



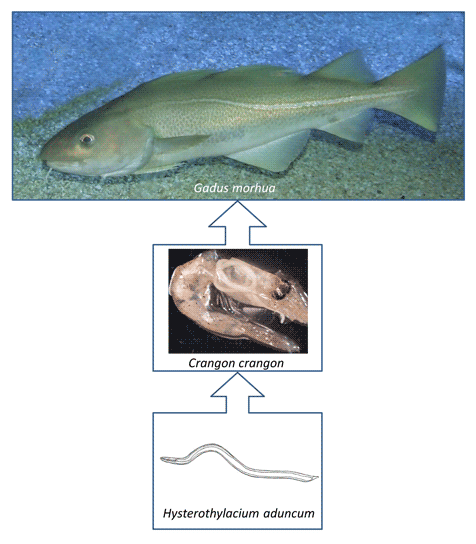
Introduction
Baltic cod (Gadus morhua) is one of the most commercially exploited fish species and is a popular food in several countries. Cod concentrates in deeper waters for spawning and migrates to the water column or shallow areas to feed (Bagge et al., Reference Bagge, Thurow, Steffensen and Bay1994). The dietary preferences of cod, which is a predatory fish throughout most of its life, depend on its ability to catch and eat specific prey species. Young cod mostly occur near the coast, in shallow water, and feed on invertebrates, especially Crustacea (e.g. Crangon crangon, Mysis mixta and Gammarus sp.). Older cod migrate to deeper, offshore waters where they prefer to eat fish (Clupea harengus and Sprattus sprattus) and larger invertebrates (Saduria entomon). Thus, the variability in the cod diet reflects the age of the fish and the biodiversity of prey species in the habitats occupied. Pachur and Horbowy (Reference Pachur and Horbowy2013) revealed that a shift in dietary composition can be observed as cod reach between 30 and 40 cm in length. This shift in the cod's diet has consequences for the risk of infection with different parasite species.
The ecological status of the Baltic Sea is deteriorating and as a consequence the state of the cod population and the condition of individual cod grow progressively worse. This might be due to low availability of fish prey in areas where cod feed (Eero et al., Reference Eero, Vinther, Haslob, Huwer, Casini, Storr-Paulsen and Koster2012); a lack of benthic prey because of lower dissolved oxygen or changes in salinity (Conley et al., Reference Conley, Bj̈orck, Bonsdorff, Carstensen, Destouni, Gustafsson, Hietanen, Kortekaas, Kuosa, Meier, Müller-Karulis, Nordberg, Norkko, Nürnberg, Pitkänen, Rabalais, Rosenberg, Savchuk, Slomp, Voss, Wulff and Zillén2009; Carstensen et al., Reference Carstensen, Andersen, Gustafsson and Conley2014); intensive exploitation of resources, which negatively influences fish stocks (Lindegren et al., Reference Lindegren, Möllmann, Nielsen and Stenseth2009) and increasing infection with nematode parasites (Haarder et al., Reference Haarder, Kania, Galatius and Buchmann2014; Mehrdana et al., Reference Mehrdana, Bahlool, Skov, Marana, Sindberg, Mundeling, Overgaard, Korbut, Strom, Kania and Buchmann2014; Horbowy et al., Reference Horbowy, Podolska and Nadolna-Ałtyn2016). The Baltic cod population has decreased drastically over the years, and the International Council for the Exploration of the Sea (ICES) has advised fisheries focused on cod in the Baltic Sea to cease fishing in 2020 (ICES, 2019).
