compounds were tested on (1) extracellular AFs; (2) intracellular amastigote forms (IA) in PMMs and (3) BALB/c mouse models infected with amastigotes as recently described (Cardoso Santos et al., Reference Cardoso Santos, Meuser Batista, Inam Ullah, Rama Krishna Reddy and Soeiro2021). PMMs were obtained from Swiss male mice (18–20 g) inoculated with 3% Brewer thioglycollate medium (Merck) previously diluted in water and autoclaved. After 4 days of stimulation, the cells were collected by rinsing the animals' peritoneum with RPMI 1640. The cells were subsequently seeded in 24- (3 × 105 cells well−1) and 96-well (5 × 104 cells well−1) plates for analysis of infection and cytotoxicity assays, respectively. Cell cultures were maintained at 37°C, 5% CO2 atmosphere in RPMI 1640 medium (pH: 7.2–7.4) without phenol red, but supplemented with 1% l-glutamine, 1% (PEN-STR) and 10% FBS. Assays using AF were maintained at 32°C using RPMI culture medium containing 5% FBS.
Cytotoxicity and in vitro leishmanicidal analysis
The toxicity of the compounds on host cells was evaluated 24 h after PMMs were seeded into culture plates. The compounds (0–500 μ m) were added to the wells and incubated as described above for 48 h, and cellular viability was evaluated using the AlamarBlue test (Invitrogen) according to the manufacturer's recommendations. The effect of the compounds (0–20 μ m) on AFs was evaluated using 106 parasites per well (0.2 mL) for 48 h, and parasite viability assessed with AlamarBlue tests as reported (Mikus and Steverding, Reference Mikus and Steverding2000; Santos et al., Reference Santos, Zhang, Batista, de Oliveira, Demarque, da Silva-Gomes, Moreira, Ogungbe and Soeiro2020). Phenotypic screenings against IAs were performed by infecting 3 × 105 PMMs with 9 × 105 amastigotes, multiplicity of infection being 3. After 48 h of drug treatment, cultures were rinsed with saline, fixed 5 min with Bouin and stained for 15 min with Giemsa solution (Sigma-Aldrich (St. Louis, USA), 32884), filtered and diluted five times in distilled water (Santos et al., Reference Santos, Lionel, Peres, Batista, da Silva, de Oliveira, da Silva, Batista, Souza, Andrade, Neves, Braga, Patrick, Bakunova, Tidwell and Soeiro2018; Feitosa et al., Reference Feitosa, da Silva, Hoelz, Souza, Come, Cardoso-Santos, Batista, Soeiro, Boechat and Pinheiro2019). Samples were subsequently evaluated by light microscopy and photographs were obtained by using a Zeiss AxioObserver MI microscope (Oberkochen, Germany). The percentage of infected host cells, the number of parasites per infected cell and the corresponding infection index were obtained. Parasites with well-defined nuclei and kinetoplasts were counted as surviving since irregular structures could mean parasites undergoing death. The results were expressed as % reduction in parasite burden, and the half-maximal effective concentration (EC50) and 90% maximal effective concentration (EC90) were calculated by non-linear regression analysis using GraphPad Prism v.9.1.2. All assays were run at least twice with three independent replicates each time.
Hep G2 viability assay
Hep G2 (CRL-11997™) cells were grown in complete growth medium (Dulbecco's modified Eagle medium: F12 containing l-glutamine and sodium bicarbonate, 10% FBS and 1% penicillin/streptomycin) incubated at 37°C in a 5% CO2 environment. Cells were seeded into 96-well plates at 5 × 105 cells mL−1 and incubated for about 24 h. Cells were treated with the compounds prepared in DMSO for 72 h at a final concentration range of 160–1.2 μ m in triplicates. Cell viability was assessed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide assay as previously described (Zhang et al., Reference Zhang, Collins, Nyamwihura, Ware, Kaiser and Ogungbe2018). Podophyllotoxin was used as cytotoxicity control, and sodium dodecyl sulphate (10%) was used as assay control.
