


INTRODUCTION
Triatomines are blood-sucking arthropods belonging to the Hemiptera order and the Reduviidae family. All stages are strictly haematophagous, female and male adults included, and take considerable amounts of blood due to their large size (4 cm for the largest species) (Lazzari et al. Reference Lazzari, Pereira, Lorenzo, Lazzari, Pereira and Lorenzo2013). A large majority of the 140 described species are endemic to the Americas; however, some species can be found in the Caribbean islands or South Asia (Dujardin et al. Reference Dujardin, Lam, Khoa and Schofield2015; Galvão and Justi, Reference Galvão and Justi2015). Most species are sylvatic, but some tend to be invasive, even domiciliated (Lazzari et al. Reference Lazzari, Pereira, Lorenzo, Lazzari, Pereira and Lorenzo2013). Indeed, they are found in close association with small mammals and birds, in their nests, in caves and rock fissures, and also in urbanized environments, as they easily prosper in the crevices of houses (Lazzari et al. Reference Lazzari, Pereira, Lorenzo, Lazzari, Pereira and Lorenzo2013). These arthropods are largely studied in South America, where they are responsible for the transmission of Trypanosoma cruzi, causative agent of Chagas disease. Consequent to the introduction of T. cruzi-infected kissing bugs’ feces in the skin, this disease, also known as American trypanosomiasis, is responsible for heart failure 10–30 years post-infection for almost 30% of affected individuals. This is the most important human parasitic disease of South America (Longo and Bern, Reference Longo and Bern2015).
Arthropod identification has a crucial importance in surveillance and the fight against vector-borne diseases. To investigate clinical cases and for epidemiological studies, accurate identification of the arthropod involved is a crucial step in evaluating the risks of infectious diseases (Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016). To circumvent the limitations of morphological analysis, such as entomological expertise and access to documentation (Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016), molecular methods have emerged in recent years for the purpose of triatomine identification (Monteiro et al. Reference Monteiro, Escalante and Beard2001) as well as that of other arthropod families (Folmer et al. Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994; Lv et al. Reference Lv, Wu, Zhang, Chen, Feng, Yuan, Jia, Deng, Wang, Wang, Mei and Lin2014). However, routine molecular biology requires extensive facilities, with high operational costs, which can be a major limitation in developing countries (Lo et al. Reference Lo, Fall, Sambe-Ba, Diawara, Gueye, Mediannikov, Sokhna, Faye, Diemé, Wade, Raoult and Fenollar2015).
In the last few years, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) has been used for the protein-profiling identification of bacteria and other microorganisms (Singhal et al. Reference Singhal, Kumar, Kanaujia and Virdi2015). Based on the querying of a comprehensive database that includes reference spectra of unambiguously identified organisms, it has been recently applied to the identification of several arthropod vectors, such as mosquitoes (Yssouf et al. Reference Yssouf, Socolovschi, Flaudrops, Ndiath, Sougoufara, Dehecq, Lacour, Berenger, Sokhna, Raoult and Parola2013b, Reference Yssouf, Parola, Lindström, Lilja, L'Ambert, Bondesson, Berenger, Raoult and Almeras2014a; Dieme et al. Reference Dieme, Yssouf, Vega-Rúa, Berenger, Failloux, Raoult, Parola and Almeras2014b), ticks (Yssouf et al. Reference Yssouf, Flaudrops, Drali, Kernif, Socolovschi, Berenger, Raoult and Parola2013a), fleas (Yssouf et al. Reference Yssouf, Socolovschi, Leulmi, Kernif, Bitam, Audoly, Almeras, Raoult and Parola2014b) and other arthropod vectors (Kaufmann et al. Reference Kaufmann, Ziegler, Schaffner, Carpenter, Pflüger and Mathis2011, Reference Kaufmann, Schaffner, Ziegler, Pflüger and Mathis2012; Hoppenheit et al. Reference Hoppenheit, Murugaiyan, Bauer, Clausen and Roesler2014; Sambou et al. Reference Sambou, Aubadie-Ladrix, Fenollar, Fall, Bassene, Almeras, Sambe-Ba, Perrot, Chatellier, Faye, Parola, Wade, Raoult and Mediannikov2015; Lafri et al. Reference Lafri, Almeras, Bitam, Caputo, Yssouf, Forestier, Izri, Raoult and Parola2016; Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016), from laboratory or collection specimens, collected from the environment or from patients. The objective of the present study was to assess the effectiveness of MALDI-TOF MS for the rapid differentiation of triatomine species. We aim to establish a triatomine reference database and to evaluate the efficacy of this new identification tool on dry field samples collected from French Guiana.
