



Introduction
Giardia duodenalis (syns G. lamblia, G. intestinalis) is one of the most common enteric parasites worldwide with a diverse range of hosts that include humans, wildlife, domestic and companion animals (Feng et al., Reference Feng and Xiao2011). Giardia duodenalis is the only species found in humans and being increasingly recognized to cause gastrointestinal infection ranging from asymptomatic to severe diarrhoea as well as chronic diseases and malabsorption (Halliez et al., Reference Halliez and Buret2013). Giardia is mainly transmitted through the ingestion of cysts in contaminated surface food or water and direct contact with infected individuals, or directly via the fecal/oral route (Cacciò et al., Reference Cacciò, Beck, Lalle, Marinculic and Pozio2008). Giardia duodenalis is now considered to be divided into at least eight distinct genetic assemblages (A to H), of which assemblages A and B have the broadest host ranges and are mainly found to infect humans and other various mammals. The remaining assemblages (C to H) have strong host specificities and narrow host ranges. Assemblages C and D, for example, have been found in domestic dogs and other canines. Assemblage E has been found largely in domestic ruminants and pigs. Assemblages F and G are specific to cats and livestock, respectively, whereas assemblage H is usually found in marine vertebrates (Feng et al., Reference Feng and Xiao2011; Ryan et al., Reference Ryan and Cacciò2013).
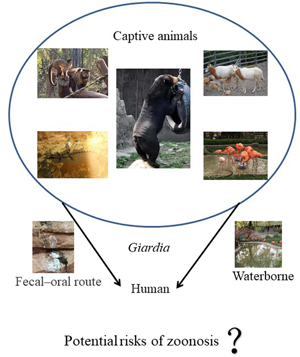
In recent years, the number of people visiting zoos has increased and people also like to be in contact with animals with the new construction of wildlife parks. Thus, the risk of potential zoonosis will increase and the role of captive animals, particularly in big cities, should be taken into consideration. Moreover, a growing number of emerging wildlife infectious diseases have posed a risk to human health and the investigations of wildlife diseases have been recognized as an important part of global health (Daszak et al., Reference Daszak, Cunningham and Hyatt2000). Giardia have been reported in captive animals such as non-human primates (NHPs) and wild birds worldwide, and zoonotic assemblages A and B of G. duodenalis have been found, which indicates the importance of the study of G. duodenalis in wild animals (Oates et al., Reference Oates, Miller, Hardin, Conrad, Melli, Jessup, Dominik, Roug, Tinker and Miller2012; Reboredo et al., Reference Reboredo-Fernández, Ares-Mazás, Cacciò and Gómez-Couso2015; Mynářová et al., Reference Mynářová, Foitová, Kváč, Květoňová, Rost, Morrogh-Bernard, Nurcahyo, Nguyen, Supriyadi and Sak2016). Animal husbandry is commonplace in China and the role of livestock and pets in the transmission of Giardia has been studied extensively (Liu et al., Reference Liu, Yang, Shen, Zhang, Wang, Zhao, Zhang, Ling and Cao2014, Reference Liu, Shen, Liu, Yin, Yuan, Jiang, Pan, Zhang, Zhao and Cao2017; Qi et al., Reference Qi, Wang, Jing, Wang, Jian, Ning and Zhang2016). However, very little information is available on the prevalence of G. duodenalis in the captive animals in China, and the assemblage distribution and the multilocus genotyping (MLG) of G. duodenalis remain unclear.
Therefore, the study was conducted to assess the prevalence and genetic characteristic description of zoonotic G. duodenalis in animals housed within Shanghai Wild Animal Zoo. The fecal samples from different animals were collected and examined the presence of G. duodenalis using nested polymerase chain reaction (nested PCR). In addition, the G. duodenalis genotypes were elucidated based on the common target genes: glutamate dehydrogenase (gdh), triose phosphate isomerase (tpi) and β-giardin (bg) (Feng et al., Reference Feng and Xiao2011). Then the potential for zoonotic transmission of G. duodenalis in these animals was assessed at the three loci.
Methods
Study area and sample collection
A total of 678 fecal specimens were collected from captive animals in a zoo in Shanghai, including NHPs (n = 190), herbivores (n = 190), carnivores (n = 151), birds (n = 138) and reptiles (n = 9). Each fresh fecal specimen was obtained using a disposable glove and plastic container. All the specimens were transported to NHC Key Laboratory of Parasite and Vector Biology of NIPD in a cooler with ice packs within 24 h of collection and processed as previously described (Fayer et al., Reference Fayer, Trout, Graczyk and Lewis2000) and were maintained at 4°C for no more than 3 days before DNA extraction.
