INTRODUCTION
The majority of nematodes are free-living, playing important roles in ecosystem health and diversity, but many are important and prolific parasites. For an idea of scale, approximately half the world's population, predominantly in tropical regions, harbour a nematode infection (www.nematode.net). These worms typically cause chronic infections that contribute significantly to morbidity, but rarely kill their hosts. The most abundant parasitic species are gastrointestinal nematodes such as Ascaris lumbricoides, which infects an estimated 807 million, closely followed by Trichuris trichiura (infecting an estimated 604 million) and the hookworms (infecting approximately 576 million) (http://www.cdc.gov/parasites/). These infections are often particularly prevalent and clinically important in children. Filarial nematodes are tissue-dwelling worms that account for an additional 157 million infections and are the causative agents of river blindness (Onchocerca volvulus) and elephantiasis (Wuchereria bancrofti and Brugia spp.). In addition to their importance as human pathogens, around 10% of global crop loss is due to plant parasitic nematodes (Nicol et al. Reference Nicol, Turner, Coyne, Den Nijs, Hockland, Maafi, Jones, Gheysen and Fenoll2011), while nematode parasites of livestock cause major economic production losses to grazing ruminants. Currently, infection is controlled by the use of anthelmintic drugs. However, anthelmintic resistance, particularly in nematodes of small ruminants such as sheep and goats, is endemic in most countries (Gilleard, Reference Gilleard2006). Given that the same anthelmintic drugs are used in mass administration campaigns to treat human nematode infection, there is concern over the possible development of resistance in human nematodes (Vercruysse et al. Reference Vercruysse, Levecke and Prichard2012), underscoring the need for the development of new drugs to control these pathogens (Geary et al. Reference Geary, Woo, McCarthy, Mackenzie, Horton, Prichard, De Silva, Olliaro, Lazdins-Helds, Engels and Bundy2010). In this review, we focus on the role of Hsp90 in both free-living and parasitic nematodes, specifically Caenorhabditis elegans and Brugia spp. (life cycles of which are outlined in Fig. 1) and discuss the potential of Hsp90 as a drug target for the treatment of some helminth infections.

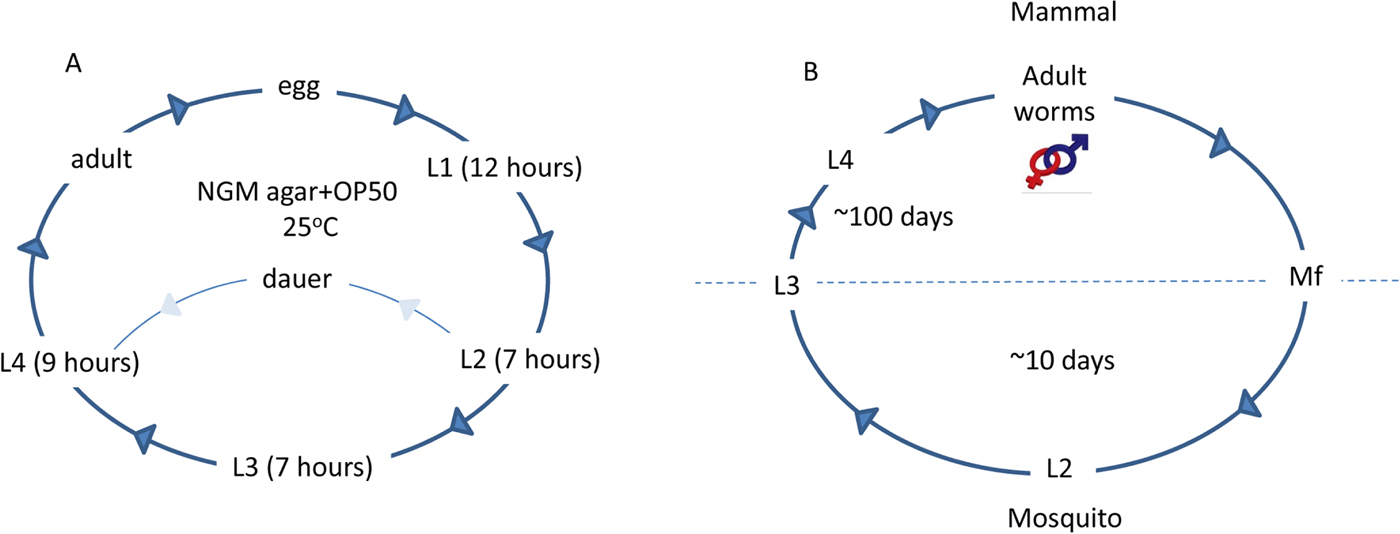
Fig. 1. Life cycle of the free-living nematode C. elegans and the parasitic lymphatic filarial nematodes. The normal hermaphroditic life cycle of C. elegans (A) takes approximately 3 days. Eggs are fertilized within the adult and after laying, hatch and worms proceed through four larval stages, each of which ends in a moult. Each adult will lay approximately 300 eggs and live for approximately 2 weeks. Under adverse conditions (i.e. lack of food, unfavourable temperatures or overcrowding), C. elegans can enter an alternative pathway called ‘dauer’. In this state, worms can remain dormant for 3 months and can re-enter the life cycle when the environment becomes favourable. All the lymphatic filarial nematodes share an invariant life cycle (B). The full developmental cycle takes place in the mosquito (the intermediate host) or the human (the definitive host). Infection of the definitive host is initiated by the bite of a mosquito harbouring the infective L3. The L3 enters the body via the puncture site and migrates to the lymphatic system of the mammalian host. L3 moult through L4 to become sexually mature adult male and female worms, which have a lifespan of approximately 8 years. After sexual reproduction the female worms release microfilariae (Mf, first stage larvae), which migrate to the bloodstream and are available for ingestion by a mosquito taking a blood meal. Development from Mf to the L3 stage within the mosquito occurs in the thoracic muscles and is a temperature-dependent process (optimal 28 °C and 80% humidity). Mature L3 migrate to the feeding structures in the head of the mosquito, which facilitates their transmission to the definitive host. It should be noted that other species of parasitic nematode do not require an arthropod intermediate host and have ‘free-living’ stages, where they are present in the environment in an infective form. The schematic presented above refers to lymphatic filarial parasites only.
