


INTRODUCTION
Schistosomes are considered the most important of the human helminth infections in terms of morbidity and mortality. More than 200 million people are infected with schistosomes and a further 800 million are at risk of schistosomiasis in >75 countries in tropical and sub-tropical latitudes. Treatment and control of schistosomiasis rely on the anthelmintic drug praziquantel, but there is concern that drug resistance will appear. New interventions, including vaccines, drugs and diagnostics, are needed for this neglected tropical disease (Hotez et al. Reference Hotez, Brindley, Bethony, King, Pearce and Jacobson2008; Brindley et al. Reference Brindley, Mitreva, Ghedin and Lustigman2009, and references therein).
Draft genome sequences for Schistosoma japonicum and S. mansoni were reported recently, a landmark event that ushered in the post-genomic era for schistosomiasis (Schistosoma japonicum Genome & Functional Analysis Consortium, 2009; Berriman et al. Reference Berriman, Haas, LoVerde, Wilson, Dillon, Cerqueira, Mashiyama, Al-Lazikani, Andrade, Ashton, Aslett, Bartholomeu, Blandin, Caffrey, Coghlan, Coulson, Day, Delcher, DeMarco, Djikeng, Eyre, Gamble, Ghedin, Gu, Hertz-Fowler, Hirai, Hirai, Houston, Ivens, Johnston, Lacerda, Macedo, McVeigh, Ning, Oliveira, Overington, Parkhill, Pertea, Pierce, Protasio, Quail, Rajandream, Rogers, Sajid, Salzberg, Stanke, Tivey, White, Williams, Wortman, Wu, Zamanian, Zerlotini, Fraser-Liggett, Barrell and El-Sayed2009; Han et al. Reference Han, Brindley, Wang and Chen2009). Despite the abundant new datasets, functional analysis of target genes to underpin new interventions for schistosomiasis requires routine approaches for both reverse and forward genetics. To date, functional genomics approaches beyond conventional RNA interference have not been available for schistosomes although reporter plasmids and RNAs have been introduced to several developmental stages (e.g. Davis et al. Reference Davis, Parra, LoVerde, Ribeiro, Glorioso and Hodgson1999; Wippersteg et al. Reference Wippersteg, Kapp, Kunz, Jackstadt, Zahner and Grevelding2002b, Reference Wippersteg, Sajid, Walshe, Khiem, Salter, McKerrow, Grevelding and Caffrey2005; Mourão et al. Reference Mourão, Dinguirard, Franco and Yoshino2009; Krautz-Peterson et al. Reference Krautz-Peterson, Bhardwaj, Faghiri, Tararam and Skelly2010; Stefanic et al. Reference Stefanic, Dvorak, Horn, Braschi, Sojka, Ruelas, Suzuki, Lim, Hopkins, McKerrow and Caffrey2010). Functional genomics including somatic and germ-line transgenesis are desirable because these techniques facilitate validation of essential genes/gene products to be targeted with drugs or vaccines (e.g. van Ooij et al. Reference van Ooij, Tamez, Bhattacharjee, Hiller, Harrison, Liolios, Kooij, Ramesar, Balu, Adams, Waters, Janse and Haldar2008; Homann et al. Reference Homann, Dea, Noble and Johnson2009; Langridge et al. Reference Langridge, Phan, Turner, Perkins, Parts, Haase, Charles, Maskell, Peters, Dougan, Wain, Parkhill and Turner2009; Buguliskis et al. Reference Buguliskis, Brossier, Shuman and Sibley2010). This review addresses genetic manipulation of schistosomes. More specifically, this review focuses on reports of genetic manipulation of schistosomes dealing with approaches targeting integration of transgenes into schistosome chromosomes.
BRIEF HISTORY OF GENETIC MANIPULATION IN SCHISTOSOMES
Advances with genetic manipulation of parasitic helminths including schistosomes have been reviewed (Grevelding, Reference Grevelding, Maule and Marks2006; Brindley and Pearce, Reference Brindley and Pearce2007; Kalinna and Brindley, Reference Kalinna and Brindley2007; Alrefaei et al. Reference Alrefaei, Okatcha, Skinner and Brindley2011; Mann et al. Reference Mann, Suttiprapa, Rinaldi and Brindley2011). In brief, transgenesis of schistosomes was pioneered by Davis and co-workers who bombarded adult stages of S. mansoni with mRNA-encoding firefly luciferase and a luciferase-encoding plasmid (Davis et al. Reference Davis, Parra, LoVerde, Ribeiro, Glorioso and Hodgson1999). Subsequently, Grevelding and colleagues undertook a series of studies employing particle bombardment of S. mansoni stages using plasmids co-precipitated on gold beads. The plasmids encoded fluorescent reporter proteins and were driven by promiscuous (e.g. HSP70) or tissue-specific gene promoters (e.g. cathepsin F) from schistosomes (Wippersteg et al. Reference Wippersteg, Kapp, Kunz and Grevelding2002a, Reference Wippersteg, Kapp, Kunz, Jackstadt, Zahner and Greveldingb, Reference Wippersteg, Ribeiro, Liedtke, Kusel and Grevelding2003, Reference Wippersteg, Sajid, Walshe, Khiem, Salter, McKerrow, Grevelding and Caffrey2005; Rossi et al. Reference Rossi, Wippersteg, Klinkert and Grevelding2003; Dvorak et al. Reference Dvorak, Beckmann, Lim, Engel, Grevelding, McKerrow and Caffrey2010).