Studies conducted on Gadiformes fish, specifically the burbot (Lota lota), by Valtonen and Julkunen (Reference Valtonen and Julkunen1995) revealed that the type of diet was a key factor determining the risk of infection with particular species of parasite. Cod can be intermediate, paratenic or definitive hosts for a large number of parasite species (Hemmingsen and MacKenzie, Reference Hemmingsen and MacKenzie2001), and the parasite fauna of cod differs depending on the size of individual fish (Zuo et al., Reference Zuo, Huwer, Bahlool, Al-Jubury, Christensen, Korbut, Kania and Buchmann2016). In the Baltic Sea, the dominant group of parasites in small cod is acanthocephalans, especially Echinorhynchus gadi, which occur in the digestive tract (Pilecka-Rapacz and Sobecka, Reference Pilecka-Rapacz and Sobecka2004). In larger cod, the most abundant parasites are nematodes, particularly Contracaecum osculatum (Szostakowska et al., Reference Szostakowska, Myjak, Wyszyński, Pietkiewicz and Rokicki2005; Nadolna and Podolska, Reference Nadolna and Podolska2014; Mehrdana et al., Reference Mehrdana, Bahlool, Skov, Marana, Sindberg, Mundeling, Overgaard, Korbut, Strom, Kania and Buchmann2014) and occasionally Anisakis sp. (Nadolna and Podolska, Reference Nadolna and Podolska2014), which occupy the liver; rarely, Pseudoterranova sp. and Anisakis sp. accumulate in the muscle tissue (Mehrdana et al., Reference Mehrdana, Bahlool, Skov, Marana, Sindberg, Mundeling, Overgaard, Korbut, Strom, Kania and Buchmann2014). Hysterothylacium sp. occurs mainly in the digestive tract of larger cod. In spite of the fact that the parasite fauna of cod from the Baltic Sea is well known and has been studied by several authors (Myjak et al., Reference Myjak, Szostakowska, Wojciechowski, Pietkiewicz and Rokicki1994; Buchmann, Reference Buchmann1995; Mellergaard and Lang, Reference Mellergaard and Lang1999; Perdiguero-Alonso et al., Reference Perdiguero-Alonso, Montero, Raga and Kostadinova2008; Haarder et al., Reference Haarder, Kania, Galatius and Buchmann2014; Mehrdana et al., Reference Mehrdana, Bahlool, Skov, Marana, Sindberg, Mundeling, Overgaard, Korbut, Strom, Kania and Buchmann2014; Nadolna and Podolska, Reference Nadolna and Podolska2014; Zuo et al., Reference Zuo, Huwer, Bahlool, Al-Jubury, Christensen, Korbut, Kania and Buchmann2016), the life cycles of Baltic cod parasites are described only in general.
Cod infection with the nematode parasite Hysterothylacium sp., as well as its basic life cycle, is documented for the Baltic Sea, but little is known about the crustacean species that play the role of intermediate host. The aim of this study was to determine the source of infection of Baltic cod with parasites by examination of invertebrates found in situ in cod stomach.
Materials and methods
The research material (cod stomachs) was collected during four research surveys in November (Q4) 2012 and 2013; as well as in February (Q1) 2013 and 2014. In total, 1681 cod stomachs were sampled in the Polish Exclusive Economic Zone, southern Baltic Sea. Ichthyological analysis of each individual cod was performed on board the survey ship. The stomachs were frozen at −20°C for further food content analysis in the laboratory on land. Analysis of the cod diet was performed: prey items found in stomachs were sorted and classified to the lowest possible taxonomic level (Żmudziński, Reference Żmudziński1990; Hayward and Ryland, Reference Hayward and Ryland1995). All invertebrates were collected, counted and analysed one by one for the presence of parasites. The organisms found had decomposed to different extents: some were partly digested, whereas others were not macerated at all. Well-preserved invertebrates were digested in the laboratory in artificial gastric juice (aqueous solution of pepsin and 35–38% HCl) to expose any parasites in the body cavity. This treatment increased the transparency of prey organisms and improved the level of detection of parasites. All parasites observed were collected and examined under the stereomicroscope for taxonomic identification on the basis of anatomo-morphological features (Fagerholm, Reference Fagerholm1982; Berland, Reference Berland1989). Infection with parasites is described according to the prevalence and intensity of infection (Bush et al., Reference Bush, Lafferty, Lotz and Shostak1997).