In vivo efficacy analysis
Male BALB/c mice (18–20 g) obtained from the animal facilities of ICTB (Institute of Science and Biomodels Technology, Fiocruz, RJ, Brazil) were housed five per cage, kept in a room at 20–24°C under a 12-light and 12-h dark cycle and provided with sterile water and chow ad libitum. Animals were subcutaneously inoculated with 5 × 105 amastigotes on the left foot paw (Godinho et al., Reference Godinho, Simas-Rodrigues, Silva, Ürmenyi, De Souza and Rodrigues2012). The treatment started 15 dpi, corresponding to the lesion onset (size diameter of 3–4 mm) (Van Bocxlaer et al., Reference Van Bocxlaer, Caridha, Black, Vesely, Leed, Sciotti, Wijnant, Yardley, Braillard, Mowbray, Ioset and Croft2019). The compounds were given for 14 days by oral gavage (100 μL) at 10 mg kg−1, twice a day (q12h). Mt™ was administered at a standardized dose (40 mg kg−1) once a day (q24h). The lesions were measured regularly in three dimensions (height, width and depth), and animals were euthanized at 31 dpi. Skin lesions aseptically removed were used for molecular analysis of parasite load by quantitative polymerase chain reaction (qPCR) and light microscopy inspections of imprints (Ribeiro-Romão et al., Reference Ribeiro-Romão, Saavedra, Da-Cruz, Pinto and Moreira2016). All procedures were carried out in accordance with the guidelines established by the FIOCRUZ Committee of Ethics for the Use of Animals (CEUA L038/2017). Statistical analysis was conducted in GraphPad Prism v.9.1.2 by analysis of variance (ANOVA): (1) one-way Fisher's LSD test or (2) two-way unpaired t-test with Welch's correction, significance P < 0.05 [95% confidence interval (CI)].
Results
All experimental details are fully described in the Supplementary material. The compounds (Fig. 1) were synthesized as shown in Fig. S1 and diluted just before the in vitro experiments. First, the compounds were screened at 10 μ m against extracellular amastigotes of L. amazonensis purified from male BALB/c paw lesions (Feitosa et al., Reference Feitosa, da Silva, Hoelz, Souza, Come, Cardoso-Santos, Batista, Soeiro, Boechat and Pinheiro2019) using colorimetric assays (AlamarBlue test, Invitrogen) (Mikus and Steverding, Reference Mikus and Steverding2000; Godinho et al., Reference Godinho, Simas-Rodrigues, Silva, Ürmenyi, De Souza and Rodrigues2012). Two (6 and 7) out of the seven compounds caused a significant decline in the total number of live parasites (⩾50% decrease compared to untreated parasites) with only 48 h of drug exposition. Next, 6 and 7 were further analysed in dose–response assays. The EC50 values obtained were 10.78 ± 0.80 and 13.12 ± 1.70 μ m, respectively (Table 1). Host cell viability showed that compounds did not exhibit any toxicity up to the highest tested concentration, leading to CC50 PMM values >500 μ m. Also, when cellular viability was assessed using Hep G2 cultures, no toxicity was observed (CC50 Hep values >160 μ m, Table 1). The selective antiparasitic activity was encouraging, and we decided to evaluate the compounds on intracellular amastigotes (IA) in PMMs (9 × 105 IA: 3 × 105 PMMs well−1), which mimics typical infection in mammalian cells. Light microscopy analysis showed that 6 was about 3-fold more active compared to the reference drug (miltefosine, P = 0.0273). Its EC50 was 4.57 μ m, and it is quite selective [selective index (SI) > 100]. Compound 6 also displayed low EC90 (9.14 μ m) and caused a 92% decrease in PMM infection at 10 μ m. Compound 7 is slightly less potent (EC50 = 9.19 μ m and EC90 = 17.2 μ m) with an SI of >50 (Table 1). Both compounds displayed an impressive leishmanicidal effect due to a drastic decrease in the number of parasites per cell and the ratio of infected cells at 20 μ m (Fig. S2). These favourable results prompted us to conduct preliminary in vivo proof of concepts using both hit compounds.
Table 1. Phenotypic studies of compounds 6 and 7 against Leishmania amazonensis and their toxicity profile

AF, amastigote forms of L. amazonensis (LTB0016 strain) purified from animal lesions; IA, intracellular forms in peritoneal macrophages; SI, selective index.
EC50 is reported as mean and s.d. values.
a Statistical analysis was performed with GraphPad prism v.9.1.2 by ordinary ANOVA test.