Heyers and colleagues introduced plasmid DNA (coated on gold beads) into miracidia, sporocysts and adults of S. mansoni by particle bombardment (Heyers et al. Reference Heyers, Walduck, Brindley, Bleiss, Lucius, Dorbic, Wittig and Kalinna2003). The plasmid construct encoded green fluorescent protein (GFP) under control of the S. mansoni HSP70 (heat shock protein 70 kDa) promoter and termination elements (Wippersteg et al. Reference Wippersteg, Kapp, Kunz, Jackstadt, Zahner and Grevelding2002b). The bombarded larvae and adults expressed GFP, and the transformed miracidia penetrated and established in the intermediate snail host Biomphalaria glabrata. Gold particles were detected in the germ balls of parasites in snail tissue, indicating feasibility of returning transformed parasites to the developmental cycle, a step expected to be useful for establishing lines of transgenic schistosomes.
Correnti and Pearce (Reference Correnti and Pearce2004) demonstrated that square wave electroporation could introduce reporter genes into schistosomes. Subsequently, the technique has found wide acceptance for introduction of plasmids, long dsRNA, siRNA, virions and other reporters into all three major species of human schistosomes, S. mansoni (e.g. Morales et al. Reference Morales, Rinaldi, Gobert, Kines, Tort and Brindley2008; Dvorak et al. Reference Dvorak, Beckmann, Lim, Engel, Grevelding, McKerrow and Caffrey2010), S. japonicum (Zhao et al. Reference Zhao, Lei, Liu, Zhu, Ren, Wang and Shen2008) and S. haematobium (Rinaldi, unpublished). Square wave electroporation has been successfully employed to introduce nucleic acids into eggs, miracidia, sporocysts, schistosomules and adult developmental stages of schistosomes, frequently using a single pulse of 125 volts for 20 milliseconds, in 4 mm gap pathway cuvettes (e.g. Correnti et al. Reference Correnti, Brindley and Pearce2005; Faghiri and Skelly, Reference Faghiri and Skelly2009; Kines et al. Reference Kines, Rinaldi, Okatcha, Morales, Mann, Tort and Brindley2010). Moreover, electroporation has also been employed to develop a method to assess for the presence of an active RNAi pathway by silencing the exogenous reporter gene, firefly luciferase. This straightforward approach offers investigators a means to study the presence of a functional RNAi pathway in less well known parasites and/or to detect the activity of non-conventional interfering molecules such as short hairpin RNAs in schistosomes (Rinaldi et al. Reference Rinaldi, Morales, Cancela, Castillo, Brindley and Tort2008; Ayuk et al. Reference Ayuk, Suttiprapa, Rinaldi, Mann, Lee and Brindley2011).
Findings reported by Grevelding and colleagues are notable in relation to the introduction of transgenes into the germline of schistosomes (Grevelding, Reference Grevelding, Maule and Marks2006; Beckmann et al. Reference Beckmann, Wippersteg, El-Bahay, Hirzmann, Oliveira and Grevelding2007). Plasmids (both supercoiled and linear) encoding GFP were introduced into miracidia by particle bombardment, after which the transformed miracidia infected Biomphalaria glabrata snails by the natural route. Investigation of the cercariae (F0 generation) shed from the snails and adult worms from hamsters infected with the cercariae and of eggs (F1) from the rodents revealed the presence of the GFP transgene. Miracidia harvested from eggs obtained from the livers of the hamsters were used for snail infections and the resulting cercariae were employed to infect hamsters to derive subsequent schistosome generations, F2 and F3. Molecular analyses of F2 and F3 cercariae or adults failed to detect transgenes. Nonetheless, the findings demonstrated that the transgenes were passed from one developmental stage to the next within one generation and, furthermore, from one generation to the next. Since the germ cells are considered to be the only invariable cell type in the developmental cycle of the schistosome, the investigators concluded that the transgenes were present in the germline and their germline-transformation approach had succeeded. Loss or instability of the transgene before (non-Mendelian) inheritance by the F2 progeny likely occurred because transgenes had not integrated into the schistosome genome, a phenomenon well known with extrachromosomal arrays of transgenes in Caenorhabditis elegans (see Semple et al. Reference Semple, Garcia-Verdugo and Lehner2010). Fig. 1 provides a time line - over the past 15 years - of the pioneering and key advances in the genetic manipulation of schistosomes.