NEMATODE GENOMES
Analysis of nematode phylogeny based on the small subunit ribosomal DNA sequences divided the phylum into five clades: Dorylaimia (Clade I), Enoplia (Clade II), Spurina (Clade III), Tylenchina (Clade IV) and Rhabditina (Clade V), each containing parasitic species (Blaxter et al. Reference Blaxter, De Ley, Garey, Liu, Scheldeman, Vierstraete, Vanfleteren, Mackey, Dorris, Frisse, Vida and Thomas1998). The free-living model organism C. elegans belongs to clade V, along with many important parasitic species of humans and animals, while filarial nematodes and Ascaris belong to clade III. C. elegans is commonly used to study the fundamental principles of biology and for understanding mechanisms of human disease, as well as acting as a model for parasitic nematodes (Kirienko et al. Reference Kirienko, Mani and Fay2010; Hashmi et al. Reference Hashmi, Wang, Parhar, Collison, Conca, Al-Mohanna and Gaugler2013; Li and Le, Reference Li and Le2013). In 1998, the sequencing of the C. elegans genome heralded a new era of whole-organism research, and inspired other nematode genome sequencing projects. Draft genome sequences are now available for a range of parasitic species including Brugia malayi (Ghedin et al. Reference Ghedin, Wang, Spiro, Caler, Zhao, Crabtree, Allen, Delcher, Guiliano, Miranda-Saavedra, Angiuoli and Creasy2007), Ascaris suum (Jex et al. Reference Jex, Liu, Li, Young, Hall, Li, Yang, Zeng, Xu, Xiong, Chen, Wu, Zhang, Fang, Kang, Anderson, Harris, Campbell, Vlaminck, Wang, Cantacessi, Schwarz, Ranganathan, Geldhof, Nejsum, Sternberg, Yang, Wang, Wang and Gasser2011), Trichinella spiralis (Mitreva et al. Reference Mitreva, Jasmer, Zarlenga, Wang, Abubucker, Martin, Taylor, Yin, Fulton, Minx, Yang, Warren, Fulton, Bhonagiri, Zhang, Hallsworth-Pepin, Clifton, McCarter, Appleton, Mardis and Wilson2011), Dirofilaria immitis (Godel et al. Reference Godel, Kumar, Koutsovoulos, Ludin, Nilsson, Comandatore, Wrobel, Thompson, Schmid, Goto, Bringaud, Wolstenholme, Bandi, Epe, Kaminsky, Blaxter and Maser2012), Haemonchus contortus (Laing et al. Reference Laing, Kikuchi, Martinelli, Tsai, Beech, Redman, Holroyd, Bartley, Beasley, Britton, Curran, Devaney, Gilabert, Hunt, Jackson, Johnston, Kryukov, Li, Morrison, Reid, Sargison, Saunders, Wasmuth, Wolstenholme, Berriman, Gilleard and Cotton2013; Schwarz et al. Reference Schwarz, Korhonen, Campbell, Young, Jex, Jabbar, Hall, Mondal, Howe, Pell, Hofmann, Boag, Zhu, Gregory, Loukas, Williams, Antoshechkin, Brown, Sternberg and Gasser2013), Loa loa (Desjardins et al. Reference Desjardins, Cerqueira, Goldberg, Dunning Hotopp, Haas, Zucker, Ribeiro, Saif, Levin, Fan, Zeng, Russ, Wortman, Fink, Birren and Nutman2013), Necator americanus (Tang et al. Reference Tang, Gao, Rosa, Abubucker, Hallsworth-Pepin, Martin, Tyagi, Heizer, Zhang, Bhonagiri-Palsikar, Minx, Warren, Wang, Zhan, Hotez, Sternberg, Dougall, Gaze, Mulvenna, Sotillo, Ranganathan, Rabelo, Wilson, Felgner, Bethony, Hawdon, Gasser, Loukas and Mitreva2014) and the plant parasitic nematodes Melodoigyne incognita (Abad et al. Reference Abad, Gouzy, Aury, Castagnone-Sereno, Danchin, Deleury, Perfus-Barbeoch, Anthouard, Artiguenave, Blok, Caillaud and Coutinho2008), Meloidogyne hapla (Opperman et al. Reference Opperman, Bird, Williamson, Rokhsar, Burke, Cohn, Cromer, Diener, Gajan, Graham, Houfek, Liu, Mitros, Schaff, Schaffer, Scholl, Sosinski, Thomas and Windham2008), as well as the necromenic species Pristionchus pacificus (Dieterich et al. Reference Dieterich, Clifton, Schuster, Chinwalla, Delehaunty, Dinkelacker, Fulton, Fulton, Godfrey, Minx, Mitreva, Roeseler, Tian, Witte, Yang, Wilson and Sommer2008; Rae et al. Reference Rae, Riebesell, Dinkelacker, Wang, Herrmann, Weller, Dieterich and Sommer2008) and additional Caenorhabditis species (Stein et al. Reference Stein, Bao, Blasiar, Blumenthal, Brent, Chen, Chinwalla, Clarke, Clee, Coghlan, Coulson, D'Eustachio, Fitch, Fulton, Fulton, Griffiths-Jones, Harris, Hillier, Kamath, Kuwabara, Mardis, Marra, Miner, Minx, Mullikin, Plumb, Rogers, Schein, Sohrmann, Spieth, Stajich, Wei, Willey, Wilson, Durbin and Waterston2003). In addition, the 50 helminth genomes project coordinated by the Wellcome Trust Sanger Institute aims to provide draft genomes for 50 helminths, including many important parasitic nematodes (see www.sanger.ac.uk/research/initiatives/globalhealth/research/helminthgenomes/). Furthermore, there are databases of expressed sequence tags (ESTs), which have proved to be very useful in establishing a catalogue of the mRNA transcripts expressed in various species and in different life cycle stages. Access to ESTs can be found in various online databases, the most extensive of which is NEMBASE4 (http://www.nematodes.org/nembase4/) which reports on over 223 000 nematode genes across >60 species. Nematode transcriptomics (RNA sequencing) is set to blossom in the coming years with the advent of next generation sequencing with high throughput at significantly lower cost, thus enabling the production of large datasets. The comparative analysis of multiple nematode genomes should facilitate the identification of candidate genes of interest and hopefully lead to new drug targets for many nematode pathogens.
HSP90 IN NEMATODES
Hsp90 is a highly conserved molecule across all species. For example, Hsp90 from B. malayi and Brugia pahangi are 99·9% identical. Hsp90 from humans and Brugia spp. share 77% identity, while Hsp90 from C. elegans and Brugia spp. are 84% identical. Despite this level of conservation, the function of Hsp90 seems to vary between different nematode species, as it does between normal and malignant mammalian cells (Kamal et al. Reference Kamal, Thao, Sensintaffar, Zhang, Boehm, Fritz and Burrows2003). The first indication that some nematodes may possess an atypical Hsp90 came from a study that demonstrated that C. elegans Hsp90 (DAF-21) was unable to bind to geldanamycin (GA), the prototype Hsp90 inhibitor (David et al. Reference David, Smith, Raynes, Pulcini and Whitesell2003). DAF-21 is clearly required in C. elegans, as demonstrated by the arrested phenotype of a loss of function mutant (Birnby et al. Reference Birnby, Link, Vowels, Tian, Colacurcio and Thomas2000). However, attempts to chemically inhibit DAF-21 by growth of the nematode on plates containing high levels of GA or by feeding worms on cultures of Streptomyces hygroscopicus (the actinomycete which synthesizes GA), produced no obvious phenotype (David et al. Reference David, Smith, Raynes, Pulcini and Whitesell2003). A subsequent survey of Hsp90 in 24 different nematode species demonstrated that C. elegans DAF-21 was not unique in this respect (Him et al. Reference Him, Gillan, Emes, Maitland and Devaney2009). Using extracts of a variety of free-living and parasitic nematodes, it was shown that the ability to bind GA was associated with the life history of a particular species. Nematodes that are obligate parasites or those that live in the environment enclosed within an egg were able to bind GA; in contrast, free-living species and those with free-living larval stages did not bind GA (summarized in Fig. 2). This analysis is consistent with the adaptive evolution hypothesis proposed by David et al. in their original paper. Streptomyces hygroscopicus is a soil-dwelling species and thus nematodes with free-living stages in the environment may be exposed to GA or similar inhibitors during their life cycle, driving the evolution of a GA-resistant form of Hsp90. To further test this hypothesis, the amino acid sequences of Hsp90 from 15 species of nematodes were compared to determine if any of the GA-resistant strains showed evidence of adaptive Darwinian evolution. Although there was evidence of rapid diversifying evolution of hsp90 along three separate nematode lineages, no evidence was found for amino acid changes that correlated with a change in GA-binding (Him et al. Reference Him, Gillan, Emes, Maitland and Devaney2009). Thus the rapid evolution of the hsp90 gene is presumably associated with some other important function, given its multifaceted role within the cell.