To confirm taxonomic identification, molecular analysis of parasites was performed. Analysis was conducted according to Zhu et al. (Reference Zhu, Gasser, Podolska and Chilton1998) and involved the amplification and sequencing of the ITS-1 (Internal Transcribed Spacer) region of rDNA. Polymerase chain reaction (PCR) products were sequenced directly using standard procedures and amplification primers. DNA was isolated using a Genomic Mini Kit (A&A Biotechnology, Gdynia, Poland) according to the manufacturer's instructions. Amplification was performed using the primers NC5 (forward) 5′-GTA GGT GAA CCT GCG GAA GGA TCA TT-3′ and NC13R (reverse) 5′-GCT GCG TTC TTC ATC GAT-3′. The reaction mixture consisted of 25 μL PCR Master MixPlus High GC (ready-to-use PCR mixture containing Taq DNA polymerase, PCR buffer, MgCl2 and dNTPs; A&A Biotechnology), 0.2 μL each primer (stock concentration 100 μ m), 20 μL DNA template and supplemented with deionized water up to 50 μL. The PCR reaction conditions were as follows: 2 min at 94°C (initial denaturation) followed by 35 cycles of 30 s at 94°C, 30 s at 60°C, 30 s at 72°C and a final extension step of 5 min at 72°C. Some DNA fragments obtained as a result of the amplification reaction were purified using a Gel-Out Concentrator Kit (A&A Biotechnology). PCR products were eluted with sterile water. Sequences were analysed using software CLC Workbench and GeneStudio and confirmed by a BLAST search of the GenBank.
Results
The food content analysis revealed 15 467 invertebrates in cod stomachs (Table 1). The dominant (the most numerous) species were Crustacea, especially C. crangon, S. entomon, M. mixta, Gammarus sp.; the polychaete Bylgides sarsi was also frequently observed. Parasitological analysis of the invertebrates found in cod stomachs revealed the presence of Hysterothylacium sp. L3 larvae, but only in the decapod C. crangon (Fig. 1). The first microscopic investigation of C. crangon was insufficient, and additional digestion in artificial gastric juice was necessary to determine the presence of nematodes in the body cavity. Among the 4731 C. crangon examined, parasites were found in nine individuals: five in the sample from November 2012; one from February 2013; two from November 2013 and one from February 2014 (Table 2). On the basis of anatomo-morphological features, all the above parasites were identified as Hysterothylacium sp. Therefore, the prevalence of C. crangon infection with Hysterothylacium sp. was 0.0027% in November 2012; 0.0005% in February 2013; 0.0067% in November 2013 and 0.0018% in February 2014. The mean prevalence of infection was 0.0029%. The intensity of infection was 1 in every case.

Fig. 1. Crangon crangon infected with Hysterothylacium aduncum (photo: J. Pawlak).
Table 1. Dominant invertebrate species among food items in cod stomachs (number of individuals)

Table 2. C. crangon infected with Hysterothylacium aduncum/Hysterothylacium sp. nematodes in stomachs of Baltic cod

Molecular analysis identified seven parasites as Hysterothylacium aduncum; two individuals were impossible to verify by DNA sequencing. The long process of obtaining parasites from under the carapace of crustaceans (digestion in stomach, freezing, additional digestion in artificial gastric juice) is likely to result in the partial degradation of the DNA in a significant proportion of cases and cause difficulties with molecular identification. Where parasites were successfully identified, the sequence similarity was 98.21–100% with H. aduncum compared to examples registered in the GenBank. Examples of the sequences obtained have been deposited in the GenBank (accession no. MW506285, MW506286, MW506287, MW506288 and MW506289).
Table 3 shows all parasites found in cod stomachs near to, but not within, food items (excluding samples from 2012, where they were not collected). The nematode Hysterothylacium sp. and the acanthocephalan E. gadi, as well as representatives of Trematoda, were found.