Fig. 1. Schematic time line of advances on genetic manipulation of schistosomes. Key events are noted, with pioneering or key reports cited.
SCHISTOSOME TRANSGENESIS WITH INTEGRATION COMPETENT VECTORS
Although approaches to genetic manipulation of schistosomes with non-integration competent vectors have been informative, there are major advantages to genomic integration of transgenes including Mendelian inheritance, sustained transgene activity and transgene-vectored RNA interference (see Giordano-Santini and Dupuy, Reference Giordano-Santini and Dupuy2011). Several classes of integration competent vectors enjoy utility in functional genomics and gene therapy for a spectrum of eukaryotes. These include transposons, gammaretroviruses, lentiviruses and recombinase systems (e.g. see Mates et al. Reference Mates, Izsvak and Ivics2007; Damasceno et al. Reference Damasceno, Beverley and Tosi2010; Turan et al. Reference Turan, Galla, Ernst, Qiao, Voelkel, Schiedlmeier, Zehe and Bode2011). Several of these are now being actively investigated for utility in integrating transgenes into the schistosome genome (Alrefaei et al. Reference Alrefaei, Okatcha, Skinner and Brindley2011).
Retroviruses
Both simple and complex retroviruses (family Retroviridae) are widely employed in functional genomics and gene therapy biotechnologies (e.g. see Hannon and Rossi, Reference Hannon and Rossi2004; Petrus et al. Reference Petrus, Chuah and VandenDriessche2010; Sliva and Schnierle, Reference Sliva and Schnierle2010). The simplex retroviruses include the genus Gammaretrovirus which includes the murine leukaemia virus (MLV). Complex retroviruses include the genus Lentivirus which includes the primate pathogens, HIV and SIV. Attributes of retroviruses that advance their appeal as gene transfer vectors include self-reliant infectious nature, ability to integrate into the chromosomes of the infected cell, potential to be modified to increase host cell range and availability of numerous constructs from commercial sources and academic colleagues.
For safety, retroviral vector systems are usually employed in two components – the retroviral vector, which does not encode viral proteins and the retrovirus packaging cell line, which provides the viral proteins necessary for vector transfer. Infectious, but replication incompetent virions are released from transfected packaging cells. The virus can infect target cells but cannot produce new virions after integration into host chromosome because the integrated provirus does not encode viral proteins (Miller, Reference Miller1992). The restricted host-cell range of retroviral vectors limits their use for stable gene transfer in eukaryotic cells. To overcome this latter limitation, Burns and colleagues pioneered to use of vesicular stomatitis virus glycoprotein (VSVG) pseudotyped murine leukaemia virus (MLV)-derived vectors in which the retroviral envelope glycoprotein is replaced by the glycoprotein (G) of the rhabdovirus, vesicular stomatitis virus (VSV) (Burns et al. Reference Burns, Friedmann, Driever, Burrascano and Yee1993). VSVG is able to bind to phospholipids on membranes of eukaryotic cells at large, endowing the VSVG pseudotyped virion with a potentially very broad range of target tissues and cells (Mastromarino et al. Reference Mastromarino, Conti, Goldoni, Hauttecoeur and Orsi1987; Emi et al. Reference Emi, Friedmann and Yee1991). VSVG pseudotyped virions can infect non-mammalian cells including fish cell lines that are ordinarily refractory to infection because they do not express a cognate receptor for the envelope (surface) protein ligands of the wild type virions.
