



Introduction
Nearly 1.7 billion cases of diarrheal disease are reported globally every year, and its socioeconomic burden on health services has been estimated at 72.8 million disability-adjusted life years annually (Ryan et al., Reference Ryan, Paparini and Oskam2017). Enteric protozoan parasites are among the major contributors to this diarrheal disease load (Fletcher et al., Reference Fletcher, Stark, Harkness and Ellis2012; Di Genova and Tonelli, Reference Di Genova and Tonelli2016). Cyclospora cayetanensis is an important global pathogen in humans, typically causing prolonged diarrhea accompanied by anorexia, malaise, nausea and cramping, among other symptoms (Shields and Olson, Reference Shields and Olson2003; Giangaspero and Gasser, Reference Giangaspero and Gasser2019). Many large cyclosporiasis outbreaks have been documented in industrialized nations (Ortega and Sanchez, Reference Ortega and Sanchez2010). In these, food has been identified as the main vehicle for Cyclospora transmission, according to source-tracing studies (Herwaldt and Ackers, Reference Herwaldt and Ackers1997; Ortega and Sanchez, Reference Ortega and Sanchez2010). Cilantro from Mexico was identified as one of the possible sources of a cyclosporiasis outbreak in the United States (USA) in 2013, with more than 600 cases of infection (Abanyie et al., Reference Abanyie, Harvey, Harris, Wiegand, Gaul, Desvignes-Kendrick, Irvin, Williams, Hall, Herwaldt, Gray, Qvarnstrom, Wise, Cantu, Cantey, Bosch, DA Silva, Fields, Bishop, Wellman, Beal, Wilson, Fiore, Tauxe, Lance, Slutsker and Parise2015). More recently, prepackaged vegetable trays and vegetable salads sold at a fast food chain have been the suspected sources of cyclosporiasis outbreaks in June and July, 2018, according to trace-back investigations (Casillas et al., Reference Casillas, Bennett and Straily2018).
Up to 31 December 2018, more than one thousand papers have been published on Cyclospora. Numerous studies of Cyclospora infections among travelers, immunodeficient patients, diarrheal and asymptomatic patients and the residents of disease-endemic areas have been reported. In this study, we review the biological characteristics, clinical features, epidemiology, detection methods and treatment of C. cayetanensis, and assess some risk factors for human infection with this foodborne pathogen.
Biological characteristics
History of discovery and research
The genus Cyclospora, created by Schneider in 1881, was first described by Eimer in 1870 (Ortega and Sanchez, Reference Ortega and Sanchez2010). Until the 1990s, the genus only included species that infect animals, such as rodents, insectivores and reptiles (Casemore, Reference Casemore1994). The earliest description of human infection with Cyclospora was from Papua New Guinea in 1979 (Ashford, Reference Ashford1979). Oocysts were subsequently observed in the faeces of patients from Haiti and Peru in 1983–1985, American travelers returning from Haiti and Mexico in 1986, British travelers who became ill in Nepal in 1989 and travelers and foreign residents in Nepal in 1993 (Herwaldt, Reference Herwaldt2000), although the identity of the pathogen was uncertain at that time. In 1994, Ortega et al. named this human causative organism C. cayetanensis (Ortega and Sanchez, Reference Ortega and Sanchez2010).
Cyclospora cayetanensis has received further attention since the first outbreak of Cyclospora-associated diarrheal illness in the USA in 1990 (Huang et al., Reference Huang, Weber, Sosin, Griffin, Long, Murphy, Kocka, Peters and Kallick1995). In 1996, more than 1400 cases of cyclosporiasis were reported in the USA and Canada (Herwaldt and Ackers, Reference Herwaldt and Ackers1997). Since then, very large studies of Cyclospora infection among travelers, immunodeficient patients, diarrheal patients and asymptomatic individuals have been reported, as have studies of detection methods and treatment measures for Cyclospora.
Morphology and taxonomy
Cyclospora cayetanensis is the only documented Cyclospora species infecting humans, and it is widely accepted that among common mammals, only humans are susceptible to infection by this microbe (Ortega and Sanchez, Reference Ortega and Sanchez2010).
Under light microscopy, C cayetanensis oocysts have a spheroid shape, 8–10 µm in diameter, with indistinguishable protoplasm (Fig. 1). When sporulated, each oocyst contains two ovoid sporocysts that, in turn, contain two sporozoites each (Ortega and Sanchez, Reference Ortega and Sanchez2010). Cyclospora oocysts are modified with Ziehl–Neelsen acid-fast stain in different ways: some stain dark red with a mottled appearance, some stain pink, whereas others do not stain all and appear as non-refractile glassy spheres against the blue-green background (Clarke and McIntyre, Reference Clarke and McIntyre1996; Zhou et al., Reference Zhou, Lv, Wang, Wang, Jian, Zhang, Ning, Fu, Wang, Qi, Yao, Zhao, Zhang, Sun, Shi, Arrowood and Xiao2011). Their autofluorescence makes C. cayetanensis oocysts readily visible in clinical samples with epifluorescence microscopy under a 330–380 nm ultraviolet (UV) filter (Zhou et al., Reference Zhou, Lv, Wang, Wang, Jian, Zhang, Ning, Fu, Wang, Qi, Yao, Zhao, Zhang, Sun, Shi, Arrowood and Xiao2011).
