







Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Ruman, Tomasz
Długopolska, Karolina
Jurkiewicz, Agata
Rut, Dagmara
Frączyk, Tomasz
Cieśla, Joanna
Leś, Andrzej
Szewczuk, Zbigniew
and
Rode, Wojciech
2010.
Thiophosphorylation of free amino acids and enzyme protein by thiophosphoramidate ions.
Bioorganic Chemistry,
Vol. 38,
Issue. 2,
p.
74.
Dąbrowska, Magdalena
Gołos, Barbara
Wałajtys-Rode, Elżbieta
Wińska, Patrycja
Cieśla, Joanna
Zieliński, Zbigniew
Jagielska, Elżbieta
and
Rode, Wojciech
2010.
Detection of Bacteria, Viruses, Parasites and Fungi.
p.
333.
Dąbrowska-Maś, Elżbieta
Frączyk, Tomasz
Ruman, Tomasz
Radziszewska, Karolina
Wilk, Piotr
Cieśla, Joanna
Zieliński, Zbigniew
Jurkiewicz, Agata
Gołos, Barbara
Wińska, Patrycja
Wałajtys-Rode, Elżbieta
Leś, Andrzej
Nizioł, Joanna
Jarmuła, Adam
Stefanowicz, Piotr
Szewczuk, Zbigniew
and
Rode, Wojciech
2012.
Tyrosinenitration affects thymidylate synthase properties.
Org. Biomol. Chem.,
Vol. 10,
Issue. 2,
p.
323.
Gołos, Barbara
Dąbrowska, Magdalena
Wałajtys-Rode, Elżbieta
Zieliński, Zbigniew
Wińska, Patrycja
Cieśla, Joanna
Jagielska, Elżbieta
Moczoń, Tadeusz
and
Rode, Wojciech
2012.
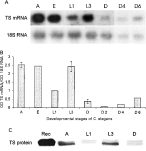
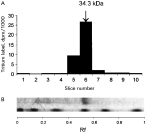
Immunofluorescent localization of thymidylate synthase in the development of Trichinella spiralis and Caenorhabditis elegans.
Molecular and Biochemical Parasitology,
Vol. 183,
Issue. 1,
p.
63.
Wilk, Piotr
Jarmuła, Adam
Ruman, Tomasz
Banaszak, Katarzyna
Rypniewski, Wojciech
Cieśla, Joanna
Dowierciał, Anna
and
Rode, Wojciech
2014.
Crystal structure of phosphoramide-phosphorylated thymidylate synthase reveals pSer127, reflecting probably pHis to pSer phosphotransfer.
Bioorganic Chemistry,
Vol. 52,
Issue. ,
p.
44.
Frączyk, Tomasz
Ruman, Tomasz
Wilk, Piotr
Palmowski, Paweł
Rogowska-Wrzesinska, Adelina
Cieśla, Joanna
Zieliński, Zbigniew
Nizioł, Joanna
Jarmuła, Adam
Maj, Piotr
Gołos, Barbara
Wińska, Patrycja
Ostafil, Sylwia
Wałajtys-Rode, Elżbieta
Shugar, David
and
Rode, Wojciech
2015.
Properties of phosphorylated thymidylate synthase.
Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics,
Vol. 1854,
Issue. 12,
p.
1922.
Ludwiczak, Jan
Maj, Piotr
Wilk, Piotr
Frączyk, Tomasz
Ruman, Tomasz
Kierdaszuk, Borys
Jarmuła, Adam
and
Rode, Wojciech
2016.
Phosphorylation of thymidylate synthase affects slow-binding inhibition by 5-fluoro-dUMP and N4-hydroxy-dCMP.
Molecular BioSystems,
Vol. 12,
Issue. 4,
p.
1333.
Jarmuła, Adam
Wilk, Piotr
Maj, Piotr
Ludwiczak, Jan
Dowierciał, Anna
Banaszak, Katarzyna
Rypniewski, Wojciech
Cieśla, Joanna
Dąbrowska, Magdalena
Frączyk, Tomasz
Bronowska, Agnieszka K.
Jakowiecki, Jakub
Filipek, Sławomir
and
Rode, Wojciech
2017.
Crystal structures of nematode (parasitic T. spiralis and free living C. elegans ), compared to mammalian, thymidylate synthases (TS). Molecular docking and molecular dynamics simulations in search for nematode-specific inhibitors of TS.
Journal of Molecular Graphics and Modelling,
Vol. 77,
Issue. ,
p.
33.
Uram, Łukasz
Nizioł, Joanna
Maj, Piotr
Sobich, Justyna
Rode, Wojciech
and
Ruman, Tomasz
2017.
N(4)-[B-(4,4,5,5-tetramethyl-1,3,2-dioxaborolan)methyl]-2′-deoxycytidine as a potential boron delivery agent with respect to glioblastoma.
Biomedicine & Pharmacotherapy,
Vol. 95,
Issue. ,
p.
749.
Markowicz, Joanna
Uram, Łukasz
Sobich, Justyna
Mangiardi, Laura
Maj, Piotr
and
Rode, Wojciech
2019.
Antitumor and anti-nematode activities of α-mangostin.
European Journal of Pharmacology,
Vol. 863,
Issue. ,
p.
172678.
Maj, Piotr
Mori, Mattia
Sobich, Justyna
Markowicz, Joanna
Uram, Łukasz
Zieliński, Zbigniew
Quaglio, Deborah
Calcaterra, Andrea
Cau, Ylenia
Botta, Bruno
and
Rode, Wojciech
2020.
Alvaxanthone, a Thymidylate Synthase Inhibitor with Nematocidal and Tumoricidal Activities.
Molecules,
Vol. 25,
Issue. 12,
p.
2894.
Maj, Piotr
Jarmuła, Adam
Wilk, Piotr
Prokopowicz, Małgorzata
Rypniewski, Wojciech
Zieliński, Zbigniew
Dowierciał, Anna
Bzowska, Agnieszka
and
Rode, Wojciech
2021.
Molecular Mechanism of Thymidylate Synthase Inhibition by N4-Hydroxy-dCMP in View of Spectrophotometric and Crystallographic Studies.
International Journal of Molecular Sciences,
Vol. 22,
Issue. 9,
p.
4758.
El Saftawy, E. A.
Aboulhoda, B. E.
Hassan, F. E.
Ismail, M. A. M.
Alghamdi, M. A.
Hussein, S. M.
and
Amin, N. M.
2024.
ACV with/without IVM: a new talk on intestinal CDX2 and muscular CD34 and Cyclin D1 during Trichinella spiralis infection.
Helminthologia,
Vol. 61,
Issue. 2,
p.
124.