In our laboratory it has been well established that the infectious replication incompetent MLV retrovirus pseudotyped with VSVG can transduce S. mansoni leading to integration of retroviral transgenes into schistosome chromosomes (Kines et al. Reference Kines, Mann, Morales, Shelby, Kalinna, Gobert, Chirgwin and Brindley2006, Reference Kines, Morales, Mann, Gobert and Brindley2008, Reference Kines, Rinaldi, Okatcha, Morales, Mann, Tort and Brindley2010; Rinaldi et al. Reference Rinaldi, Suttiprapa and Brindley2011). This was notable given it was thought that evolutionary blocks would constrain the utility of MLV in non-mammalian taxa (Dirks and Miller, Reference Dirks and Miller2001). The MLV-derived vector pLNHX was modified to include reporter genes (firefly luciferase or green fluorescent protein) under the control of an endogenous schistosome gene promoter – the RNA polymerase II (Pol II) schistosome actin gene promoter or the RNA polymerase III (Pol III) Spliced Leader (SL) RNA gene promoter. Constructs and the plasmid encoding VSVG were employed to transfect GP2-293 packaging cells modified to express the MLV gag and pol genes (Mann et al. Reference Mann, Morales, Kines and Brindley2008). Eggs, primary sporocysts, schistosomules and adult stages of S. mansoni have been successfully transduced with the VSVG pseudotyped MLV virions. Two-colour immunofluorescence, Southern hybridization and RT-PCR confirmed successful transduction of the schistosomes by this gammaretrovirus. Furthermore, an anchored PCR (retrotransposon-anchored PCR, RAP) approach that employs primers specific for multi-copy endogenous mobile genetic elements interspersed in the schistosome genome was successfully deployed to locate integration junctions of transgenes in the genome of S. mansoni, definitively establishing the presence of proviral MLV transgenes integrated into schistosome chromosomes (Kines et al. Reference Kines, Morales, Mann, Gobert and Brindley2008). In terms of promoters, schistosome actin, HSP70 and spliced leader (SL) gene promoters, as well as the 5′-LTR of MLV, all were found to drive transgene expression in virion-transduced schistosomes (Kines et al. Reference Kines, Mann, Morales, Shelby, Kalinna, Gobert, Chirgwin and Brindley2006, Reference Kines, Morales, Mann, Gobert and Brindley2008).
It is likely that the schistosome cells transduced by the virions were frequently tegumental and/or intestinal cells (Mann et al. Reference Mann, Morales, Kines and Brindley2008). However, we have also fragmented adult worms into several pieces before exposing the (still visibly motile) fragments to virions, which resulted in increased density of transgenes integrated into the schistosome chromosomes (Rinaldi et al. Reference Rinaldi, Suttiprapa and Brindley2011). Nonetheless, in order for heritable transmission to occur, germline transduction would have to have taken place. The schistosome egg represents an advantageous developmental stage at which to direct transgenes (Kines et al. Reference Kines, Rinaldi, Okatcha, Morales, Mann, Tort and Brindley2010; Mann et al. Reference Mann, Suttiprapa, Rinaldi and Brindley2011). Accordingly, we proceeded to transduce schistosome eggs with VSVG-pseudotyped MLV facilitated by electroporation. Square wave electroporation was more effective in delivering VSVG-pseudotyped MLV into schistosome eggs than passive soaking. Quantitative PCR (qPCR) analysis revealed that schistosome eggs electroporated with virions had several fold more copies of provirus than eggs simply soaked in virions (Kines et al. Reference Kines, Rinaldi, Okatcha, Morales, Mann, Tort and Brindley2010). These findings highlight the potential of the schistosome egg as a target into which to deliver chromosomal integration competent transgenes, aiming to establish germline transgenesis in schistosomes.
VSVG-pseudotyped MLV has been employed to transfer transgenes into S. japonicum; Yang et al. (Reference Yang, Brindley, Zeng, Li, Zhou, Liu, Liu, Cai, Zeng, Wei, Lan and McManus2010) transduced schistosomules (perfused from rabbits) with retroviral transgene encoding human telomerase reverse transcriptase (hTERT). RT-PCR, in situ hybridization immunohistochemistry and immunoblot analysis determined that S. japonicum can be successfully transduced with VSVG-pseudotyped MLV and that the MLV vector can transport sizeable genes as cargo – the hTERT gene was ∼3·5 kb in length (Yang et al. Reference Yang, Brindley, Zeng, Li, Zhou, Liu, Liu, Cai, Zeng, Wei, Lan and McManus2010). These findings also suggested the tantalizing possibility of using the hTERT transgene to immortalize cells from schistosome tissues, utilizing the oncogenic potential of hTERT to establish schistosome cell lines.