MATERIALS AND METHODS
Triatomine insects
Fresh Triatoma infestans and Rhodnius prolixus specimens from laboratory-rearing were used to standardize and evaluate the MALDI-TOF MS-based identification tool. They were reared in our laboratory (Aix-Marseille University, Marseille, France) using standard methods, at a temperature of 26 ± 1 °C, relative humidity of 70 ± 10% and a 12 h light/12 h dark cycle in incubators (Panasonic cooled incubator). Blood meals were given through a Parafilm-membrane and an artificial feeding device (Hemotek membrane feeding systems, Discovery Workshops, UK), using fresh human blood obtained from the French Blood Establishment. Ethical approval for the use of in vitro human blood was obtained from the Laboratory Research Ethics Board for Molecular Hematology, of the French Blood Establishment. Dry T. infestans, R. prolixus, Rhodnius robustus, Rhodnius pictipes, Panstrongylus geniculatus, Panstrongylus lignarius, Panstrongylus rufotuberculatus and Eratyrus mucronatus species were used to assess the performance of the method on dry specimens (Fig. 1). All specimens were unambiguously identified by one of the authors (JMB) with his identification key for French Guiana triatomines (Berenger et al. Reference Berenger, Pluot-Sigwalt, Pagés, Blanchet and Aznar2009).

Fig. 1. Pictures of triatomine species included in this study. Triatoma infestans, Rhodnius prolixus, Rhodnius robustus, Rhodnius pictipes, Pasntrongylus geniculatus, Panstrongylus lignarius, Panstrongylus rufotuberculatus, Eratyrus mucronatus.
From each sample, one femur was dissected with a sterile surgical blade. The remaining body parts were stored at −20 °C as a backup sample.
MALDI-TOF sample preparation
All living triatomines were killed by storage at −80 °C for 10 min just before the experiment. All specimens were rinsed in sterile water and dried on paper at room temperature. The femur of the left median leg was then dissected. The triatomine femurs were crushed by a Tissue Lyser instrument (Qiagen, Hilden, Germany) at a frequency of 30 movements per second for two cycles of 3 min each in 40 µL of 70% formic acid and 40 µL of 50% acetonitrile in 1·5 mL microcentrifuge tubes. All homogenates were centrifuged at 7168 RCF for 30 s, and 1 µL of each supernatant was spotted onto a steel target plate (Bruker Daltonics, Germany) in quadruplicate. One microlitre of a CHCA matrix suspension composed of saturated α-cyano-4-hydroxycinnamic acid (Sigma), 50% acetonitrile, 10% trifluoroacetic acid and high-performance liquid chromatography-grade water was directly spotted onto each sample on the target plate, to allow co-crystallization (Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016). The target plate was dried for several minutes at room temperature before insertion into the MALDI-TOF MS instrument.
MALDI-TOF MS parameters
Protein mass profiles were obtained using a Microflex LT MALDI-TOF Mass Spectrometer (Bruker Daltonics, Germany), with detection in the linear positive-ion mode at a laser frequency of 50 Hz within a mass range of 2–20 kDa. The acceleration voltage was 20 kV, and the extraction delay time was 200 ns. Each spectrum corresponds to the ions obtained from 240 laser shots performed in six regions of the same spot and were automatically acquired using the AutoXecute feature of the Flex Control v.2.4 software package (Bruker Daltonics). The average spectrum profiles obtained were visualized with Flex analysis v.3.3 software and exported to ClinProTools software v.2.2 and MALDI-Biotyper v.3.0. (Bruker Daltonics, Germany) for data processing (smoothing, baseline subtraction and peak picking) as previously described.