DNA extraction and PCR amplification
Genomic DNA was extracted from all fecal samples using the QIAamp DNA Stool Mini Kit (QIAGEN, Valencia, USA) following the manufacturer's instructions. Purified DNA samples (200 μL) were stored at −20°C for further molecular analysis. To determine G. duodenalis subtypes, gdh, tpi and bg genes were simultaneously amplified and the PCR amplification parameters have been previously described (Sulaiman et al., Reference Sulaiman, Fayer, Bern, Gilman, Trout, Schantz, Das, Lal and Xiao2003; Lalle et al., Reference Lalle, Pozio, Capelli, Bruschi, Crotti and Cacciò2005; Cacciò et al., Reference Cacciò, Beck, Lalle, Marinculic and Pozio2008). In all the PCR reactions, a dog-derived Giardia-positive DNA specimen (assemblage C) and distilled water were used as positive and negative controls, respectively. The second PCR products were analysed using 1.5% agarose gel stained with ethidium bromide.
Sequence and genotype analysis
All target fragments were amplified at least three times and all the PCR-positive products were sequenced in both directions using the secondary primers on an ABI 3730 DNA Analyzer (Applied Biosystems, Foster City, USA) using a Big Dye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems) (Long et al., Reference Long, Cai and He2018). Nucleotide sequences obtained in this study were aligned, examined and compared with reference sequences downloaded from the GenBank database using Clustal X 1.83 and Basic Local Alignment Search Tool (BLAST).
Accession numbers of the nucleotide sequences
The new sequences generated in this study have been deposited in the GenBank under accession numbers MT811038–MT811045.
Results
Prevalence of G. duodenalis by PCR
Of the 678 fecal samples examined, 42 (6.2%) were tested G. duodenalis-positive at least based on a single locus. A total of 30 (4.4%), 30 (4.4%) and 22 (3.2%) specimens were successfully amplified and sequenced at gdh, tpi and bg loci, respectively. MLG data at the three loci tested were available for 14 samples and no mixed infections were found. In addition, 12 and 16 G. duodenalis isolates were successfully amplified at two and one loci, respectively.
Different animals have different infection rates; NHPs have the highest infection rate reaching 17.4% (33/190). Of the 33 positive isolates, 14 Lemur catta, 8 Pantroglodytes, 7 Lemur variegatus, 2 Rhinopithecus and 2 Saimiri sciureus were obtained for G. duodenalis. Following by, the infection rate of carnivores was 2.8% (4/142), with four Helarctos malayanus being identified. The infection rate of birds was 1.4% (2/138) including one Dromaius novaehollandia and one Acridotheres cristatellus, and 1.1% (2/190) of herbivore animals with one Oryx and one Damaliscus dorcas being identified. One Myrmecophaga tridactyla was also being identified to be Giardia-positive. No giardia infection was found in reptiles (Table 1).
Table 1. Infection rates based on gdh, tpi and bg loci used by nested PCR

Molecular genotyping at the bg gene
Amplification of a 510 fragment of the bg gene was obtained from 22 of 42 (52.38%) G. duodenalis isolates. At bg gene, 19 sequences were identified as assemblage B and three were assemblage A. In addition, sequence analysis revealed one (SHWD185) and three (SHWD159, SHWD168 and SHWD357) isolates and have 100% similarity with the human-derived isolates MG746614 and MG736242, respectively. A new isolate (GenBank no. MT811038) was identified at bg locus with one base mutation (A→G) with the reference sequence KJ88974 in the GenBank.
Molecular genotyping at the gdh gene
At 30 gdh-positive specimens, assemblages A and B were found in 3 (10%) and 27 (90%) specimens, respectively, with assemblage B being the major genotypes (Table 2). Two new sequences were identified in NHPs, one sequence (GenBank no. MT811041) has two base differences (G→A, G→A) and the other sequence (MT811040) also has two base mutations (C→T, C→T) compared with the deposited sequence in the GenBank (KM977648) (see Supplementary Table S2).
Table 2. Characterization of Giardia duodenalis based on multi-loci of gdh, tpi and bg genes

Molecular genotyping at the tpi gene
Based on analysis targeting tpi gene, assemblages A and B were found in 6 (20%) and 24 (80%) specimens, respectively, with assemblage B also being the major genotype. The sequence alignment indicated that six isolates (SHWD12, SHWD25, SHWD363, SHWD 365, SHWD366 and SHWD160) were identical to the human-derived isolates (KT948104). Five new assemblage subtypes were identified and the single-nucleotide polymorphisms (SNPs) are shown in the Supplementary Materials (Table S2). Using MF095053 as a reference sequence, isolate MT811043 has two SNPs at nucleotide positions 175 (A→G) and 522 (T→G); isolate MT811045 has two SNPs at 77 (T→C) and 175 (A→G); isolate MT811041 has two SNPs at 153 (C→T) and 522 (T→G); isolate MT811045 has one SNP at 250 (G→A) and isolate MT811045 has two SNPs at 250 (G→A) and 382 (G→A).