Transposons
Transposons are naturally occurring mobile genetic elements that move by a cut-and-paste mechanism; they are flanked by inverted terminal repeat (ITR) sequences and mobilized by a transposase encoded by their single open reading frame. There are ∼20 superfamilies of these Class II mobile genetic elements, with member species widespread throughout eukaryote phyla (Feschotte and Pritham, Reference Feschotte and Pritham2007; Yuan and Wessler, Reference Yuan and Wessler2011). Several are known from schistosomes including examples of the Merlin and CACTA groups (Berriman et al. Reference Berriman, Haas, LoVerde, Wilson, Dillon, Cerqueira, Mashiyama, Al-Lazikani, Andrade, Ashton, Aslett, Bartholomeu, Blandin, Caffrey, Coghlan, Coulson, Day, Delcher, DeMarco, Djikeng, Eyre, Gamble, Ghedin, Gu, Hertz-Fowler, Hirai, Hirai, Houston, Ivens, Johnston, Lacerda, Macedo, McVeigh, Ning, Oliveira, Overington, Parkhill, Pertea, Pierce, Protasio, Quail, Rajandream, Rogers, Sajid, Salzberg, Stanke, Tivey, White, Williams, Wortman, Wu, Zamanian, Zerlotini, Fraser-Liggett, Barrell and El-Sayed2009). Transposons can frequently mobilize in species phylogenetically distant from where they were first isolated, a facility which has been harnessed in functional genomics and experimental gene therapy (Plasterk et al. Reference Plasterk, Izsvak and Ivics1999; Ivics et al. Reference Ivics, Li, Mates, Boeke, Nagy, Bradley and Izsvak2009). Accordingly, it is feasible that exogenous transposons might also mobilize in schistosomes and thereby supply integration competent vectors for functional genomics of schistosomes. Several well-studied transposons including piggyBac, Hermes and mariner, are transpositionally active in planarians (Gonzalez-Estevez et al. Reference Gonzalez-Estevez, Momose, Gehring and Salo2003). In evolutionary terms, it is notable that host-parasite interactions play a key role in the horizontal transfer of transposons across phyla (Gilbert et al. Reference Gilbert, Schaack, Pace, Brindley and Feschotte2010).
The piggyBac transposon is used widely in functional genomics and experimental gene therapy (Gonzalez-Estevez et al. Reference Gonzalez-Estevez, Momose, Gehring and Salo2003; Balu et al. Reference Balu, Shoue, Fraser and Adams2005; Wilson et al. Reference Wilson, Coates and George2007). This transposon was isolated from the genome of a moth. It is a short inverted terminal repeat element of 2·5 kb in length with ITRs of 13 bp in length and a single open reading frame encodings the transposase. piggyBac exhibits precise excision upon transposition and affinity for TTAA target sites (Fraser et al. Reference Fraser, Brusca, Smith and Summers1985, Reference Fraser, Ciszczon, Elick and Bauser1996; Cary et al. Reference Cary, Goebel, Corsaro, Wang, Rosen and Fraser1989; Elick et al. Reference Elick, Bauser and Fraser1996). Recently, it has been determined that piggyBac is also active in schistosomes. Morales and colleagues examined whether the piggyBac transposon could deliver reporter transgenes into the genomes of S. mansoni (Morales et al. Reference Morales, Mann, Kines, Gobert, Fraser, Kalinna, Correnti, Pearce and Brindley2007). Linearized piggyBac donor plasmid carrying the firefly luciferase gene as reporter cargo under the control of schistosome gene promoters – actin (pXL-BacII-SmAct-Luc) or HSP70 (pXL-BacII-SmHSP70-Luc) – was introduced together with mRNA encoding the piggyBac transposase into cultured schistosomules by square wave electroporation. Activity of the helper transposase mRNA was confirmed by hybridization of genomic DNA from the transformed schistosomes to a luciferase gene probe. The hybridization signals indicated that the piggyBac transposon had integrated into numerous sites within schistosome chromosomes. Integration events were recovered using an anchored PCR approach employing several endogenous mobile genetic elements from the schistosome genome as anchors, which revealed characteristic piggyBac TTAA footprints in the vicinity of several protein encoding genes, annotated as adenylosuccinate lyase, glutathione peroxidase 1 and glutathione S transferase, as well as loci near endogenous mobile genetic elements including Boudicca and SR2. These findings provided the first direct evidence of somatic transgenesis of schistosomes, or indeed of any parasitic helminth. They demonstrated the transpositional activity of piggyBac in schistosomal tissues, expanding the host range of piggyBac to the digenetic trematodes. Very recently, we reported that vector-based RNAi activity driven by transgene cargo carried by the piggyBac vector pXL-BacII (Ayuk et al. Reference Ayuk, Suttiprapa, Rinaldi, Mann, Lee and Brindley2011) – see below.
Other integration competent vectors
We are unaware of reports of other integrated transgenes in schistosomes beyond the findings with MLV and piggyBac (above). However, it is not unlikely that other retrovirus and transposons, including endogenous schistosome mobile genetic elements, could find utility in genome studies of schistosomes. A potential advantage of endogenous mobile genetic elements is that they may not suffer infection blocks from host restriction/innate immunity factors (e.g. Takeuchi and Matano, Reference Takeuchi and Matano2008; Strebel et al. Reference Strebel, Luban and Jeang2009). DNA site-specific recombinases (SSRs), such as Cre, FLPe and φC31, from bacteriophages of fungi, and other provenances are influential tools for analyzing gene function in vertebrates (e.g. see Bischof and Basler, Reference Bischof and Basler2008) and might also be active in schistosomes. Several are now in service for site-specific gene manipulation of Plasmodium falciparum (Adjalley et al. Reference Adjalley, Lee and Fidock2010; O'Neill et al. Reference O'Neill, Phuong, Healer, Richard and Cowman2011).