MALDI-TOF MS analysis and reference database creation
MALDI-TOF's performance was first assessed on fresh specimens of T. infestans and R. prolixus. Spectra reproducibility of femurs from triatomine specimens was evaluated by comparing the average spectra of each sample within its respective species using ClinProTools 2.2 and Flex analysis v.3.3 softwares (Bruker Daltonics, Germany). Reference spectra (MSP, Main Spectrum Profile) were generated by the automated function of the MALDI-Biotyper software v3.0 (Bruker Daltonics, Germany) by combining the results of the spectra of at least three specimens per condition in order to create a database. MSP were produced based on an unbiased algorithm taking information on the peak position, intensity and frequency. The consistency of MS profiles between males and females of the same species but also from the different developmental stages of the same species was assessed by comparison of the MS profiles with Flex Analysis. Six T. infestans and nine R. prolixus random spectra of good intensity and good reproducibility from both species were included in the reference spectra database. The consistency of MS profiles between fresh and dry specimens was assessed by blind test comparison of fresh and dry T. infestans and R. prolixus leg MS spectra. Random spectra of good intensity and good reproducibility of specimens from the other species for which we had more than six specimens were loaded into the MALDI-Biotyper 3.0 software to create a reference spectrum database. This way, we upgraded the database with nine new reference spectra from R. pictipes (n = 3), R. robustus (n = 3) and P. geniculatus (n = 3). This database was formerly composed of a total of six tick species (Amblyomma variegatum infected by Rickettsia africae, Rhipicephalus sanguineus, Hyalomma rufipes, Ixodes ricinus, Dermacentor marginatus and Dermacentor reticulatus), 30 mosquito species (Anopheles gambiae molecular form M and An. gambiae molecular form S, Anopheles funestus, Anopheles ziemanni, Anopheles arabiensis, Anopheles wellcomei, Anopheles rufipes, Anopheles pharoensis, Anopheles coustani, Anopheles claviger, Anopheles hyrcanus, Anopheles maculipennis, Culex quinquefasciatus, Culex pipiens, Culex modestus, Culex insignis, Culex neavei, Aedes albopictus, Aedes excrucians, Aedes vexans, Aedes rusticus, Aedes dufouri, Aedes cinereus, Aedes fowleri, Aedes aegypti, Aedes caspius, Mansonia uniformis, Orthopodomyia reunionensis, Coquillettidia richiardii and Lutzia tigripes), and other arthropods, including lice (Pediculus humanus corporis), triatomines (T. infestans) and bedbugs (Cimex lectularius), as well as the spectra obtained from the bodies (without the abdomens) of five flea species (Ctenocephalides felis, Ctenocephalides canis, Archaeopsylla erinacei, Xenopsylla cheopis and Stenoponia tripectinata) (Yssouf et al. Reference Yssouf, Socolovschi, Flaudrops, Ndiath, Sougoufara, Dehecq, Lacour, Berenger, Sokhna, Raoult and Parola2013b, Reference Yssouf, Parola, Lindström, Lilja, L'Ambert, Bondesson, Berenger, Raoult and Almeras2014a, Reference Yssouf, Socolovschi, Leulmi, Kernif, Bitam, Audoly, Almeras, Raoult and Parolab, Reference Yssouf, Almeras, Berenger, Laroche, Raoult and Parola2015a, Reference Yssouf, Almeras, Raoult and Parola2016; Dieme et al. Reference Dieme, Yssouf, Vega-Rúa and Berenger2014a; Sambou et al. Reference Sambou, Aubadie-Ladrix, Fenollar, Fall, Bassene, Almeras, Sambe-Ba, Perrot, Chatellier, Faye, Parola, Wade, Raoult and Mediannikov2015; Kumsa et al. Reference Kumsa, Laroche, Almeras, Mediannikov, Raoult and Parola2016; Lafri et al. Reference Lafri, Almeras, Bitam, Caputo, Yssouf, Forestier, Izri, Raoult and Parola2016).
Blind test validation
A total of 32 spectra from the different triatomine species were introduced into the database as reference spectra. For the species whose spectra differ according to developmental stage and/or sex, reference spectra for each variable condition were introduced in the database. The remaining 108 spectra were all selected to query the upgraded database. Since the protein extracted from each specimen was spotted on the MALDI-TOF target plate in quadruplicate, each specimen was associated with four spectra. The results of the database queries are presented as log score values (LSVs) for each spectrum, corresponding to a matched degree of signal intensities of mass spectra of the query and the reference spectra. LSVs range from 0 to 3. LSVs allow good evaluation of the reproducibility between a queried spectrum and a reference spectrum as it is the result of thorough comparison of peaks position and intensity between those two spectra. A LSV was obtained for each spectrum of the samples tested blindly. For each specimen, the spectrum with the highest LSVs was selected for identification. Up to three levels of identification were assessed here. Identification was considered perfect if species, stage and sex (if applicable) were identified correctly in blind test analyses.