Discussion
Giardia duodenalis is one of the most common intestinal parasites resulting in waterborne and foodborne diarrhoea; it is also an important cause of traveller's diarrhoea. With the increasing reports on this parasitic outbreak, more and more investigations on this parasite in captive animals have been reported worldwide. The aim of this study was to investigate the prevalence and the zoonotic potential of Giardia based on genotypes by amplifying different loci in various captive animal species in a zoo in Shanghai, China.
In this study, the overall infection rate of G. duodenalis was 42 (6.2%) in 678 fecal samples examined from captive animals in zoological gardens. Zoonotic assemblages A (n = 6) and B (n = 36) were found, with assemblage B being more prevalent and in agreement with a previous study (Zhong et al., Reference Zhong, Tian, Li, Huang, Deng, Cao, Geng, Fu, Shen, Liu and Peng2017). In these animals involved in this study, the infection rate of NHPs was the highest (17.4%, 33/190). Giardia duodenalis infections in NHPs have been reported globally and the overall infection rate varied from 0 to 40% of examined G. duodenalis infection in NHPs in parks, zoos, farms or laboratories in China.
In the current study, the infection rate of NHPs was 17.4% and close to the prevalence in Dalian zoo (16.7%) (Zhang et al., Reference Zhang, Wang, Lan, Dan, Ren, Cao, Shen, Deng, Zuo, Yu, Wang, Ma, Liu, Zhou, Hu, Fu, He, Geng, Gu, Peng, Wang and Zhong2020). It was higher than those reported in Qianling Park in Guiyang (8.5%) (Ye et al., Reference Ye, Xiao, Ma, Guo, Liu and Feng2012), Sichuan and Guizhou provinces (7.7%) (Zhong et al., Reference Zhong, Tian, Li, Huang, Deng, Cao, Geng, Fu, Shen, Liu and Peng2017), Qinling Mountain (2.0%) (Du et al., Reference Du, Zhao, Shao, Fang, Tian, Zhang, Wang, Wang, Qi and Yu2015) and two other additional comprehensive parasite infection studies in China (2.2 and 1.3%) (Ye et al., Reference Ye, Xiao, Ma, Guo, Liu and Feng2012; Li et al., Reference Li, Dong, Wang, Yu, Wu, Chang, Wang, Qi and Zhang2017) Table 3. We can find that different areas have different G. duodenalis infection rates in NHPs in China due to a range of factors, including the diagnostic method and study design, geographical conditions, number of samples and sampling season. Our results and those of previous studies indicate that G. duodenalis infection is common in NHPs and has a wide geographic distribution in China.
In other countries in the world, the overall infection rate of G. duodenalis in NHPs is between 2.2 and 47.0% (Berrilli et al., Reference Berrilli, Prisco, Friedrich, Cerbo, Cave and Liberato2011; Karim et al., Reference Karim, Zhang, Jian, Li, Zhou, Zhang, Sun, Yang, Zou, Dong, Li, Rume, Qi, Wang, Ning and Xiao2014). Reports of the prevalence rate of giardiasis in NHPs from different countries varies from 6.0% in Italy (Berrilli et al., Reference Berrilli, Prisco, Friedrich, Cerbo, Cave and Liberato2011) and 7.0% in Thailand (Sricharern et al., Reference Sricharern, Inpankaew, Keawmongkol, Supanam, Stich and Jittapalapong2016), and it was lower than 11.1% in Uganda (Johnston et al., Reference Johnston, Gillespie, Rwego, McLachlan, Kent and Goldberg2010) and 50% in Croatia (Beck et al., Reference Beck, Sprong, Bata, Lucinger, Pozio and Cacciò2011). The different infection rates may be explained by diagnostic methods, the climate among these regions, environmental management and NHP species. We also observed that the high infection rate of NHPs could be attributed to the frequent contact within the range of activities. It can actually increase the risk of G. duodenalis transmission.