In overview, transgenesis mediated by integration competent vectors such as MLV and piggyBac can provide a routine functional genomics platform for forward and reverse genetics of schistosomes. Forward genetics, where specimens displaying a mutant phenotype after insertional (or chemical, etc.) mutagenesis are selected – the ‘from phenotype to genotype’ approach – can now be attempted using transduction of schistosomes (e.g. with MLV or piggyBac) followed by high throughput sequence analysis of the schistosome genome in similar fashion to other pathogens (e.g. Langridge et al. Reference Langridge, Phan, Turner, Perkins, Parts, Haase, Charles, Maskell, Peters, Dougan, Wain, Parkhill and Turner2009). Further, given that draft schistosome genomes are now available, MLV or piggyBac can be used for reverse genetics. With reverse genetics, functional analysis involves targeting a known gene sequence for inactivation where the function of the gene of interest is then inferred from the resulting phenotype (‘from genotype to phenotype’ approach) (Boutros and Ahringer, Reference Boutros and Ahringer2008). Conventional RNAi and more recently vector-based RNAi in schistosomes are the reverse genetic tools of choice for reverse genetics, approaches widely used for discovery of targets for experimental drug and/or vaccine development (e.g. Sayed et al. Reference Sayed, Cook and Williams2006; Mourão et al. Reference Mourão, Dinguirard, Franco and Yoshino2009; Stefanic et al. Reference Stefanic, Dvorak, Horn, Braschi, Sojka, Ruelas, Suzuki, Lim, Hopkins, McKerrow and Caffrey2010).
VECTOR-BASED RNAi
Experimental RNAi works well in schistosomes, in general (see Krautz-Peterson et al. Reference Krautz-Peterson, Bhardwaj, Faghiri, Tararam and Skelly2010). Skelly and co-workers and Boyle and colleagues first described successful knockdown in S. mansoni (Boyle et al. Reference Boyle, Wu, Shoemaker and Yoshino2003; Skelly et al. Reference Skelly, Da'dara and Harn2003), and since then numerous reports describing endogenous and reporter gene knockdown in S. mansoni and S. japonicum have appeared (e.g. Kumagai et al. Reference Kumagai, Osada, Ohta and Kanazawa2009; Rinaldi et al. Reference Rinaldi, Morales, Alrefaei, Cancela, Castillo, Dalton, Tort and Brindley2009). RNAi is active in S. haematobium (Rinaldi and co-workers, unpublished). However, conventional RNAi by double stranded RNA frequently leads to transient gene silencing and, in addition, may be inaccessible to some developmental stages and/or tissues of schistosomes. In vivo, e.g. vector-based RNAi approaches that lead to integration of transgenes encoding cassettes that express short interfering RNAs can circumvent deficiencies with exogenous RNAi approaches by providing continuous and/or conditional gene silencing (see Sliva and Schnierle, Reference Sliva and Schnierle2010). In brief, these experimental systems frequently employ a gene construct encoding an oligonucleotide of the target siRNA, a short loop domain (∼9 residues), followed by the reverse complement of the siRNA, and driven by a Pol III (or Pol II) promoter. The construct can then be introduced into target cells for endogenous expression of shRNA targeting the gene of interest. The shRNA is processed in the cytoplasm to siRNA (Manjunath and Dykxhoorn, Reference Manjunath and Dykxhoorn2010). Both plasmid-based and retroviral (integrating) vectors are widely used for vector-based RNAi procedures, the latter offering long term gene silencing of expression (Couto and High, Reference Couto and High2010; Sliva and Schnierle, Reference Sliva and Schnierle2010). Zhao and co-workers pioneered the approach in schistosomes, demonstrating silencing of the expression of the Mago nashi gene in S. japonicum by siRNAs derived from shRNA expressed by mammalian Pol III promoter H1 (Zhao et al. Reference Zhao, Lei, Liu, Zhu, Ren, Wang and Shen2008). We recently demonstrated that MLV encoding long hairpin RNAs, ∼120 bp long (hpRNA), driven by a RNA polymerase II promoter (S. mansoni actin) targeting S. mansoni cathepsin B in the adult stage of S. mansoni delivered silencing of the protease (Fig. 2, left panel) (Tchoubrieva et al. Reference Tchoubrieva, Ong, Pike, Brindley and Kalinna2010). On the other hand, in many species including insects, mammals, birds and pathogenic protozoa, Pol III promoter-based DNA vectors have been employed to express small interfering RNA (siRNA) or short hairpin RNA (∼21 bp long) (shRNA) (Lambeth et al. Reference Lambeth, Moore, Muralitharan, Dalrymple, McWilliam and Doran2005; Wakiyama et al. Reference Wakiyama, Matsumoto and Yokoyama2005; Wise et al. Reference Wise, Schafer, Lambeth, Tyack, Bruce, Moore and Doran2007; Linford et al. Reference Linford, Moreno, Good, Zhang, Singh and Petri2009). Aiming to establish vector-based RNAi driven by a Pol III promoter, we cloned S. mansoni and human U6 gene promoters (∼270 bp) into pLNHX driving shRNA targeting firefly luciferase. We targeted luciferase because the effect of RNAi against luciferase can be readily discerned (in contrast to many endogenous genes) (Rinaldi et al. Reference Rinaldi, Morales, Cancela, Castillo, Brindley and Tort2008). Luciferase activity was significantly reduced in worms transduced with piggyBac encoding shLuc (Fig. 2, right) (Ayuk et al. Reference Ayuk, Suttiprapa, Rinaldi, Mann, Lee and Brindley2011).