Table 3. Parasites found in stomachs of Baltic cod

Discussion
Several recent studies have revealed a remarkable increase in the prevalence of cod infection with Anisakidae nematodes (Haarder et al., Reference Haarder, Kania, Galatius and Buchmann2014; Mehrdana et al., Reference Mehrdana, Bahlool, Skov, Marana, Sindberg, Mundeling, Overgaard, Korbut, Strom, Kania and Buchmann2014; Nadolna and Podolska, Reference Nadolna and Podolska2014; Horbowy et al., Reference Horbowy, Podolska and Nadolna-Ałtyn2016; Zuo et al., Reference Zuo, Huwer, Bahlool, Al-Jubury, Christensen, Korbut, Kania and Buchmann2016). The negative effect of such an increase in the intensity of infection on the condition of fish has also been reported (Horbowy et al., Reference Horbowy, Podolska and Nadolna-Ałtyn2016). The parasite fauna of Baltic cod is well known (Myjak et al., Reference Myjak, Szostakowska, Wojciechowski, Pietkiewicz and Rokicki1994; Buchmann, Reference Buchmann1995; Mellergaard and Lang, Reference Mellergaard and Lang1999; Perdiguero-Alonso et al., Reference Perdiguero-Alonso, Montero, Raga and Kostadinova2008; Haarder et al., Reference Haarder, Kania, Galatius and Buchmann2014; Mehrdana et al., Reference Mehrdana, Bahlool, Skov, Marana, Sindberg, Mundeling, Overgaard, Korbut, Strom, Kania and Buchmann2014; Nadolna and Podolska, Reference Nadolna and Podolska2014; Zuo et al., Reference Zuo, Huwer, Bahlool, Al-Jubury, Christensen, Korbut, Kania and Buchmann2016). Invertebrates, which represent important basic food items for Baltic cod in their early stages of development, are thought to be the first intermediate host and transmitter of pathogenic nematodes to fish (Bagge et al., Reference Bagge, Thurow, Steffensen and Bay1994; Horbowy et al., Reference Horbowy, Podolska and Nadolna-Ałtyn2016; Engelhardt et al., Reference Engelhardt, Frisell, Gustavsson, Hansson, Sjoberg, Collier and Balk2020). The role of particular invertebrate species in the life cycles of specific parasites is not precisely defined. Clearly, a high-quality diet is essential for healthy fish development.
Similar to the research conducted by Pachur and Horbowy (Reference Pachur and Horbowy2013), in the current study, the dominant invertebrate food items in cod stomach were Malacostraca, especially C. crangon, S. entomon, M. mixta, Gammarus sp. and Polychaeta B. sarsi. The presence of the parasite in the brown shrimp suggests that this invertebrate is not only a source of nutrients, but might also be a route of infection with parasites. However, parasites were found only in the brown shrimp, C. crangon (Decapoda), which frequently occurs in offshore sandy and sandy-muddy habitats in the Baltic and North Seas, and also along the north and west coasts of Europe and the American coastal waters of the North Atlantic (Żmudziński, Reference Żmudziński1967). Brown shrimp is a migratory species: in the autumn, when temperatures decrease, it migrates into deeper waters, returning to shallower waters in the spring (Żmudziński, Reference Żmudziński1961). Therefore, it is accessible to both large demersal cod in the colder part of the year and to small cod individuals present in shallow waters. In the Polish waters of the southern Baltic Sea, C. crangon reaches 55 (male) to 70 (female) mm in size (Szaniawska, Reference Szaniawska1991), making it easily available even to small cod; consequently, it is a common item in the cod diet (Pachur and Horbowy, Reference Pachur and Horbowy2013). The role of C. crangon as a transmitter of the nematode parasite Anisakis simplex to Baltic cod has been described by Pawlak et al. (Reference Pawlak, Nadolna-Ałtyn, Szostakowska, Pachur, Bańkowska and Podolska2019). The results presented in this report are the first in situ confirmation that C. crangon may be a route for cod infection with H. aduncum.