Fig. 2. Outline of nematode species and GA-binding properties. Extracts of nematodes were incubated with GA beads and pull-downs analysed by SDS-PAGE and immune-blotting. Details in Him et al. (2009).
Studies using chimeric Hsp90 molecules demonstrated that the inability of C. elegans DAF-21 to bind GA was associated with the N-terminal region (David et al. Reference David, Smith, Raynes, Pulcini and Whitesell2003). GA is a natural product of S. hygroscopicus and studies on this organism and on other microbes that produce Hsp90 inhibitors, have cast some light upon the adaptive mechanisms by which these organisms avoid self-toxicity. A combination of elegant structural and mutational studies in S. hygroscopicus (Millson et al. Reference Millson, Chua, Roe, Polier, Solovieva, Pearl, Sim, Prodromou and Piper2011) and in the fungus, Humicola fuscoatra, which synthesizes the Hsp90 inhibitor Radicicol, demonstrated the importance of naturally occurring amino acid substitutions in the N-terminal domain of Hsp90, which alter the binding of the appropriate drug. For H. fuscoatra, a single leucine to isoleucine substitution (L34I) within the N-terminal binding pocket of Hsp90 significantly altered the binding of Radicicol, while retaining the ability to bind GA and ATP and its chaperone activity (Prodromou et al. Reference Prodromou, Nuttall, Millson, Roe, Sim, Tan, Workman, Pearl and Piper2009). These studies raise the question of whether such evolutionary pressures can result in similar mutations within hsp90 of higher organisms. Despite its widespread use as a model organism, our understanding of the ecology of C. elegans in the wild remains patchy: does C. elegans live in the same ecological niche as inhibitor-producing microbes in nature? A recent study surveyed the bioavailability of over 1000 drug-like molecules in C. elegans and demonstrated that many molecules fail to accumulate in the worm and thus have no apparent activity (Burns et al. Reference Burns, Wallace, Wildenhain, Tyers, Giaever, Bader, Nislow, Cutler and Roy2010), further suggesting that protection against toxins could be a credible selection pressure. However, while this phenomenon may contribute to the lack of activity of Hsp90 inhibitors on C. elegans, it does not explain the inability of DAF-21 to bind to Hsp90 inhibitors in soluble extracts.
FUNCTIONAL STUDIES ON HSP90 IN C. ELEGANS
Hsp90 has been well characterized in yeast and cultured mammalian cell lines, as an ATP-dependent, ubiquitous, molecular chaperone, which interacts with multiple ‘client’ proteins; these are proteins which require the activity of Hsp90 for folding or stability. However, much less is known about the cellular function of Hsp90 in the complete metazoan. Inevitably, much of our understanding of the role of Hsp90 in nematodes comes from work on C. elegans. This free-living nematode is easily maintained in the laboratory setting, unlike parasitic species, which often have complex life cycles involving intermediate and definitive hosts and/or stages that develop in the environment (summarized in Fig. 1). The life cycle of C. elegans takes only 3 days and, additionally, a range of powerful molecular techniques such as RNA interference (RNAi), mutagenesis and transgenesis, can be employed in C. elegans, whereas the application of such methods to most parasitic nematodes is in its infancy.
Mammals and yeast contain two isoforms of Hsp90 present in the cytosol, while in C. elegans there is a single hsp90 orthologue, known as daf-21, located on chromosome V. Worms with a loss of function mutation in daf-21 arrest at the L2–L3 stage (Birnby et al. Reference Birnby, Link, Vowels, Tian, Colacurcio and Thomas2000) confirming the requirement for wild type DAF-21 in life cycle progression in C. elegans. Early studies demonstrated that C. elegans underwent a classical heat shock response following exposure to elevated temperature, with the induction of a number of genes, particularly those encoding small Hsps (Candido et al. Reference Candido, Jones, Dixon, Graham, Russnak and Kay1989; Jones et al. Reference Jones, Dixon, Graham and Candido1989). The expression of most Hsps in response to elevated temperature is regulated by heat shock factor (HSF-1), a transcription factor that binds to conserved heat shock elements in the upstream region of Hsp genes (Wu, Reference Wu1995). However, in C. elegans knock-down of hsf-1 by RNAi had minimal effect upon the levels of DAF-21 in nematodes exposed to heat shock, while significantly decreasing the expression of the small Hsp, Hsp-16 (Walker et al. Reference Walker, Thompson, Brawley, Scanlon and Devaney2003). These data suggest that daf-21 expression may be regulated by factors other than the classic HSF-1. Interestingly, previous studies had shown that daf-21 was up-regulated 15-fold in dauer larvae, an alternative developmental stage that C. elegans can enter upon encountering unfavourable conditions, such as overcrowding or lack of food. Dauer larvae are stress-resistant and long-lived. Entry into, and recovery from, the dauer stage is regulated by levels of a pheromone produced by the worms (Golden and Riddle, Reference Golden and Riddle1982). When worms were stimulated to exit the dauer stage, levels of daf-21 mRNA declined rapidly (Dalley and Golomb, Reference Dalley and Golomb1992). Similar findings were reported by Jones et al. (Reference Jones, Riddle, Pouzyrev, Velculescu, Hillier, Eddy, Stricklin, Baillie, Waterston and Marra2001) who showed that the expression of daf-21 was up-regulated in the dauer stage compared with mixed life cycle stages, but that this was not the case for hsp-70 (Jones et al. Reference Jones, Riddle, Pouzyrev, Velculescu, Hillier, Eddy, Stricklin, Baillie, Waterston and Marra2001). However, comparative proteomic analysis of C. elegans mixed life cycle stages and dauer larvae showed no over-expression of DAF-21 protein in dauers (Jeong et al. Reference Jeong, Na, Jeong, Chitwood, Shim and Paik2009). Why daf-21 mRNA should be so highly expressed in the dauer stage remains unclear. One possibility is that daf-21 mRNA may be accumulated in dauer larvae in readiness for the transition to normal development, where elevated levels of DAF-21 could be required to chaperone proteins required for non-dauer development. Further studies would need to examine levels of DAF-21 over a time course, as the animal exits the dauer stage. Additional evidence for a role for DAF-21 in the dauer pathway comes from studies of a gain of function daf-21 mutant. These worms bear a single amino acid substitution (E292K) in DAF-21 and are dauer-constitutive (Daf-c), i.e. enter the dauer state under favourable conditions of growth due to a defect in chemosensory ability. This Daf-c phenotype is shared with a second mutant, daf-11, which encodes a protein homologous to transmembrane guanylyl cyclases. Both the daf-11 and the daf-21 Daf-c phenotypes can be rescued using an analogue of cGMP. It was proposed that DAF-21 was required to stabilize DAF-11 or another component in the cGMP pathway (Birnby et al. Reference Birnby, Link, Vowels, Tian, Colacurcio and Thomas2000). A better understanding of the key client proteins of DAF-21 in C. elegans would help explain the requirement for Hsp90 in different life cycle stages of the nematode.