Fig. 2. Vector-based RNA interference (RNAi) in Schistosoma mansoni. Panel A: Vector-based RNAi mediated by long hairpin RNAs (hpRNA) driven by a RNA polymerase II promoter (from the S. mansoni actin gene) carried by a retroviral vector to knock down of the S. mansoni cathepsin B1. Top: schematic representation of retroviral vector construct and dsRNA hairpin. Bottom: knock down of S. mansoni cathepsinB1. Panel B: Vector-based RNAi mediated by short hairpin RNAs (shRNA) driven by a RNA polymerase III promoter (from the S. mansoni U6 gene) carried by the piggyBac donor plasmid to knock down of the exogenous reporter gene firefly luciferase. Top: schematic representation of construct encoding the shRNA. Bottom: knock down of reporter firefly luciferase activity. Adapted from Tchoubrieva et al. (Reference Tchoubrieva, Ong, Pike, Brindley and Kalinna2010) and Ayuk et al. (Reference Ayuk, Suttiprapa, Rinaldi, Mann, Lee and Brindley2011) with permission.
DEVELOPMENTAL STAGES OF SCHISTOSOMES FOR TRANSGENESIS
Since developmental cycles of the three major schistosomes of humans can be accomplished using laboratory rodents as the mammalian hosts and laboratory-reared snails as the intermediate hosts (Lewis, Reference Lewis, Coligan, Kruisbeek, , Margulies, Shevach and Strober1998), most developmental stages of these schistosomes are theoretically accessible to genetic manipulation (Fig. 3). Moreover, some stages can be cultured ex vivo or in vitro and returned to the snails or mice to continue development (see Mann et al. Reference Mann, Morales, Rinaldi and Brindley2010). On the other hand, discrete stages are differentially accessible to delivery of transgenes using approaches including particle bombardment, square wave electroporation, cationic polymer-based gene delivery, and/or transduction by virions or other infectious agents (Heyers et al. Reference Heyers, Walduck, Brindley, Bleiss, Lucius, Dorbic, Wittig and Kalinna2003; Beckmann et al. Reference Beckmann, Wippersteg, El-Bahay, Hirzmann, Oliveira and Grevelding2007; Mann et al. Reference Mann, Morales, Kines and Brindley2008). Other approaches, such as microinjection, should be of value, as indicated by progress with introduction of transgenes in tapeworms and parasitic nematodes (Spiliotis et al. Reference Spiliotis, Mizukami, Oku, Kiss, Brehm and Gottstein2010).
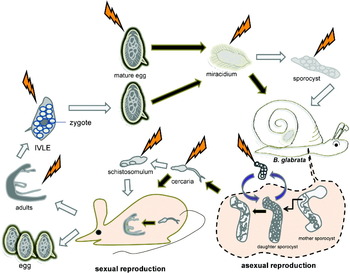
Fig. 3. Cartoon representation of points of the developmental cycle of Schistosoma mansoni amenable to genetic manipulation. Both the mammalian stages involved in sexual reproduction and the snail stages with asexual reproduction are presented. Thunder bolts suggest accessible points of introduction of transgenes into the schistosome e.g. transgene delivery by electroporation, microinjection, etc. Black arrows indicate processes that occur naturally whereas white arrows represent processes that can be manipulated. Dashed line indicates events inside the snail. IVLE, in vitro laid eggs.