Cluster analysis
In order to visualize MS profiles similarity and distances, hierarchical clustering of the mass spectra of all tested species was performed using the dendrogram function of MALDI Biotyper, version 3.0. Dendrograms are based on the results of Composite Correlation Index (CCI) matrix. CCIs are calculated by dividing spectra into intervals and comparing these intervals across a dataset. The composition of correlations of all intervals provides the CCI, which is used as a parameter that defines the distance between spectra. A CCI match value of 1 represents complete correlation, whereas a CCI match value of 0 represents an absence of correlation (Laroche et al. Reference Laroche, Almeras, Pecchi, Bechah, Raoult, Viola and Parola2017).
DNA extraction
Two triatomines from each of the eight species included in this study were cut lengthwise in two equal halves, using a sterile surgical blade for each specimen. The remaining body parts were maintained at −20 °C as a backup sample. Each half triatomine was crushed with a pestle in 400 µL of a Qiagen G2 buffer (lysis buffer used in combination of proteinase K) solution containing 40 µ m of proteinase K (Qiagen, Hilden, Germany) and incubated at 56 °C overnight. After 1 min of centrifuging at 7168 RCF, 200 µL of the supernatant was then collected prior to DNA extraction. Triatominae genomic DNA was individually extracted using the EZ1 DNA tissue extraction kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Triatominae DNAs were then eluted in 100 µL of Tris EDTA buffer using the DNA extracting EZ1 Advanced XL Robot (Qiagen, Hilden, Germany) as previously described. The DNA was either immediately used or stored at −20 °C until molecular analysis. The DNA extracting EZI Advanced XL Robot was disinfected after each batch of extraction as per the manufacturer's recommendations, to avoid cross-contamination.
Sequencing and GenBank accession numbers
DNA samples were submitted to 16S gene conventional PCR amplification using a Bio-Rad Thermocycler (Bio-Rad Laboratories, Hercules, California, USA) prior to sequencing. Previously published primers amplifying a 520 bp region of the 16S gene were designed for this study (forward: 5′-CGCCTGTTTATCAAAAACAT-3′; reverse: 5′-CTCCGGTTTGAACTCAGATCA-3′) (Weirauch and Munro, Reference Weirauch and Munro2009). A mixture without DNA served as negative control. The cycling protocol consisted of 15 min at 95 °C followed by 35 cycles of denaturing at 95 °C for 30 s, annealing at 52 °C for 30 s, extension of 1 min at 72 °C, followed by a final cycle of 1 min at 72 °C and sampling while held at 4 °C. Amplification products were visualized by electrophoresis through a 1·5% agarose-tris-borate-EDTA gel containing ethidium bromide. PCR products were sequenced in both directions using a Big Dye Terminator kit and an ABI PRISM 3130 Genetic Analyzer (Applied BioSystems, Courtabeauf, France). The sequences were analysed using the ABI PRISM DNA Sequencing Analysis software version 3.0 (Applied BioSystems) and compared with sequences available in the GenBank database using the BLAST algorithm (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
RESULTS
Triatomines
A total of 149 triatomine specimens belonging to eight different species were included in this study. All 60 fresh specimens were T. infestans (n = 27) and R. prolixus (n = 33) specimens reared in our laboratory. Thirteen dry specimens of each of these two species were included in the study to assess MS reproducibility between fresh and dry specimens. All other specimens were dry (n = 63), and belonged to six other species, R. pictipes (n = 12), R. robustus (n = 12), P. geniculatus (n = 24), P. lignarius (n = 2), P. rufotuberculatus (n = 3) and E. mucronatus (n = 10), all collected in French Guiana.
Study evaluation on fresh samples
The femur-derived MS profiles of fresh T. infestans and R. prolixus were highly similar between adult specimens of the same species and different between the two species. All spectra displayed strong peak intensity, a mass range of 2–16 kDa and a number of peaks ranging from 110 to 143 peaks per spectrum.
As shown in Fig. 2, spectra for fresh specimens were highly reproducible within the same species. However, variations were observed depending on the developmental stage for both species, but also for sex in the case of T. infestans. As shown on Fig. 3, for both species, immature instars displayed the same MS profile, which was different from the spectrum of the adults of the same species. The database was consequently upgraded with random spectra from the different categories observed: male and female T. infestans, immature T. infestans, adult R. prolixus and immature R. prolixus.