While studies from this laboratory have failed to demonstrate a significant induction of DAF-21 in C. elegans following heat shock (as quantified by immuno-blotting with a specific antibody) (Thompson et al. Reference Thompson, Cockroft, Wheatley, Britton and Devaney2001; Devaney et al. Reference Devaney, O'Neill, Harnett, Whitesell and Kinnaird2005), other studies using in situ hybridization with daf-21 specific probes have revealed differences in both the staining patterns and abundance of daf-21 mRNA in C. elegans upon heat shock (Inoue et al. Reference Inoue, Takamura, Yamae, Ise, Kawakami, Tabuse, Miwa and Yamaguchi2003). At normal growth temperatures, DAF-21 is predominantly localized to the germline of C. elegans, as detected by antibody staining (Gillan et al. Reference Gillan, Maitland, McCormack, Nik Him and Devaney2009) as well as in situ hybridization (Inoue et al. Reference Inoue, Takamura, Yamae, Ise, Kawakami, Tabuse, Miwa and Yamaguchi2003). A genome-wide RNAi screen reported that knock-down of daf-21 leads to defects in oogenesis (Piano et al. Reference Piano, Schetter, Mangone, Stein and Kemphues2000; Inoue et al. Reference Inoue, Hirata, Kuwana, Fujita, Miwa, Roy and Yamaguchi2006). In more detailed experiments from this lab, it was shown that one of the most penetrant phenotypes obtained upon daf-21(RNAi) was a protruding vulva and sterility in the F1 generation, due to a lack of gonad development (Gillan et al. Reference Gillan, Maitland, McCormack, Nik Him and Devaney2009). Interestingly, Drosophila Hsp90 is expressed in the germline (Xiao and Lis, Reference Xiao and Lis1989) and in Xenopus, Hsp90 is expressed during oogenesis suggesting a conserved function between different species (Coumailleau et al. Reference Coumailleau, Billoud, Sourrouille, Moreau and Angelier1995). While the precise function of Hsp90 in the germline is still poorly understood, identification of DAF-21 client proteins can help explain mutant phenotypes, as exemplified by the interaction between the WEE-1.3 kinase and Hsp90. Here it was shown that DAF-21 indirectly regulates the meiotic prophase/metaphase transition during oocyte development through maintaining the activity of WEE-1.3 (Myt-1 orthologue in C. elegans), which is involved in cell cycle progression (Inoue et al. Reference Inoue, Hirata, Kuwana, Fujita, Miwa, Roy and Yamaguchi2006).
The accumulation of DAF-21 in the germline of C. elegans is interesting in the context of recent studies, which have described a role for Hsp90 in various silencing pathways, including the piRNA and microRNA (miRNA) pathways (Izumi et al. Reference Izumi, Kawaoka, Yasuhara, Suzuki, Sugano, Katsuma and Tomari2013; Martinez and Gregory, Reference Martinez and Gregory2013). These studies link the original observations of Lindquist and colleagues on the capacity of Hsp90 to buffer environmental change with a possible molecular mechanism. Altering the function of Hsp90 in Drosophila by mutation or by treatment with GA resulted in flies with various abnormalities of wings and eyes (Rutherford and Lindquist, Reference Rutherford and Lindquist1998). It was proposed that high levels of cellular Hsp90 buffered pre-existing cryptic variation, which was then expressed when Hsp90 activity was compromised. However, the molecular mechanisms underlying these observations have remained elusive. Recent studies have shown that Hsp90 interacts in the Piwi pathway in Drosophila. piRNAs are a germline-specific class of small RNAs that silence transposons. Inhibition of Hsp90 function is proposed to inhibit the loading of RNA onto PIWI proteins (Izumi et al. Reference Izumi, Kawaoka, Yasuhara, Suzuki, Sugano, Katsuma and Tomari2013), resulting in the activation of transposons in Drosophila and the appearance of de novo mutations (Specchia et al. Reference Specchia, Piacentini, Tritto, Fanti, D'Alessandro, Palumbo, Pimpinelli and Bozzetti2010; Gangaraju et al. Reference Gangaraju, Yin, Weiner, Wang, Huang and Lin2011), suggesting one mechanism by which Hsp90 may regulate phenotypic change. Additional studies have implicated Hsp90 in epigenetic regulation of gene expression via its interaction with Trithorax (Trx), a chromatin remodelling factor (Ruden and Lu, Reference Ruden and Lu2008; Tariq et al. Reference Tariq, Nussbaumer, Chen, Beisel and Paro2009). Trx is associated with active transcription and Hsp90 inhibition degraded Trx and resulted in a down-regulation in the expression of a number of genes. Genome-wide studies in Drosophila cells using chromatin immunoprecipitation showed that Hsp90 associated with multiple genes and that these were transcriptionally paused. Inhibition of Hsp90 released the pause, activating gene expression (Sawarkar et al. Reference Sawarkar, Sievers and Paro2012).
Hsp90 is also involved in the loading of small interfering RNA duplexes (siRNA) and miRNA onto argonaute proteins in Drosophila and in human cells; argonaute proteins are the main components of the RNA Induced Silencing Complex (RISC). siRNA and microRNAs can only fulfil their function as post-transcriptional repressors of gene function (often referred to as the ‘fine-tuners’ of the genome) if they are successfully loaded onto these complexes (Iwasaki et al. Reference Iwasaki, Kobayashi, Yoda, Sakaguchi, Katsuma, Suzuki and Tomari2010; Miyoshi et al. Reference Miyoshi, Takeuchi, Siomi and Siomi2010). Whether Hsp90 plays similar roles in nematodes remains to be seen, but given the phenotypes of daf-21(RNAi) (F1 sterility, protruding vulva phenotype, lack of gonad development, reduced brood size) and larval arrest in a homozygous daf-21 mutant strain, it is an area which warrants further investigation.