In this study, C. crangon was found to be infected with L3-stage H. aduncum larvae. Genetic identification confirmed the results of anatomo-morphological analysis. To my best knowledge, this is the first evidence for this host–parasite system in the Baltic Sea. The prevalence of C. crangon infection with Hysterothylacium sp. was 0.0027% in November 2012; 0.0005% in February 2013; 0.0067% in November 2013 and 0.0018% in February 2014; the intensity of infection was 1 in all cases. Although the prevalence of infection is low, C. crangon is an important food item in the cod diet, and therefore this invertebrate may play a role as a transmitter of parasites to cod.
The nematode Hysterothylacium sp. (mainly H. aduncum), a member of the Raphidascarididae family, is a common parasite of marine fish throughout the world (Andersen, Reference Andersen1993; Rello et al., Reference Rello, Adroher and Valero2008; Knoff et al., Reference Knoff, Felizardo, Iñiguez, Maldonado, Torres, Pinto and Gomes2012; Moravec et al., Reference Moravec, Taraschewski, Appelhoff and Weyl2012; Kong et al., Reference Kong, Fan, Zhang, Akao, Dong, Lou, Ding, Tong, Zheng, Chen, Ohta and Lu2015; Morsy et al., Reference Morsy, Bashtar, Mostafa, El Deeb and Thabet2015; Shamsi et al., Reference Shamsi, Poupa and Justine2015; Shamsi et al., Reference Shamsi, Ghadam, Suthar, Mousavi, Soltani and Mirzargar2016; Shamsi, Reference Shamsi2017; Ghadam et al., Reference Ghadam, Banaii, Mohammed, Suthar and Shamsi2018; Roca-Geronès et al., Reference Roca-Geronès, Montoliu, Godínez-González, Fisa and Shamsi2018) and is the most frequently occurring parasite in invertebrates acting as intermediate hosts: it has been reported in 70 different invertebrate species (Lick, Reference Lick1991). Hysterothylacium aduncum (Rudolphi 1802) has a circumpolar distribution in the Northern hemisphere (Deardorff and Overstreet, Reference Deardorff and Overstreet1981) and has been found in the north-west Atlantic (Marcogliese, Reference Marcogliese1996), the North Sea and the Baltic Sea (Lick, Reference Lick1991; Klimpel and Ruckert, Reference Klimpel and Ruckert2005), but also in the Mediterranean Sea (Dural et al., Reference Dural, Genc, Sangun and Güner2011; Abdel-Ghaffar et al., Reference Abdel-Ghaffar, Abdel-Gaber, Bashtar, Morsy, Mehlhorn, Al Quraishy and Saleh2015), the Black Sea (Pekmezci et al., Reference Pekmezci, Bolukbas, Gurler and Onuk2013) and the waters around Japan (Moravec and Nagasawa, Reference Moravec and Nagasawa1986; Kong et al., Reference Kong, Fan, Zhang, Akao, Dong, Lou, Ding, Tong, Zheng, Chen, Ohta and Lu2015). In the southern hemisphere, it has been found among other locations in the south-west Atlantic (Navone et al., Reference Navone, Sardella and Timi1998), and around Australia (Shamsi et al., Reference Shamsi, Poupa and Justine2015, Reference Shamsi, Steller and Chen2018).