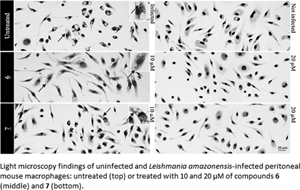
In the in vivo experiment, we observed a gradual increase in skin lesion size of mice treated with the vehicle alone, attaining 438.8 ± 42.05 mm3 at the endpoint (Fig. 2). Mice treated with compound 6 displayed similar lesions size (443.7 ± 47.76 mm3, P = 0.4798), whereas a slight increase was observed using 7 (550.9 ± 30.23 mm3, P = 0.2398), as compared to the vehicle-treated group. Mt™ achieved a 72% decrease in the size of skin lesions and significantly improved the clinical condition (P < 0.0001 (Fig. 2A and B). The qPCR standardization curves achieved all the desired efficiency and linearity coefficient criteria, ranging between 93.2–91.4% and 0.98–0.99% for the parasite kDNA amplification (Fig. S3A) and mice GAPDH target (Fig. S3B), respectively. At 31 dpi, qPCR readout expressed as DNA equivalents per mg of tissue (14) showed a 5–10-fold increase in the parasite load of mice treated with 6 (276 526 ± 40 519 eq. par mg−1 tissue, P = 0.0208) and 7 (153 765 ± 54 932 eq. par mg−1 tissue, P = 0.0829) as compared to the vehicle-treated group (27 753 ± 9539 eq. par mg−1 tissue) (Fig. 2B). Mt™ completely suppressed (99.99%) parasite load (0.256 ± 0.1022 eq. par mg−1 tissue, P = 0.0437). The qPCR showed that the hit compounds lacked in vivo efficacy, and it was corroborated by a qualitative light microscopy analysis of lesions imprints (Fig. 2C). The findings clearly showed that both 6- and 7-treated animal groups have high parasitaemia coupled with the presence of lymphomononuclear diffuse infiltrates and several parasite niches inside mononuclear host cells (Fig. 2).

Fig. 2. Effect of 6 and 7 in CL experimental mouse models using BALB/c mice infected with Leishmania amazonensis. The graphics show: average lesion size during treatment (A), the correlation between parasite load by qPCR (black bars) and average lesion size measurements (grey bars) at 31 dpi (B), according to each experimental group. Light microscopy of lesion imprints of infected mice treated with vehicle and after administration of compounds 6 and 7 at 10 mg kg−1 q12h po (C). Arrow: intracellular parasites. Statistical analysis was performed with GraphPad prism v.9.1.2 by ANOVA test (95% CI). When P value ⩾0.05 = not significant (ns).
The lack of in vivo efficacy suggests low bioavailability of the compounds in mice. Low bioavailability is generally caused by impaired absorption or high clearance of compounds. Therefore, an in silico tool, pkCSM, was used to predict some critical absorption, distribution and metabolism parameters of 6 and 7, comparing to miltefosine (Gleeson, Reference Gleeson2008; Pires et al., Reference Pires, Blundell and Ascher2015). Compounds 6 and 7 do not violate Lipinski's rule. The cLog Ps of 6 and 7 are 2.97 and 2.13, respectively, while that of miltefosine is 5.68 (Table 2). Since miltefosine is zwitterionic, its relatively high cLog P is ideal for high intestinal absorption. Similarly, since compounds 6 and 7 have low molecular weights (324.34 and 297.27 g mol−1, respectively) and are neutral molecules at physiological pH with relatively low cLog P, sufficient intestinal adsorption was expected from oral gavage. The predicted high intestinal absorption further supports this in addition to the predicted moderate to high Caco-2 permeability (0.83 and 1.04, respectively) from in silico Absorption, Distribution, Metabolism, and Excretion (ADME) calculations. Oral gavage was selected for this study because oral administration is the preferred route for the clinical use of new antileishmanial drugs. The predicted volume of distribution (VDss) is high and ideal for tissue distribution for both compounds. However, the predicted unbound fraction for 6 is relatively low (5%) and ideal for 7 (21%). Both compounds are predicted as CYP3A4 substrates, which suggest significant first-pass hepatic clearance and low overall plasma and tissue concentration. Therefore, a comparative pharmacokinetics study (oral and intravenous) can provide insight into plasma concentrations and bioavailability of both compounds in addition to determining the appropriate dosage and structural changes necessary for in vivo efficacy. Also, in silico toxicological analysis of both compounds suggests probable mutagenicity (positive for Ames tests) and potential to inhibit hERG II and that 6 is potentially hepatotoxic. However, animals treated with compounds 6 and 7 did not show any observable side-effects. A cell proliferation assay using Hep G2 cells indicates that the compounds are not hepatotoxic at the tested concentrations (160–1.2 μ m).