HSP90 IN PARASITIC NEMATODES
In comparison to the information available on DAF-21 in C. elegans, relatively little is known about Hsp90 in parasitic species. hsp90 has been cloned and partially characterized from the filarial worm B. pahangi (Bp-hsp90) (Devaney et al. Reference Devaney, O'Neill, Harnett, Whitesell and Kinnaird2005) and from the clade I parasite T. spiralis (Yang et al. Reference Yang, Qin, Zarlenga, Cao and Tian2013). In common with most other parasitic nematodes, lymphatic filarial worms undergo a heat shock as part of their life cycle. The infection is transmitted to humans by the bite of a mosquito carrying the third stage larvae (L3) in the mouthparts and head. The L3 enter the lymphatics where they develop through two moults to adult male and female adult worms, which can live for approximately 8 years. Following sexual reproduction, the adult female releases microfilariae into the lymph which then migrate to the circulatory system, where they provide a reservoir of infection for the vector (summarized in Fig. 1B). The transfer of the L3 between vector and mammalian host is associated with an elevation in temperature from the ambient (mosquito) to 37 °C (mammalian host). Using 35S methionine labelling, an increase in expression of a range of heat shock proteins, including Hsp90, was observed as L3 were shifted from 28 to 37 °C (Jecock and Devaney, Reference Jecock and Devaney1992). However, there is no evidence to suggest that the expression of Bp-hsp90 (either mRNA or protein) is significantly increased following exposure of adult worms to heat shock conditions (Thompson et al. Reference Thompson, Cockroft, Wheatley, Britton and Devaney2001; Devaney et al. Reference Devaney, O'Neill, Harnett, Whitesell and Kinnaird2005), suggesting that most Bp-hsp90 is constitutively expressed. Parasitic nematodes secrete a range of molecules when cultured in serum-free medium (the so called excretory-secretory products or ES), and analysis of the ES of adult B. malayi demonstrated that B. malayi Hsp90 (Bm-Hsp90) was present (Kumari et al. Reference Kumari, Lillibridge, Bakeer, Lowrie, Jayaraman and Philipp1994), a finding which has been confirmed in this laboratory using B. pahangi (Devaney et al. unpublished observations). Hsp90 is best characterized as a cytosolic chaperone, but studies on some cancer cells have shown that it can be expressed at the cell surface and secreted into medium (Eustace and Jay, Reference Eustace and Jay2004; Eustace et al. Reference Eustace, Sakurai, Stewart, Yimlamai, Unger, Zehetmeier, Lain, Torella, Henning, Beste, Scroggins, Neckers, Ilag and Jay2004). Extracellular Hsp90 was shown to be required for tumour cell invasion via the chaperoning of matrix metalloproteinases. It is difficult to envisage an extracellular function for Bp-Hsp90 in adult B. pahangi, but it could conceivably be important for chaperoning and folding other secreted molecules.
While there are many similarities in the molecular architecture of DAF-21 and Bp-Hsp90 there are also significant differences in the function of the molecule in the respective species. As referred to above, DAF-21 does not bind GA, while Bp-Hsp90 does. The subtle complexities of the Hsp90 machinery in different nematodes was demonstrated in a study which attempted to rescue the C. elegans daf-21 mutant phenotype by heterologous expression of parasite hsp90 genes (Gillan et al. Reference Gillan, Maitland, McCormack, Nik Him and Devaney2009). Previous studies had demonstrated the efficacy of inter-species complementation experiments in transgenic C. elegans, where a parasite gene can rescue the C. elegans mutant phenotype and restore wild type traits (Kwa et al. Reference Kwa, Veenstra, Van Dijk and Roos1995; Britton and Murray, Reference Britton and Murray2002; Couthier et al. Reference Couthier, Smith, Mcgarr, Craig and Gilleard2004; Massey et al. Reference Massey, Bhopale, Li, Castelletto and Lok2006). However, despite a high level of amino acid homology (84% identical, 91% similar), expression of a Bp-hsp90 transgene could not rescue a C. elegans daf-21 mutant. Similarly, no rescue was observed by expression of the Bp-hsp90 construct in wild type C. elegans, in which endogenous daf-21 levels were reduced by RNAi. In these experiments, Bp-hsp90 was introduced into C. elegans by microinjection of a plasmid construct containing the Bp-hsp90 coding sequence under the control of the C. elegans daf-21 promoter and 3′UTR regions, in an attempt to mimic the expression of the endogenous gene as closely as possible. To confirm that the parasite gene was efficiently transcribed in C. elegans, injection of a Bp-hsp90 LacZ translational reporter construct demonstrated that the parasite Hsp90 was expressed in most tissues of the transformed worms and, importantly, that the B. pahangi protein expressed in C. elegans bound to GA beads in a pull-down experiment (Gillan et al. Reference Gillan, Maitland, McCormack, Nik Him and Devaney2009). Despite these findings, complementation of the daf-21 mutant or daf-21(RNAi) worms was never observed. A more successful inter-species rescue was obtained using hsp90 from H. contortus (Hc-hsp90), a trichostrongyloid nematode of sheep, which belongs to the same clade as C. elegans. Hc-Hsp90 is 88% identical and 93% similar to DAF-21 and experiments with an Hc-hsp90 construct provided partial rescue of the daf-21 mutant phenotypes. As control experiments had demonstrated that complementation with the wild-type daf-21 in the same injection cassette restored the wild type phenotype of the daf-21 mutant worms, a ‘gradient’ of rescue was proposed, i.e. despite high sequence homology the B. pahangi transgene was unable to rescue the mutant, Hc-hsp90 was able to confer partial rescue but it required complementation with the wild-type C. elegans gene to completely rescue the mutant phenotypes and restore fertility (see Table 1 for summary). These results suggest that in metazoans, the ability to complement Hsp90 function may depend on factors other than sequence homology; successful rescue may require specific co-chaperones for full function of the transgene, or alternatively these data may reflect difference in the ability of the respective Hsp90 proteins to chaperone key client proteins. In contrast to the studies on nematode Hsp90, other inter-species complementation experiments using mutant Saccharomyces cerevisiae have been successful even when the degree of homology between the respective hsp90 genes was relatively limited (see Table 2). Indeed, both C. elegans wild-type daf-21 and human hsp90b were able to rescue an S. cerevisiae hsp90 mutant despite sharing only 60·5 and 60·3% sequence homology, respectively. However, in this case complementation required the presence of STI-1/Hop (Hsp organizing protein) indicating that the expression of co-chaperone proteins is of primary importance to the functionality of Hsp90 (Piper et al. Reference Piper, Panaretou, Millson, Trumana, Mollapour, Pearl and Prodromou2003).