In general, in the H. aduncum life cycle, invertebrates play the role of intermediate hosts and fish are the final host. Natural infection with larval H. aduncum has been documented in seven phyla of both benthic and planktonic invertebrates in the north-western Atlantic (Norris and Overstreet, Reference Norris and Overstreet1976; Marcogliese, Reference Marcogliese1996). Third-stage larvae of H. aduncum have been obtained from C. crangon in the Ythan estuary (Scotland) (Gibson, Reference Gibson1972). In the Canadian Bras d'Or Lake, H. aduncum uses a variety of intermediate hosts including the mysids Neomysis americana and M. stenolepis and the chaetognath Sagitta elegans, as well as mixture of zooplankton: calanoid copepods, crab zoea and megalops and euphausiid larvae (Jackson et al., Reference Jackson, Marcogliese and Burt1997). Hysterothylacium sp. has been reported by Lick (Reference Lick1991) in several invertebrate species from the North Sea and Baltic Sea (studies limited to German coastal waters), including Acartia bifilosa, Eurytemora affinis, Temora longicornis, Pseudocalanus elongatus and M. mixta. Infection of gammarid species including Gammarus locusta, G. salinus and G. zaddachi with Hysterothylacium sp. in the same area has been noted by Lick (Reference Lick1991) and Zander et al. (Reference Zander, Reimer, Barz, Dietel and Strohbach2000). The same nematode parasite has also been recorded in representatives of Calanoida (Svendsen, Reference Svendsen1990; Marcogliese, Reference Marcogliese1995), especially in Hyperia galba and Idotea spp. from the North Sea (Klimpel and Ruckert, Reference Klimpel and Ruckert2005) and Neomysis. integer from both the North Sea and German coastal waters of the Baltic Sea (Lick, Reference Lick1991; Klimpel and Ruckert, Reference Klimpel and Ruckert2005). Pawlak et al. (Reference Pawlak, Nadolna-Ałtyn, Szostakowska, Pachur and Podolska2018) found S. entomon infected with H. aduncum in situ in cod stomach, which was the first evidence of such a host–parasite system, similarly to the C. crangon and H. aduncum system in the Baltic Sea presented here.
Experimental research described in the literature confirms that the role of intermediate host may be served by invertebrates, for example, Crustacea, Polychaeta, Ctenophora, Echinodermata, Chaetognatha and Mollusca (Køie, Reference Køie1993; Münster et al., Reference Münster, Klimpel, Fock, MacKenzie and Kuhn2015). The life cycles of Hysterothylacium sp. and potential intermediate hosts were also studied after experimental infection of T. longicornis (Køie and Fagerholm, Reference Køie and Fagerholm1995) and calanoid species (Hurst, Reference Hurst1984; Køie, Reference Køie1993), especially N. integer (Køie and Fagerholm, Reference Køie and Fagerholm1995).
In the Baltic Sea, the life cycle of H. aduncum, and in particular which invertebrate species serve as intermediate hosts for this parasite, has been described only in general terms. The marine environment is changing and new species that act as intermediate hosts might appear. It is known, however, that the eggs of this nematode (which contain developed larvae) can be ingested by both benthic and pelagic crustaceans. Eggs hatch in the intestine of these invertebrates and the parasite larvae migrate to the haemocoel of the intermediate host. Larger invertebrates can play the role of second intermediate hosts (Køie, Reference Køie1993).
Pawlak et al. (Reference Pawlak, Nadolna-Ałtyn, Szostakowska, Pachur and Podolska2018) revealed that moulting and transformation from L4 larva to the adult nematode (H. aduncum) might take place inside invertebrate hosts (S. entomon). Iglesias et al. (Reference Iglesias, Valero, Galvez, Benitez and Adroher2002) conducted in vitro cultivation of H. aduncum from L3 to egg-laying adults and described the conditions (temperature = 13°C; pH = 4; 5% CO2 in the growth atmosphere, etc.) for optimal development and survival of these nematodes. The authors proved that if the medium was supplemented with pepsin, all larvae reached the adult stage. Similar conditions were used for in vitro cultivation of L3-stage H. aduncum larvae obtained from the fish host through a complete developmental cycle of the parasite to L3 larvae hatched from eggs obtained during the experiment (Adroher et al., Reference Adroher, Malagon, Valero and Benitez2004).