Fig. 2. Representative MS spectra of Triatoma infestans and Rhodnius prolixus specimens displaying high reproducibility of the spectra within each group. Spectra analysis was performed using Flex analysis 3.3 software. Abbreviations: a.u., arbitrary units; m/z, mass-to-charge ratio.

Fig. 3. Representative MS spectra of adult and immature Triatoma infestans and Rhodnius prolixus specimens displaying variability according to developmental stage. Spectra analysis was performed using Flex analysis 3.3 software. Abbreviations: a.u., arbitrary units; m/z, mass-to-charge ratio.
Study validation and application to dry specimens using blind test analyses
Regarding fresh T. infestans and R. prolixus specimens, database query resulted in perfect identification of all species groups to the species, stage and sex, with LSVs ranging from 1·879 to 2·841 and 1·797 to 2·840, respectively (Table 1). All (100%) of the specimens were perfectly identified with LSVs ⩾2 for the highest spot.
Table 1. Arthropods used to establish the reference database of MALDI-TOF spectra and arthropods used in the blind test

a Calculation of score range includes LSVs of the four spots of each specimen. Blind test was not done on species whose number of specimens was too low.
M, male; F, female; N, nymph stage.
Spectra from dry T. infestans and R. prolixus specimens were subjected to MALDI-TOF analysis. Samples were correctly identified to the species level but with insufficient LSVs. The database was therefore upgraded with quality spectra from dry T. infestans and R. prolixus specimens. The consequent database interrogation resulted in the perfect identification of all specimens with LSVs ⩾1·8 for the highest spot. LSVs ranged from 1·707 to 2·861 for dry T. infestans specimens and from 1·741 to 2·236 for dry R. prolixus specimens.
All dry specimens from other species were then subjected to MALDI-TOF analysis and provided spectra of good intensity and good reproducibility within each group (Fig. 4). All spectra were subjected to blind test analysis. No correct identification was obtained before upgrading the database with random spectra from dry specimens. A dendrogram was realized and illustrated the segregation of different species based on their MS profile (Fig. 5). As shown in Table 1, 100% of the tested dry specimens were correctly identified by MALDI-TOF MS after a comprehensive database upgrade, with lower score for E. mucronatus specimens.