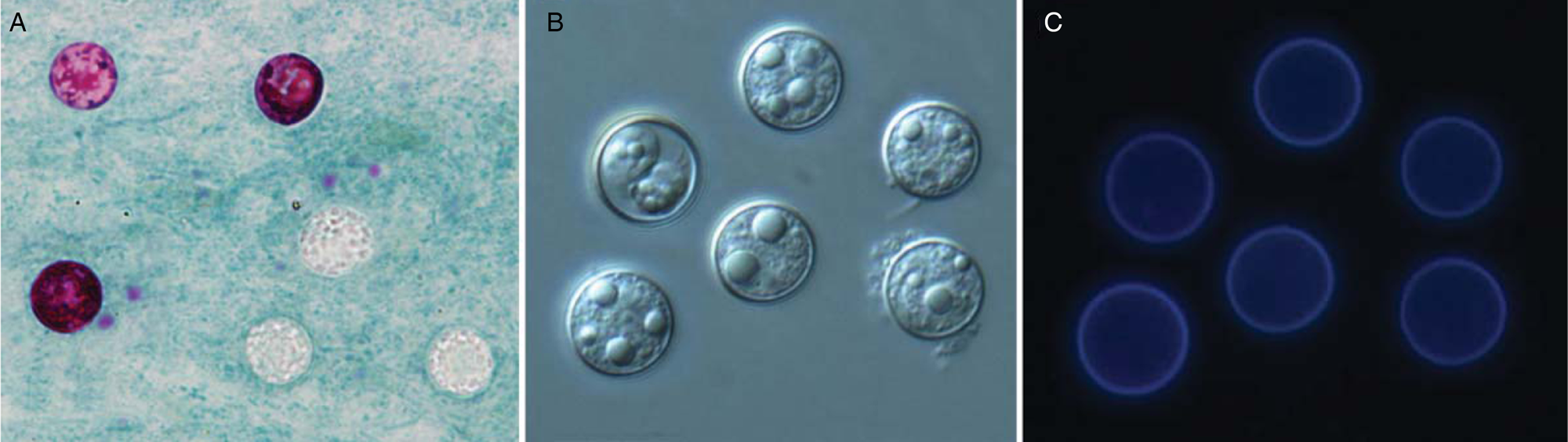
Fig. 1. Morphology of C. cayetanensis oocysts under microscopy. Oocysts in stool smears stained with modified acid-fast stain under light microscopy; two oocysts are stained with different intensities (A); differential interference contrast microscopy of wet mounts, a partially sporulated oocyst can be seen (B); epifluorescence microscopy with a 330–380 nm UV excitation filter (C).
Cyclospora cayetanensis belongs to the subphylum Apicomplexa, subclass Coccidiasina, family Eimeriidae and genus Cyclospora (Ortega and Sanchez, Reference Ortega and Sanchez2010). Phylogenetic analyses have shown that human-associated Cyclospora is closely related to members of the genus Eimeria (Fig. 2) (Relman et al., Reference Relman, Schmidt, Gajadhar, Sogin, Cross, Yoder, Sethabutr and Echeverria1996; Liu et al., Reference Liu, Wang, Zheng, Xu, Roellig, Li, Frace, Tang, Arrowood, Moss, Zhang, Feng and Xiao2016). Cyclospora cercopitheci in vervet monkeys (Cercopithecus aethiops), C. colobi in colobus monkeys (Colobus guereza) and C. papionis in olive baboons (Papio anubis) were characterized in 1999 (Eberhard et al., Reference Eberhard, da Silva, Lilley and Pieniazek1999); C. macacae was described in rhesus monkeys (Macaca mulatta) in 2015 (Li et al., Reference Li, Ye, Arrowood, Ma, Wang, Xu, Feng and Xiao2015); and C. duszynskii and C. yatesi were characterized in moles (Scalopus aquaticus) in 2018 (McAllister et al., Reference McAllister, Motriuk-Smith and Kerr2018). A total of 22 Cyclospora species have so far been described in vipers, moles, myriapodes, rodents, monkeys and humans (Lainson, Reference Lainson2005; Li et al., Reference Li, Ye, Arrowood, Ma, Wang, Xu, Feng and Xiao2015; McAllister et al., Reference McAllister, Motriuk-Smith and Kerr2018). However, Cyclospora-like organisms have also been described in dogs, cattle, chickens, rats, house mice, birds, monkeys, shellfish, etc., and even in environmental samples (Sherchand and Cross, Reference Sherchand and Cross2001; Chu et al., Reference Chu, Sherchand, Cross and Orlandi2004; Li et al., Reference Li, Xiao, Zhou, Li and Wadeh2007; Cordón et al., Reference Cordón, Prados, Romero, Sánchez Moreno, Pontes, Osuna and Rosales2008; Aksoy et al., Reference Aksoy, Marangi, Papini, Ozkoc, Bayram Delibas and Giangaspero2014; Helenbrook et al., Reference Helenbrook, Wade, Shields, Stehman and Whipps2015; Ghozzi et al., Reference Ghozzi, Marangi, Papini, Lahmar, Challouf, Houas, Ben Dhiab, Normanno, Babba and Giangaspero2017).