Table 1. Summary of hsp90 inter-species rescue experiments in C. elegans. Extracted from Gillan et al. (Reference Gillan, Maitland, McCormack, Nik Him and Devaney2009)

Table 2. Summary of hsp90 inter-species rescue experiments in S. cerevisiae. Extracted from Palmer et al. (Reference Palmer, Louvion, Tibbetts, Engman and Picard1995); Piper et al. (Reference Piper, Panaretou, Millson, Trumana, Mollapour, Pearl and Prodromou2003); Wider et al. (Reference Wider, Peli-Gulli, Briand, Tatu and Picard2009)

HSP90 CO-CHAPERONES IN NEMATODES
The interaction of Hsp90 with client and co-chaperone proteins is regulated by ATP-induced conformational changes in Hsp90, resulting in the folding and activation of substrate or ‘client’ proteins. Over the years, the number of Hsp90 client proteins has grown significantly and it has been shown to be essential in the maturation of many different types of protein in mammalian cells, including transcription factors, steroid receptors, serine/threonine and tyrosine kinases. A list of Hsp90 client proteins is curated by The Picard laboratory (http://www.picard.ch). There are no published data from parasitic nematode species on the client proteins that require Hsp90 for function, but there is an extensive list of both predicted and confirmed clients for DAF-21 (http://www.wormbase.org). Many of these clients have homologues in the human system, such as small ribosomal proteins, serine/threonine kinases and cyclin-dependent kinases, in addition to some ‘worm-specific’ proteins, such as those essential for pharyngeal pumping, a process required for food intake. While it is likely that at least some C. elegans client proteins will be similar in other nematodes, the degree of conservation is currently unknown.
The identification and function of co-chaperones in different species is of interest given their importance in the outcome of client protein maturation. For example, some Hsp90 clients require a select group of co-chaperones and alternative co-chaperones may have differential effects on client proteins (Riggs et al. Reference Riggs, Cox, Cheung-Flynn, Prapapanich, Carrigan and Smith2004; Felts et al. Reference Felts, Karnitz and Toft2007; Smith and Toft, Reference Smith and Toft2008). These findings are strengthened by evidence that many co-chaperones display mutually exclusive binding to Hsp90 (Pratt et al. Reference Pratt, Galigniana, Harrell and Defranco2004; Harst et al. Reference Harst, Lin and Obermann2005; Pearl and Prodromou, Reference Pearl and Prodromou2006). A study by Johnson and Brown illustrated the plasticity of the co-chaperone machinery in diverse eukaryotes and showed that the architecture of the Hsp90/co-chaperone machine varied in a species-specific manner. Their results suggested that with increasing complexity of the organism, additional co-factors may be required. While Hsp90 is universally required in metazoans, it is likely that the Hsp90 complex varies in different tissues/cells. Caenorhabditis elegans was shown to express orthologues of nine of the ten human Hsp90 co-chaperones used in the directed Psi-BLAST search against the complete C. elegans genome (Johnson and Brown, Reference Johnson and Brown2009). The only co-chaperone absent in C. elegans was Cyp40, which was first identified in mammalian steroid receptor complexes (Riggs et al. Reference Riggs, Cox, Cheung-Flynn, Prapapanich, Carrigan and Smith2004). However, another striking difference in C. elegans was shown to occur within the sequence of the adaptor protein Hop (STI-1 in C. elegans), which has an essential role in mediating interactions between Hsp70 and Hsp90. In most eukaryotes, Hop consists of five domains: three tetratricopeptide repeat (TPR) domains and two dipeptide (DP) repeats, which act as linker domains, arranged TPR1–DP1–TPR2A–TPR2B–DP2 (Scheufler et al. Reference Scheufler, Brinker, Bourenkov, Pegoraro, Moroder, Bartunik, Hartl and Moarefi2000). However, C. elegans STI-1 lacks the TPR1 and DP1 domains (www.wormbase.org). This is an intriguing finding as, in other systems, the TPR1 domain has been shown to be essential for binding Hsp70, and for linking Hsp70 with Hsp90 (which binds Hop via TPR2A) (Scheufler et al. Reference Scheufler, Brinker, Bourenkov, Pegoraro, Moroder, Bartunik, Hartl and Moarefi2000). Hsp70 was observed to bind to C. elegans STI-1 despite the lack of the hypothetical binding site; however, in the presence of DAF-21, Hsp70 binding was completely abrogated suggesting that in C. elegans there may be no requirement for both chaperones to interact simultaneously, distinguishing C. elegans from other species (Gaiser et al. Reference Gaiser, Brandt and Richter2009). The lack of the TPR1 domain was also observed in the closely related Caenorhabditis briggsae and Caenorhabditis remanei. Interestingly, it appears this atypical Hop is not limited to the Caenorhabditis genus, as the B. malayi genome contains an orthologue of C. elegans STI-1, BMA-STI-1 (http://www.wormbase.org). This protein shares 65·5% amino acid identity with C. elegans STI-1 and similarly appears to lack TPR1 and DP1 domains suggesting that this could be a nematode-wide scenario (see Fig. 3).

Fig. 3. Comparison of the domain structures of Hop from divergent species. TPR domains are shown in blue and DP domains in pink. Homologues of Hop show differences in domain organization. TPR1 and DP1 are absent in the nematode species C. elegans and B. malayi while only DP1 is lacking in D. melanogaster. All five domains are present in H. sapiens and S. cerevisiae. Adapted from Gaisier et al. (2009).
In 2007, a genome-wide promoter analysis addressed the tissue-specific expression of most proteins in C. elegans and showed that DAF-21 was expressed in many different cell types. This analysis allowed the correlation of DAF-21 expression with the expression profiles of its presumed co-chaperone proteins (Dupuy et al. Reference Dupuy, Bertin, Hidalgo, Venkatesan, Tu, Lee, Rosenberg, Svrzikapa, Blanc, Carnec, Carvunis, Pulak, Shingles, Reece-Hoyes, Hunt-Newbury, Viveiros, Mohler, Tasan, Roth, Le Peuch, Hope, Johnsen, Moerman, Barabasi, Baillie and Vidal2007). Analysing the differential expression of DAF-21 and its co-chaperones allows for some speculation on the cellular activity of the Hsp90/co-chaperone complexes, as it is likely that tissue localization corresponds to function (data accessed through www.wormbase.org). For example, co-chaperones P23 (ZC395.1), Hop (STI-1) and FKB-6 were all shown to be ubiquitously expressed in C. elegans, whereas PP5 (PPH-5) and CDC-37 are expressed only in the intestine and embryo respectively, suggesting a more specific role. Perhaps the best example of cellular location correlating with function is illustrated by the expression pattern of the TPR-containing cofactor UNC-45 (uncoordinated), which is expressed in body wall muscle, intestinal muscle and vulval muscles. In C. elegans, the 95 rhomboid-shaped body wall muscle cells comprise one of the major tissues of the adult worm (www.wormatlas.org). Down-regulation of UNC-45 leads to paralysis via decreased myosin assembly, whereas an excess of UNC-45 results in myosin accumulation, both of which result in defective myofibril organization (Barral et al. Reference Barral, Hutagalung, Brinker, Hartl and Epstein2002; Landsverk et al. Reference Landsverk, Li, Hutagalung, Najafov, Hoppe, Barral and Epstein2007). The details of how UNC-45 and DAF-21 work together are still unclear, although it has been reported that UNC-45 interacts with Hsp90 via a TPR domain (Russell et al. Reference Russell, Whitt, Chen and Chinkers1999; Scheufler et al. Reference Scheufler, Brinker, Bourenkov, Pegoraro, Moroder, Bartunik, Hartl and Moarefi2000; Barral et al. Reference Barral, Hutagalung, Brinker, Hartl and Epstein2002) and myosin motor domains through its COOH-terminal regions (Barral et al. Reference Barral, Bauer, Ortiz and Epstein1998, Reference Barral, Hutagalung, Brinker, Hartl and Epstein2002). Elegant studies by Ni et al. demonstrated that DAF-21 and UNC-45 interact in pull-down experiments using C. elegans lysates and proposed that this interaction might have an inhibitory effect on the myosin-chaperoning activity of UNC-45 (Ni et al. Reference Ni, Hutagalung, Li and Epstein2011).