Adult worms can be obtained from infected rodents and can be maintaining in culture. ‘Viable’ fragments of worms – obtained by dicing adult schistosomes into several pieces – can be used as a study model as well (Rinaldi et al. Reference Rinaldi, Suttiprapa and Brindley2011); whereas these fragments are not as tractable as primary cell cultures of Echinococcos multilocularis (Spiliotis et al. Reference Spiliotis, Mizukami, Oku, Kiss, Brehm and Gottstein2010) or fragments of planarians (Shibata et al. Reference Shibata, Rouhana and Agata2010), they do allow access to internal organs and cells of the schistosome. The schistosome egg and the miracidium have desirable attributes for consideration in relation to genetic manipulation, these include the presence of a single celled zygote within the egg-shell upon its release from the blood fluke (Jurberg et al. Reference Jurberg, Goncalves, Costa, de Mattos, Pascarelli, de Manso, Ribeiro-Alves, Pelajo-Machado, Peralta, Coelho and Lenzi2009), favourably high ratio of germ to somatic cells even as it develops and ease of maintenance in vitro. Primary sporocysts transformed from miracidia in vitro are worthy targets for genomic manipulation because this developmental stage can be transplanted into Biomphalaria glabrata snails to establish lines of S. mansoni (Kapp et al. Reference Kapp, Coustau, Wippersteg, Jourdane, Kunz and Grevelding2003). Schistosomules obtained by mechanical transformation of cercariae shed from snails have been used to investigate the activity of transgenes and/or schistosome gene promoters driving transgenes (Correnti et al. Reference Correnti, Brindley and Pearce2005, Reference Correnti, Jung, Freitas and Pearce2007; Morales et al. Reference Morales, Mann, Kines, Gobert, Fraser, Kalinna, Correnti, Pearce and Brindley2007).
The ‘in vitro laid egg’ (IVLE) deserves special mention. Pearce and colleagues demonstrated that eggs develop after release from adult schistosomes in vitro (Freitas et al. Reference Freitas, Jung and Pearce2007). Eggs released from the fertilized adult female schistosome can develop in vitro and eventually release viable miracidia, provided that the eggs have been laid soon after the adult worms have been perfused from experimentally infected rodents. (From about 48 hours after perfusion from mice, eggs shed from worms exhibit reduced viability.) Notably, when released from the female worm, the schistosome egg includes a single cell zygote, in which cleavage has yet to take place (Jurberg et al. Reference Jurberg, Goncalves, Costa, de Mattos, Pascarelli, de Manso, Ribeiro-Alves, Pelajo-Machado, Peralta, Coelho and Lenzi2009). Accordingly, introduction of transgenes into this young egg may be able to accomplish germ (and somatic) transgenesis in a developmental stage that seems to be reasonably accessible in the laboratory (Mann et al. Reference Mann, Suttiprapa, Rinaldi and Brindley2011).
Finally, it is obvious that the availability of immortalized cell lines would enhance functional genomics investigations (Brindley and Pearce, Reference Brindley and Pearce2007). Unfortunately, none are yet available. Progress with primary cell cultures in related flatworms (Spiliotis et al. Reference Spiliotis, Mizukami, Oku, Kiss, Brehm and Gottstein2010) indicates that cell lines can be established and perhaps progress with transgenesis of schistosomes with oncogenes such as hTERT (Yang et al. Reference Yang, Brindley, Zeng, Li, Zhou, Liu, Liu, Cai, Zeng, Wei, Lan and McManus2010) will provide a route forward in this area.
PERSPECTIVE
We have reviewed advances in functional genomics and transgenesis of schistosomes, focusing on approaches leading to chromosomal integration of transgenes. The retrovirus MLV and the transposon piggyBac have now both been shown to integrate reliably into the chromosomes of S. mansoni and hence both exhibit great potential as vectors to drive functional genomics for schistosomes. However, improvements are needed to establish transgenic schistosomes and protocols. An impediment has been the difficulty of delivering transgenes to the germline. Targeting integration competent transgenes to IVLE may surmount this roadblock (Mann et al. Reference Mann, Suttiprapa, Rinaldi and Brindley2011). Other gateways to the schistosome germline include the daughter sporocysts where the germ cells are comparatively massive (see Coustau and Yoshino, Reference Coustau and Yoshino2000). We envisage that advances in technologies which will drive functional genomics forward quickly, including expansion of in vivo RNAi, high-throughput insertional mutagenesis and, hopefully, gains-of-function approaches involving drug selection of transgenic schistosomes. Advances in S. mansoni can be expected to be adapted to the other schistosomes, to the food-borne flukes such as Opisthorchis viverrini, Clonorchis sinensis and Fasciola hepatica, and other helminth parasites at large.
ACKNOWLEDGEMENTS
Support from NIH-NIAID award R01AI072773 is gratefully acknowledged. We thank Tunika Okatcha and Danielle Skinner for critical review of the manuscript.