Fig. 4. Representative MS spectra of dry specimens from Triatoma infestans, Rhodnius prolixus, Rhodnius robustus, Rhodnius pictipes, Panstrongylus geniculatus, Panstrongylus lignarius, Panstrongylus rufotuberculatus, Eratyrus mucronatus species. Spectra analysis was performed using Flex analysis 3.3 software. Abbreviations: a.u., arbitrary units; m/z, mass-to-charge ratio.
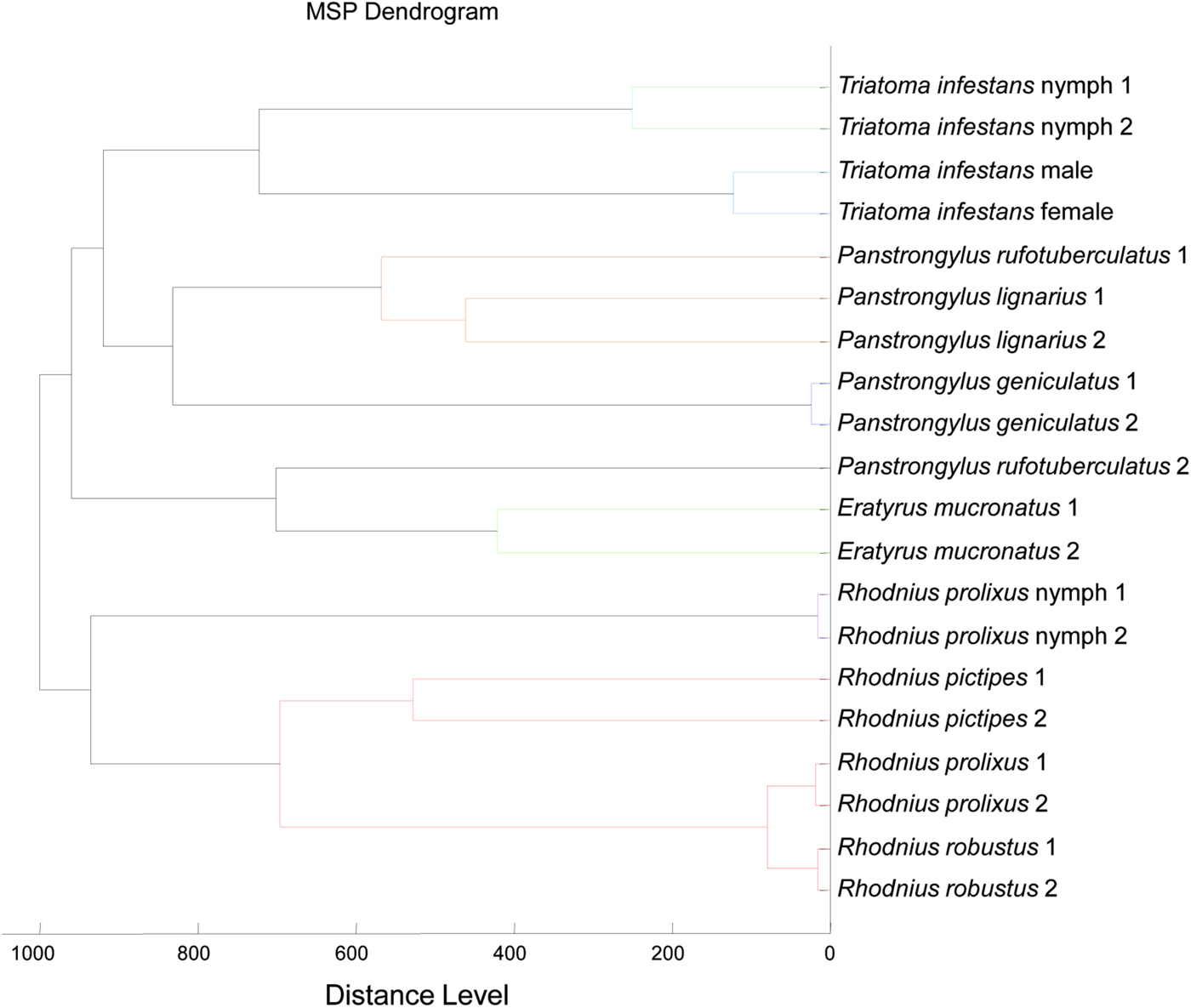
Fig. 5. Dendrogram obtained by cluster analysis of spectra obtained from two specimens of each triatomines species of the database. Clustering analysis was performed using MALDI Biotyper software. M, male; F, female; N, nymph.
Molecular analysis
The 16S gene sequences were obtained for all triatomine specimens subjected to sequencing. As shown in Table 2, T. infestans, R. prolixus, R. robustus, R. pictipes, P. geniculatus and E. mucronatus specimens displayed more than 99% identity with published GenBank sequences. No P. lignarius and P. rufotuberculatus 16S sequences were available in GenBank. New P. lignarius and P. rufotubercularus 520 bp partial 16S sequences are available in GenBank at #KY748238 and #KY748238, respectively.
Table 2. Identification of triatomines based on morphology, MALDI-TOF MS, and molecular biology approaches

Molecular identification is based on the sequencing of 520 bp fragment of the 16S gene. All new sequences were submitted to GenBank and the accession number is indicated.
DISCUSSION
Triatomine identification is currently principally morphologic (Monteiro et al. Reference Monteiro, Escalante and Beard2001). Some dichotomy keys exist, but their use is limited since they are often written in Spanish or Portuguese, and their access is limited. Without expertise in Reduviidae identification, Triatominae can easily be misidentified (Monteiro et al. Reference Monteiro, Escalante and Beard2001). As a matter of fact, Reduviidae is a large family in which only a small subfamily is haematophagous. Other species are mostly entomophagous (Schuh and Slater, Reference Schuh and Slater1995). Moreover, among Triatominae, vector competence regarding transmission of T. cruzi can be very variable, even between very close species (Pavan and Monteiro, Reference Pavan and Monteiro2007). The 16S gene sequencing was performed to validate unambiguously the identification of our specimens. The new sequences were submitted to GenBank.