Online sources (http://www.wormbase.org) demonstrate that C. elegans encodes 80 TPR proteins, so perhaps the fact that only nine are known homologues of human Hsp90 co-chaperones is surprising (for comparison, the proteome of S. cerevisiae contains about 25 proteins with TPR domains, seven of which are confirmed to interact with either Hsp90 or Hsp70). In a recent study, Haslbeck et al. analysed previously uncharacterized TPR proteins, known to be associated with the Hsp90/Hsp70 complex. The entire proteome of C. elegans was analysed and scores were given to each protein based on the level of homology to known co-chaperone TPR proteins of Homo sapiens, S. cerevisiae and C. elegans. The proteins with the highest score for potential binding to Hsc70/Hsp70 and Hsp90 were the ten C. elegans TPR proteins used in the sample set (SGT-1, UNC-45, FKB-6, PPH-5, STI-1, HIP-1, CHN-1, C56C10.10/AIP-1, C17G10.10/CNS-1). In addition, three uncharacterized C. elegans open-reading frames with homologues in Drosophila or humans emerged as possible Hsp90 interactors. Two of these, C34B2.5 and ZK370.8 were shown to bind both Hsc70 and DAF-21 with low micromolar affinities; mutation of amino acids in the Hsp90 binding site for TPR proteins (EEVD sequence) was shown to disrupt the interaction. Interestingly, this study demonstrates that the majority of TPR proteins in C. elegans have no binding affinity for DAF-21 (Haslbeck et al. Reference Haslbeck, Eckl, Kaiser, Papsdorf, Hessling and Richter2013).
HSP90 AS A DRUG TARGET IN NEMATODES
The realization that Hsp90 acted as a chaperone for various oncogenic proteins led to a major effort to develop novel small molecule inhibitors of Hsp90 for use in various tumours (Neckers et al. Reference Neckers, Mimnaugh and Schulte1999). More recently the potential of Hsp90 inhibitors as novel chemotherapeutic agents for various parasitic infections has been studied (see other articles in this special issue). As referred to previously, there is a need for novel drugs to treat humans and animals infected with helminth parasites and studies in vitro have shown a dependence on Hsp90 in parasitic nematodes (Devaney et al. Reference Devaney, O'Neill, Harnett, Whitesell and Kinnaird2005) and in the trematode parasite Schistosoma japonicum (Wenkert et al. Reference Wenkert, Ramirez, Shen and Kron2010). The design of new anthelmintics generally centres on attempts to establish and exploit molecular targets that are exclusive to the parasite, with the aim of minimizing potential damage to the host. Although this approach can be successful, an alternative course is to target a common pathway shared by the infectious agent and the host, which has evolved over time to perform different functions unique to the respective species. The benefits of focusing drug discovery on such pathways are two-fold; the scope for identifying new targets is broadened and not confined to parasite-specific molecules, many of which are hypothetical proteins of unknown function, and the ubiquity of such molecules in other systems may allow for drugs designed in this way to be used in other parasitic infections. The repurposing of compounds developed to treat other conditions is an attractive proposition for drug development for neglected tropical diseases, where the potential to recoup drug development costs is limited. In this respect, Hsp90 inhibitors have been shown to be effective against a variety of tropical pathogens including Plasmodium (Kumar et al. Reference Kumar, Musiyenko and Barik2003; Shahinas et al. Reference Shahinas, Liang, Datti and Pillai2010, Reference Shahinas, Folefoc, Taldone, Chiosis, Crandall and Pillai2013), Trypanosoma (Pallavi et al. Reference Pallavi, Roy, Nageshan, Talukdar, Pavithra, Reddy, Venketesh, Kumar, Gupta, Singh, Yadav and Tatu2010) and Leishmania (Petersen et al. Reference Petersen, Guedes, Versoza, Lima, De Freitas, Borges and Veras2012).
Recent studies from this laboratory have focused on the potential of Hsp90 inhibitors as macrofilaricidal agents in filarial infection. Currently, the drugs used to control filarial nematodes in mass drug administration (MDA) programmes, such as diethylcarbamazine (DEC) and ivermectin, largely target the circulating microfilariae (Mf), resulting in reduced transmission rates. However, Mf repopulate the circulation necessitating continued treatment over the long reproductive lifespan of adult worms, incurring significant costs for control programmes and increasing the chances of resistance emerging (Prichard et al. Reference Prichard, Basanez, Boatin, McCarthy, Garcia, Yang, Sripa and Lustigman2012). It has long been a goal of WHO to identify a suitable drug that kills adult filarial worms but, to date, no such agent has been developed (Molyneux et al. Reference Molyneux, Bradley, Hoerauf, Kyelem and Taylor2003). At higher concentrations of drug, DEC can have macrofilaricidal activity, although in most long-term studies, Mf were shown to re-emerge in the blood of some patients, 1–5 years post-treatment. This suggests either that a proportion of adult worms survived DEC treatment and recover Mf production or may reflect re-infection (Terhell et al. Reference Terhell, Haarbrink, Van Den Biggelaar, Mangali, Sartono and Yazdanbakhsh2003). Targeting the Wolbachia intracellular endosymbiont of many pathogenic filarial worms with doxycycline remains the most promising new therapy that affects adult worms. However, this antibiotic is contraindicated in pregnant women and children and prolonged dosing is not ideal for MDA, but the search continues for a more satisfactory antibiotic (Hoerauf, Reference Hoerauf2008). While the B. malayi genome may yet illuminate novel parasite-specific drug targets, we have previously identified a requirement for Hsp90 as a possible weak spot in filarial nematodes. The prototype Hsp90 inhibitor, GA, a naturally occurring benzoquinone ansamycin, binds the N-terminal ATP pocket of Hsp90 disrupting its function and resulting in the degradation and/or inhibition of client proteins. In initial studies, in vitro exposure of B. pahangi adult female worms to 1 μ m GA (a concentration chosen as it inhibits mammalian Hsp90 activity) was shown to result in a significant reduction in Mf production from adult worms after 24 h and by 48 h, Mf release had ceased completely (Devaney et al. Reference Devaney, O'Neill, Harnett, Whitesell and Kinnaird2005). As well as inhibiting Mf output, GA was shown to kill adult worms, as after 7 days of exposure to drug, 100% of adult female worms were dead. GA probably has a specific effect on adult worms (as well as inhibition of Mf output), as further experiments demonstrated that adult males were also killed by exposure to 1·0 μ m GA. As referred to above, most species of filarial worms are known to harbour endosymbiotic bacteria, and to ensure that these effects were not a result of GA targeting the Wolbachia of B. pahangi, the experiments were repeated using the Wolbachia-free species Acanthocheilonema viteae and similar results were obtained. Comparable data were produced by Wenkert et al. who tested GA and a number of GA-derivatives against adult B. malayi in vitro. In their study, all four derivatives tested were active at concentrations down to 500 nm. While these results show promise, GA suffers from limitations as a chemotherapeutic agent due to unacceptable levels of hepatoxicity and would not be suitable for use in filarial infection. Thus most recent studies on Hsp90 inhibitors have focused on novel small molecule inhibitors (Wenkert et al. Reference Wenkert, Ramirez, Shen and Kron2010).