Table 2. Predicted ADME properties of 6, 7 and miltefosine

a The predicted values are for humans.
Discussion
Leishmania amazonensis is one of the most common Leishmania species in Brazil, known to cause a wide spectrum of pathologies, including highly severe and diffuse CL (Lainson et al., Reference Lainson, Shaw, Silveira, de Souza, Braga and Ishikawa1994). Most CL patients live in impoverished communities with limited access to primary healthcare facilities (Okwor and Uzonna, Reference Okwor and Uzonna2016). The lack of access to primary healthcare typically means that the disease is usually at an advanced stage before medical intervention is sought, if available (Ruoti et al., Reference Ruoti, Oddone, Lampert, Orué, Miles, Alexander, Rehman, Njord, Shu, Brice, Sinclair and Krentel2013). In this study, we explored the in vitro and in vivo leishmanicidal effects of compounds derived from two nitroaromatics scaffolds (4-nitrophenylacetyl and 4-nitro-1H-imidazolyl). The in vitro data demonstrated that two 4-nitro-1H-imidazolyl compounds have acceptable selectivity indices (>50 for intracellular forms of L. amazonensis) as previously discussed by Caridha et al. (Reference Caridha, Vesely, Van Bocxlaer, Arana, Mowbray, Rafati, Uliana, Reguera, Kreishman-Deitrick, Sciotti and Buffet2019) as well as by Alcântara et al. (Reference Alcântara, Ferreira, Gadelha and Miguel2018). Therefore, their antileishmanial activities were investigated further. The compounds were administered to BALB/c mouse models of CL for 14 days at a relatively low dose, starting at the onset of lesions (Godinho et al., Reference Godinho, Simas-Rodrigues, Silva, Ürmenyi, De Souza and Rodrigues2012). The results showed that the in vitro potency of the hit compounds 6 and 7 did not translate to desired in vivo efficacy. The measurements of the mouse lesions (using a paquimeter) and the molecular analysis of animal parasitism (qPCR) demonstrated a lack of in vivo activity of both compounds, whereas Mt™ suppressed both parameters, as reported (Van Bocxlaer et al., Reference Van Bocxlaer, Caridha, Black, Vesely, Leed, Sciotti, Wijnant, Yardley, Braillard, Mowbray, Ioset and Croft2019). The lack of in vivo efficacy of the tested compounds could be, at least in part, due to low bioavailability and/or high metabolic clearance. Nevertheless, as nitro-drugs are activated by Leishmania nitroreductases (NTR2) (Wyllie et al., Reference Wyllie, Roberts, Norval, Patterson, Foth, Berriman, Read and Fairlamb2016), it is possible that 6 and 7 may be also activated in a similar way, and thus, the in vitro leishmanicidal efficacy supports their use as templates for nitroimidazole-based antileishmanial drug discovery programmes focusing on analogues that have the appropriate Target Product Profile (TPP) for new CL drugs, as recommended (Drugs for Neglected Diseases Initiative, 2020).
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182021002079
Acknowledgements
The authors acknowledge the Fortalecimento dos Programas de Gestão Estratégica de Pesquisa da Fiocruz Rede de Plataformas Fiocruz (VPPLR-001-Fio 14) and Programa de Excelência Acadêmica (PROEX) from CAPES. They appreciate the excellent technical contribution of Roberson Donola Girão, Dr Ana Lia Mazzetti and Dr Cristiane França da Silva (Laboratory of Cellular Biology/IOC/Fiocruz) and Kingsley Bimpeh (Jackson State University).
Financial support
The current study was supported by grants from Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho Nacional Desenvolvimento científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Fundação Oswaldo Cruz PAEF/CNPq/Fiocruz. MNCS and OCM are research fellows of CNPq and CNE and JCNE researchers. The work at Jackson State University was supported, in part, by the US National Institutes of Health (SC3GM122629 and G12MD007581).
Conflict of interest
None.
Ethical standards
All procedures were carried out in accordance with the guidelines established by the FIOCRUZ Committee of Ethics for the Use of Animals (CEUA L038/2017).