In this work, to obtain triatomine-specific spectra, legs were selected for MALDI-TOF analysis. Indeed, as they are not part of the digestive tract, leg-derived spectra will not display any proteins from ingested blood. The same strategy was applied to other arthropod families (Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016). Ticks (Yssouf et al. Reference Yssouf, Flaudrops, Drali, Kernif, Socolovschi, Berenger, Raoult and Parola2013a) and mosquitoes (Yssouf et al. Reference Yssouf, Socolovschi, Flaudrops, Ndiath, Sougoufara, Dehecq, Lacour, Berenger, Sokhna, Raoult and Parola2013b) were identified based on leg-derived protein extract, while Culicoides (Sambou et al. Reference Sambou, Aubadie-Ladrix, Fenollar, Fall, Bassene, Almeras, Sambe-Ba, Perrot, Chatellier, Faye, Parola, Wade, Raoult and Mediannikov2015) and fleas (Yssouf et al. Reference Yssouf, Socolovschi, Leulmi, Kernif, Bitam, Audoly, Almeras, Raoult and Parola2014b) without the abdomen were selected for MS analyses. Preliminary work showed that leg profiles from a single triatomine specimen are identical no matter which leg is chosen. However, because we planned to work on dry, fragile specimens, we chose the median leg, which is the most rarely damaged. Whole leg protein extraction did not provide satisfying results, since they displayed heterogeneity within different specimens of the same species. This strategy was refined by choosing a smaller quantity of biological sample. Further experiments were performed on the femur of the median leg alone, which provided high-quality spectra that were highly reproducible within the same species. For all tested species except E. mucronatus, all specimens displayed adequate spectra quality for further analysis. However, for E. mucronatus, 4/10 specimens provided poor quality spectra. MALDI-TOF analysis requires several steps that may influence spectrum quality. The first parameter is the quality and knowledge of the sample's composition. The presence of potential entomopathogenic microorganisms may modify the host spectrum. However, as illustrated by previous studies, infection with arthropod-borne pathogens generally changes the spectrum without altering its quality. Moreover, it still allows a precise identification of the arthropod vector (Fotso Fotso et al. Reference Fotso Fotso, Mediannikov, Diatta, Almeras, Flaudrops, Parola and Drancourt2014; Yssouf et al. Reference Yssouf, Almeras, Berenger, Laroche, Raoult and Parola2015a, Reference Yssouf, Almeras, Terras, Socolovschi, Raoult and Parolab). Conservation mode is one of the most important parameters influencing spectral quality. Indeed, previous studies have described significant profile modifications, depending on whether samples were fresh or preserved in alcohol (Kumsa et al. Reference Kumsa, Laroche, Almeras, Mediannikov, Raoult and Parola2016). We also showed in the present work that fresh and dry samples displayed different MALDI-TOF spectra. However, E. mucronatus samples were preserved in the same conditions as the other species, so the conservation does not appear to be the reason for the poor quality spectra. Manipulator influence in the former crushing method with manual pestles may impair spectral quality (Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016). However, E. mucronatus samples were automatically crushed by a Tissue Lyser device, with the same programme and reagents as the other species. The quality of the CHCA matrix also has a major influence on the spectrum's quality (Yssouf et al. Reference Yssouf, Almeras, Raoult and Parola2016), but the same matrix was used to spot all samples. However, more samples are needed to conclude the failure of the experiment for this species.
Regarding the other Triatominae species, all tested specimens provided spectra of quality. This allowed for the construction of a reference database and the perfect identification of remaining specimens. The MS dendrogram realized with the database specimens displays a clear distinction for each species. However, the wrong position of a P. rufotuberculatus in the dendrogram illustrates that this tool is not adequate for phylogeny, as it was pointed out in previous studies (Yssouf et al. Reference Yssouf, Flaudrops, Drali, Kernif, Socolovschi, Berenger, Raoult and Parola2013a). The P. rufotuberculatus specimens provided the spectra with the lowest intensity of the whole study. The low intensity could be a reason for the incorrect classification of these specimens in the dendrogram. Nevertheless, the performance of the MS identification for triatomines is illustrated by the clear distinction of Rhodnius species, on the dendrogram but also in blind test analyses. Indeed, Rhodnius species have a very similar morphology and their distinction may be challenging (Pavan and Monteiro, Reference Pavan and Monteiro2007). Rhodnius prolixus and R. robustus, two close species, are one major example of the difficulty discriminating between Rhodnius species. Moreover, while R. prolixus is one of the most important vectors of T. cruzi to humans, species belonging to the R. robustus cryptic species complex comprise secondary vectors. As these species live in sympatry, accurate identification of each species is critical for epidemiological surveillance (Pavan and Monteiro, Reference Pavan and Monteiro2007). Specimens of these two species were subjected to MALDI-TOF analysis in the present study, and resulted in 100% specificity of the identification to the species level. If the MS database is comprehensive, unambiguous identification of these two species can be obtained without any entomological knowledge. These preliminary data are encouraging for further distinction of triatomine species belonging to the same complex such as the Triatoma complex (Pita et al. Reference Pita, Lorite, Nattero, Galvão, Alevi, Teves, Azeredo-Oliveira and Panzera2016).