Based on MLG, the MLG model was used to better understand the infection and genetic characteristics of G. duodenalis in animals and humans, which is helpful for unveiling zoonotic potential and dynamic transmission. In our study, the genetic diversity of these positive G. duodenalis isolates was determined by the amplification and sequencing of the gdh, tpi and bg genes, with 30 gdh, 30 tpi and 22 bg gene sequences being obtained (Table 2). gdh and tpi genes gave a higher PCR amplification rate. Molecular characterization of G. duodenalis isolates revealed that all the infected NHPs were zoonotic assemblage B, which is consistent with previous reports (Johnston et al., Reference Johnston, Gillespie, Rwego, McLachlan, Kent and Goldberg2010; Ryan et al., Reference Ryan and Cacciò2013; Karim et al., Reference Karim, Wang, Yu, Li, Dong, Li, Zhang, Li, Jian, Zhang, Rume, Ning and Xiao2015). At bg locus, one isolate (SHWD185) and three isolates (SHWD159, SHWD168 and SHWD357) were identical to the human-derived isolates (MG746614 and MG736242). At gdh locus, one isolate (SHWD367) showed complete homologous to the reference sequence of the human-derived isolate (MH311013). Six isolates have 100% similarity with human-derived isolates (GenBank no. KT948104) based on the tpi gene (Wegayehu et al., Reference Wegayehu, Karim, Li, Adamu, Erko, Zhang and Tilahun2016). The sequence data indicated these animals could be potentially infectious to other animals or surroundings, and further study is essential to clarify the transmission. In addition, we also observed a high genetic diversity in assemblage B subtypes at tpi locus.
Table 3. Occurrence and assemblages of G. duodenalis for NHPs from different locations in China

Giardia duodenalis assemblages A and B are zoonotic and can infect a broad range of hosts including humans. However, fecal–oral route is the primary way of G. duodenalis transmission. Ingestion of infective cysts through contaminated water and food is the common transmission route. To the best of our knowledge, during the past decade, more and more studies have showed that the close correlation between human activity and the occurrence of wildlife diseases (Thompson et al., Reference Thompson, Lymbery and Smith2010; Brearley et al., Reference Brearley, Rhodes, Bradley, Baxter, Seabrook, Lunney, Liu and McAlpine2013). In this zoo, different kinds of animals such as tigers, lions or birds lived in a relatively independent environment. However, they could be indirectly in contact with humans through the surrounding water. In addition, breeders and visitors have great chance to get in touch with some kinds of caged birds and NHPs. Previous studies have reported that the most Giardia infections naturally occurring in wildlife may be related to the wildlife habitats, and influenced by direct contact or environmental routes (Thompson, Reference Thompson2013; Abeywardena et al., Reference Abeywardena, Jex and Gasser2015). In fact, cross-species transmission among the same kinds of animals is possible, and cleaning up the feces in time can reduce the pollution to a great extent. However, different kinds of animals have little chance to be in contact with each other because they lived in a relatively independent environment. Therefore, the feeding and management strategies should be improved and the disease surveillance should also be strengthened to prevent Giardia infection. For example, we suggest that zoo manager can reduce the frequency of human–animal contact, increase the construction of zoo zoning and clean up the feces excreted by animals in time to reduce the pollution to the surrounding environment, animals or humans. In the future studies, the breeders and water sources around these animals, in addition to the other animals in contact with them should also be investigated for the occurrence of G. duodenalis and its genetic characteristics. Also, the effect of Giardia infection on the health of captive animals in zoological gardens needs to be resolved.
Conclusions
Our study revealed the presence of G. duodenalis in captive animals in zoological gardens and new isolates were identified in NHPs. Assemblages A and B were found with assemblage B being dominated. The sequences were 100% homologous with registered human-derived sequences in the GenBank indicating the potential possibility of zoonotic transmission. Nevertheless, to evaluate whether the surrounding environmental contamination caused by these animals or other anthropogenic activities, the detection of more specimens including water, other animals or human feces will be involved in further study.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182021000913
Acknowledgements
We thank the Shanghai Wild Animal Zoo for the permission to collect fecal samples.
Author contributions
YS, HL and JC conceived and designed the study. BW and ZY conducted sample collection. BW and HL performed laboratory experiments. YJ, JZ and JY performed statistical analyses. HL wrote the manuscript. YS revised the manuscript. All authors have read and approved the final version of the manuscript.
Financial support
This study was supported by The National Key Research and Development Program of China (2017YFD0500400), the National Science and Technology Major Project (Nos. 2018ZX10713001-004 to YS and 2018ZX10201002-009 to JY), the National Natural Science Foundation of China (No. 81772224 to YS) and the Fifth Round of Three-Year Public Health Action Plan of Shanghai, China (No. GWV-10.1-XK13 to JC). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Conflict of interest
None.
Ethical standards
Prior to collection of fecal specimens, permission was obtained from zoo managers. No specific permits were required for the described field studies. All fecal specimens were collected from the ground using plastic bags the following day. Animals were not harmed in any way during the procedure. This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention. The protocol was approved by the Laboratory Animal Welfare & Ethics Committee (LAWEC), National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention (No. 2014001).