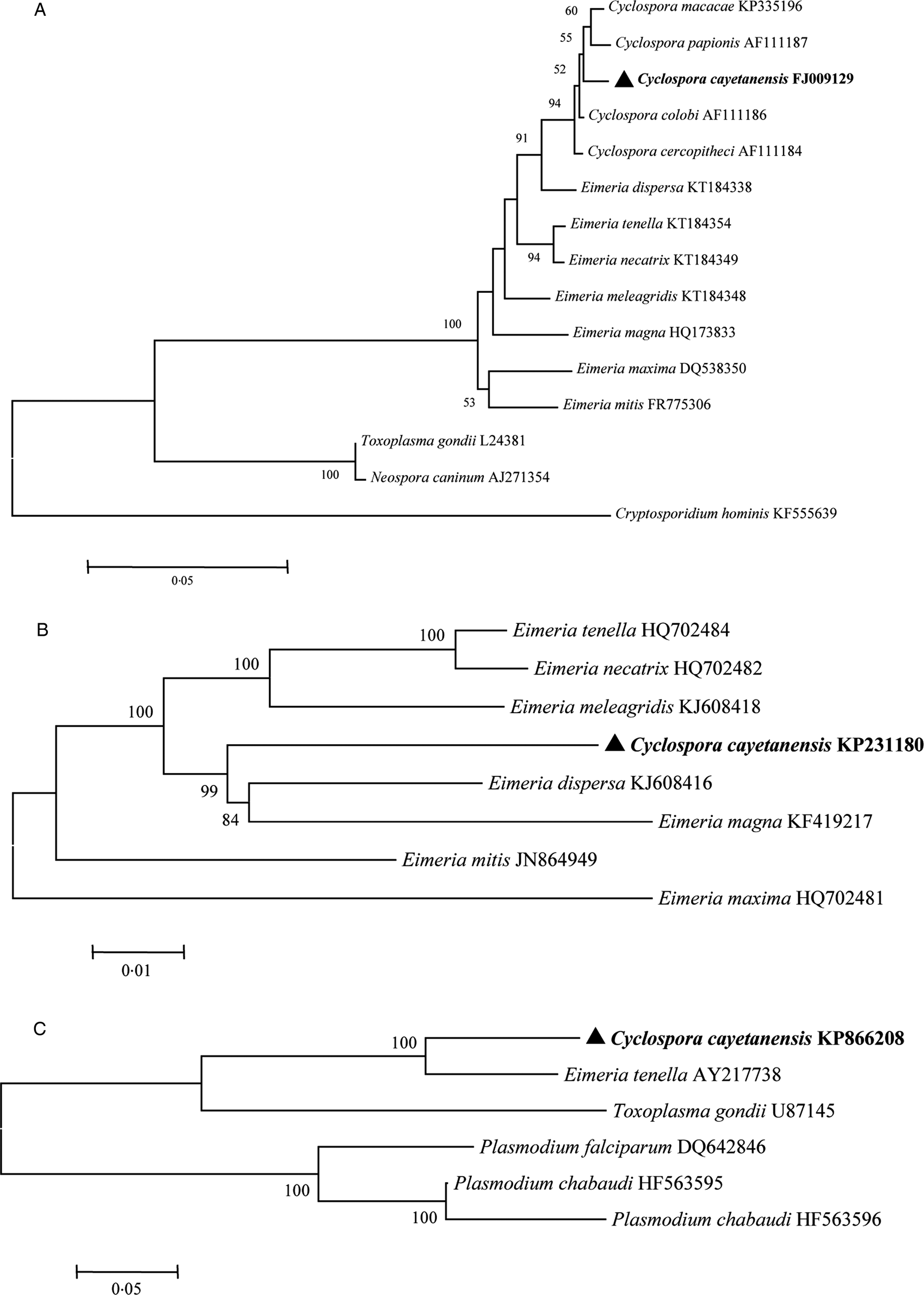
Fig. 2. Phylogenetic relationships of C. cayetanensis and other apicomplexan protozoa. Phylogeny inferred with a neighbour-joining analysis of small-subunit ribosomal RNA gene sequences (A) reported by Li et al. (Reference Li, Chang, Shi, Wang, Fu, Li, Xu, Jia, Guo and Zhang2017); mitochondrial genomes (B) reported by Cinar et al. (Reference Cinar, Gopinath, Jarvis and Murphy2015) and apicoplast genomes (C) reported by Tang et al. (Reference Tang, Guo, Zhang, Rowe, Roellig, Frace, Li, Liu, Feng and Xiao2015), based on distances calculated with the Kimura 2-parameter model. Bootstrap values >50% from 1000 replicates are shown at the nodes. Scale bars indicate estimated substitutions per site.