In the life cycle of H. aduncum the final hosts are fish. The larval stages live in different tissues of several fish species and in numerous invertebrate species (Norris and Overstreet, Reference Norris and Overstreet1976; Hurst, Reference Hurst1984; Marcogliese, Reference Marcogliese1996). The parasites enter the fish with food and are able to penetrate the stomach wall of the fish to get to the body cavity and internal organs, e.g. liver (Myjak et al., Reference Myjak, Szostakowska, Wojciechowski, Pietkiewicz and Rokicki1994). Sexually mature adult individuals of H. aduncum are often found in the digestive tract of fish (Deardorff and Overstreet, Reference Deardorff and Overstreet1981), such as eelpout and Atlantic cod (Jackson et al., Reference Jackson, Marcogliese and Burt1997), including the Baltic Sea (Køie, Reference Køie1993). Hysterothylacium sp. has also been reported in the Baltic in flatfishes (Køie, Reference Køie1993), eel Anguilla anguilla (Køie, Reference Køie1993), sea trout Salmo trutta trutta (Unger and Palm, Reference Unger and Palm2016), sprat (Skrzypczak and Rolbiecki, Reference Skrzypczak and Rolbiecki2015), sticklebacks (Køie, Reference Køie1993) and Gobiidae fish (Zander et al., Reference Zander, Strohbach and Groenewotd1993, Reference Zander, Groenewold and Strohbach1994; Zander, Reference Zander2004). Gadoids are considered to be the main final hosts for Hysterothylacium sp., however (Berland, Reference Berland1961). Therefore C. crangon might also be the source of infection with this parasite for other listed fish species in the Baltic Sea that feed on the invertebrate.
The emergence of new intermediate hosts is an interesting development, to which the changing climate may be a contributory factor. Rokicki (Reference Rokicki2009) noted that, in general, environmental changes affect the occurrence and abundance of parasites either directly by their influence on the free-living larval stages of parasites or indirectly by their effect on the respective hosts (mainly invertebrate). This problem has already been noted in several parts of the world, such as Australia, where environmental changes have negatively impacted the survival of early-stage parasite larvae in their first intermediate hosts. The absence of Anisakis larvae in fish collected in this area shows the importance of the role of zooplankton and crustaceans in the food chain and in the ecosystem more generally (Shamsi et al., Reference Shamsi, Steller and Chen2018). In Canada, the limited availability of specific food components may be one reason for the decreasing number of parasites in fish (Khan and Chandra, Reference Khan and Chandra2006).
In this study, the parasites Hysterothylacium sp., E. gadi and representatives of Trematoda were found in the stomachs of cod adjacent to food items. These parasites might therefore be present in the body cavity of prey before digestion in the stomach, which could be a source of infection.
It must be emphasized that when cod are caught in a particular area it does not unequivocally indicate that parasites found in its stomach or in its food items were obtained by the fish in the same area. Cod is a migratory species and the adult cod can migrate up to 1000 km (Saulamo and Neuman, Reference Saulamo and Neuman2002) without clear spatial or temporal distribution patterns (Aro, Reference Aro2000).
In summary, to my best knowledge, H. aduncum has been reported in the Baltic Sea in C. crangon for the first time and this also represents the first report of this host–parasite system found in situ in cod stomach. Because C. crangon plays an important role in the food composition of cod, this invertebrate is likely to be an intermediate host in the life cycle of H. aduncum in the Baltic Sea. The results of this research are important for the development of the parasitology, fish biology and ecology of the Baltic Sea, in particular for an improved understanding of the function of individual components of the food web in the transmission of cod parasites.
Acknowledgements
The author is grateful to Professor Magdalena Podolska and Dr Katarzyna Nadolna-Ałtyn for advice, and to Dr Marzenna Pachur for help with cod stomach analysis. The research material was collected as part of the National Programme for Collection of Fisheries Data (EU DCF).
Financial support
This study was supported by the National Science Centre (Poland): grant number 2015/19/N/NZ9/00173.
Conflict of interest
The author declares none.