Database upgrades will be needed in order to include other reference spectra from fresh or dry triatomine specimens. It would also be of interest to assess the consistency of MS spectra from diverse triatomine populations, since our fresh specimens were only from laboratory colonies. The performance of the identification could finally be challenged with identification of species belonging to the same complex. This technology has also been challenged by the concomitant identification of arthropod vectors and their associated pathogens. MALDI-TOF MS then appeared to be a promising tool for the direct detection of arthropod-borne pathogens, as was demonstrated with Borrelia (Fotso Fotso et al. Reference Fotso Fotso, Mediannikov, Diatta, Almeras, Flaudrops, Parola and Drancourt2014) and Rickettsia (Yssouf et al. Reference Yssouf, Almeras, Berenger, Laroche, Raoult and Parola2015a,Reference Yssouf, Almeras, Terras, Socolovschi, Raoult and Parolab) in ticks. It allowed the distinction of infected and uninfected ticks, based on the protein extracts from tick legs (Yssouf et al. Reference Yssouf, Almeras, Terras, Socolovschi, Raoult and Parola2015b) or haemolymph (Yssouf et al. Reference Yssouf, Almeras, Berenger, Laroche, Raoult and Parola2015a). The presence of parasites was detected as well in arthropods by MALDI-TOF MS since this technology was able to distinguish Anopheles mosquitoes infected or not by Plasmodium berghei (Laroche et al. Reference Laroche, Almeras, Pecchi, Bechah, Raoult, Viola and Parola2017). From these previous studies, it can be hypothesized that it would be of interest to assess whether this approach might be applicable to the detection of T. cruzi in triatomines. Until recently, it was accepted that the infection with T. cruzi did not have significant effect on the triatomine host (Elliot et al. Reference Elliot, Adler and Sabelis2003). However, recent studies described a significant impact on the arthropod's biology, particularly its reproduction (Fellet et al. Reference Fellet, Lorenzo, Elliot, Carrasco and Guarneri2014), locomotion (Marliére et al. Reference Marliére, Latorre-Estivalis, Lorenzo, Carrasco, Alves-Silva, de Rodrigues, de Ferreira, de Lara, Lowenberger and Guarneri2015), molting and survival (Elliot et al. Reference Elliot, de Rodrigues, Lorenzo, Martins-Filho and Guarneri2015). Such modifications might be associated with MS detectable proteome modifications.
If this infection is detectable with MALDI-TOF MS, this technique would be a rapid tool for the monitoring of the triatomine population T. cruzi endemic areas. This tool will allow the analysis of large populations of triatomines at low costs, and therefore identify human populations exposed to T. cruzi vectors.
Concluding remarks
MALDI-TOF appears to be a relevant tool for the precise identification of triatomine vectors. This method is independent from molecular biology or any knowledge of entomology, once the database is rendered comprehensive. The comprehensiveness of the database is the main limitation of this approach, which will always require unambiguous identification, first provided by morphology and molecular biology. The low cost and rapidity of this technique makes it an appropriate approach for epidemiological studies and the monitoring of triatomine populations in T. cruzi-endemic developing countries. In these countries, where vector control is the most conceivable approach to fight T. cruzi transmission, the rapid identification of exposed populations can provide a valuable advantage.
ACKNOWLEDGEMENTS
We thank the SEAG (Société Entomologique Antilles-Guyane) for kindly providing several triatomine specimens. We also thank Claudio Lazzari and Frédéric Lardeux for providing the Rhodnius prolixus and Triatoma infestans triatomines we are currently rearing in our insectary.
FINANCIAL SUPPORT
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
CONFLICT OF INTEREST
The authors declare that they have no conflicts of interest.
AUTHORS’ CONTRIBUTIONS
PP and ML designed and developed the experiments. ML, GG, J-MB and DB performed the experiments. ML, and PP analysed the data. ML and PP wrote the paper. DR and PP reviewed the manuscript. All authors reviewed and approved the final version.