In subsequent studies, we investigated whether members of the purine-scaffold series of Hsp90 inhibitors, developed by the Chiosis laboratory (Chiosis and Tao, Reference Chiosis and Tao2006; Taldone et al. Reference Taldone, Zatorska, Patel, Zong, Rodina, Ahn, Moulick, Guzman and Chiosis2011), would have activity against adult B. pahangi. In initial experiments, a fluorescence polarization (FP) assay, originally developed as a high-throughput screen for the detection of small molecule inhibitors of Hsp90 in tumour cells, was adapted to Brugia sp. (Taldone et al. Reference Taldone, Gillan, Sun, Rodina, Patel, Maitland, O'Neill, Chiosis and Devaney2010). The assay is based on the ability of small molecules to compete with the binding of fluorescently labelled GA to Hsp90 in cellular homogenates, thus negating the need for the production of purified recombinant protein as well as enabling investigation of Hsp90 in its native conformation. Previous studies had demonstrated that Hsp90 in tumour cells is present in multi-chaperone complexes with high ATPase activity and a higher affinity for N-terminal inhibitors compared with Hsp90 from normal cells, where it is present in a latent form with a reduced affinity for these inhibitors (Kamal et al. Reference Kamal, Thao, Sensintaffar, Zhang, Boehm, Fritz and Burrows2003). Subsequent studies confirmed this finding and showed that Hsp90-specific pull-downs using the purine scaffold compound, PU-H71 conjugated to a solid support, identified a complex of interacting proteins from tumour cells (Moulick et al. Reference Moulick, Ahn, Zong, Rodina, Cerchietti, Gomes Dagama, Caldas-Lopes, Beebe, Perna, Hatzi, Vu, Zhao, Zatorska, Taldone, Smith-Jones, Alpaugh, Gross, Pillarsetty, Ku, Lewis, Larson, Levine, Erdjument-Bromage, Guzman, Nimer, Melnick, Neckers and Chiosis2011). Interestingly, the affinity of Bp-Hsp90 for GA was shown to be similar to tumour cell Hsp90 in the FP assay, while extracts of C. elegans did not show significant binding, consistent with previous results. In addition, some selectivity was noted in the binding of the purine-scaffold compounds to Bp-Hsp90 (Taldone et al. Reference Taldone, Gillan, Sun, Rodina, Patel, Maitland, O'Neill, Chiosis and Devaney2010); a 3-fold change in selectivity ratio was observed between PU-H71 and PU-WS10, two compounds with almost identical structure, suggesting that the FP assay could identify molecules which may specifically target parasite Hsp90.
In more recent work from this laboratory, we examined whether Hsp90 inhibitors might affect adult worms in vivo (Gillan et al. Reference Gillan, O'Neill, Maitland, Sverdrup and Devaney2014). For this purpose we selected an isoxazole inhibitor, NVP-AUY922, which had previously been demonstrated to exhibit anti-tumour activity in a mouse xenograft tumour model (Eccles et al. Reference Eccles, Massey, Raynaud, Sharp, Box, Valenti, Patterson, De Haven Brandon, Gowan, Boxall, Aherne, Rowlands, Hayes, Martins, Urban, Boxall, Prodromou, Pearl, James, Matthews, Cheung, Kalusa, Jones, McDonald, Barril, Brough, Cansfield, Dymock, Drysdale, Finch, Howes, Hubbard, Surgenor, Webb, Wood, Wright and Workman2008; Garon et al. Reference Garon, Finn, Hamidi, Dering, Pitts, Kamranpour, Desai, Hosmer, Ide, Avsar, Jensen, Quadt, Liu, Dubinett and Slamon2013). Experiments in vitro had shown NVP-AUY922 to be remarkably active against both adult worms and Mf at very low concentrations, killing Mf at concentrations down to 1·56 nm and 50% of adult worms at 25 nm. Using a model system in which adult B. pahangi worms are transplanted into the peritoneal cavity of mice (Devaney et al. Reference Devaney, Gillan, Wheatley, Jenson, O'connor and Balmer2002), we demonstrated that administration of three doses of NVP-AUY922 at 50 mg kg−1 effectively killed adult worms (Gillan et al. Reference Gillan, O'Neill, Maitland, Sverdrup and Devaney2014). No weight loss or other deleterious effects were observed in treated animals over the time course. While these experiments provide proof of principle that inhibition of Hsp90 is lethal to adult filarial worms in vivo, further experiments are required with different routes of drug administration and different doses of drug.
CONCLUSIONS
Recent studies on the free-living model nematode C. elegans have shed new light on the mechanisms of Hsp90 function in metazoans, particularly in relation to the tissue-specific requirement for certain co-chaperones. Additional studies will be required to fully explain the apparent resistance of C. elegans to Hsp90 inhibitors and the molecular mechanisms underlying this phenomenon. However, given the studies reviewed above it is likely that the function of Hsp90 differs between free-living nematodes and their parasitic counterparts. It is possible that part of this discrepancy will be explained by differences in the Hsp90 interactome in free-living and parasitic species, such as filarial worms, perhaps equivalent to the differences in Hsp90 function in normal versus tumour cells. While aspects of their basic biology, such as moulting, are conserved amongst all nematodes, there are significant differences between free-living and parasitic species in life cycles, modes of transmission, reproduction and dependence on hosts. As momentum to eliminate neglected tropical diseases grows, the prospect of repositioning existing drugs is an appealing one. However, while preliminary results show that chemical inhibition of Hsp90 is lethal to adult filarial worms, further research is required to determine whether Hsp90 truly represents an ‘Achilles heel’ in filarial nematodes, suitable for exploitation as a chemotherapeutic target.
ACKNOWLEDGEMENTS
We acknowledge the support for our studies on Hsp90 from the Wellcome Trust (grant number 076734/Z/05/Z) and the BBSRC (grant number BB/E013473/1) and we would like to thank Dr Collette Britton and Dr Jane Kinnaird for critical reading of this manuscript.