Life cycle of C. cayetanensis
Infections of C. cayetanensis mainly occur via the faecal–oral transmission route. Fresh (unsporulated) oocysts are excreted in stools. Oocysts are spheroid, 8–10 µm in diameter, and contain indistinguishable protoplasm (Brown and Rotschafer, Reference Brown and Rotschafer1999). In the environment outside the host, freshly excreted oocysts are not infectious until their sporulation is complete, which occurs within a few days to weeks (at maximum) at temperatures between 22 and 30 °C. Storage at either 4 or 37 °C retards sporulation (Smith et al., Reference Smith, Paton, Mitambo and Girdwood1997). The sporulation of the oocysts occurs irrespective of whether they are stored in deionized water or potassium dichromate solution, and results in the division of the sporont into two sporocysts, each containing two elongated sporozoites (Smith et al., Reference Smith, Paton, Mitambo and Girdwood1997). During this time, food or water can act as the vehicle for Cyclospora transmission. Once the sporulated oocysts in food, water or soil are ingested by a new host, the mature oocysts usually excyst in the small bowel, and sporozoites are released to invade the epithelial cells of the upper small intestine (duodenum or jejunum) (Ortega and Sanchez, Reference Ortega and Sanchez2010).
The presence of asexual and sexual stages in the same host suggests that the life cycle of this microorganism can be completed within one host (Ortega et al., Reference Ortega, Nagle, Gilman, Watanabe, Miyagui, Quispe, Kanagusuku, Roxas and Sterling1997). The intracellular developmental stages begin with the formation of intracytoplasmic parasitophorous vacuoles in the intestinal epithelium cells (Sun et al., Reference Sun, Ilardi, Asnis, Bresciani, Goldenberg, Roberts and Teichberg1996; Ortega and Sanchez, Reference Ortega and Sanchez2010), which are sometimes also observed in biliary epithelium cells (Zar et al., Reference Zar, El-Bayoumi and Yungbluth2001). Asexual multiplication results in type I and II meronts (Ortega et al., Reference Ortega, Nagle, Gilman, Watanabe, Miyagui, Quispe, Kanagusuku, Roxas and Sterling1997). Type I meronts give rise to 8–12 merozoites that then infect neighbouring epithelial cells, and this type of asexual reproduction is often quite prolific. Type II meronts form later, releasing four merozoites to invade neighbouring cells. Some of these meronts form macrogametes, whereas others undergo multiple fission events to form microgametocytes containing flagellated microgametes (Ortega et al., Reference Ortega, Nagle, Gilman, Watanabe, Miyagui, Quispe, Kanagusuku, Roxas and Sterling1997). The macrogametocyte is fertilized by the microgametocyte, producing a zygote, in the sexual stages. Once fertilization occurs, an environmentally resistant wall is formed, and the oocyst is excreted from the host into the environment as an unsporulated oocyst in the faeces (Shields and Olson, Reference Shields and Olson2003; Ortega and Sanchez, Reference Ortega and Sanchez2010).
Molecular characteristics
The characteristics of the polymorphic regions of the Cyclospora genome have been studied to better understand the microorganism's mode of infection and epidemiology. Small subunit ribosomal RNA (SSU rRNA) gene sequences show minimal genetic diversity among C. cayetanensis isolates from around the world (Sulaiman et al., Reference Sulaiman, Ortega, Simpson and Kerdahi2014), and a phylogenetic analysis showed that C. cayetanensis is genetically related to members of the genus Eimeria (Fig. 2A) (Relman et al., Reference Relman, Schmidt, Gajadhar, Sogin, Cross, Yoder, Sethabutr and Echeverria1996).
However, the internal transcribed spacer (ITS) sequences in C. cayetanensis are highly variable within and between samples, and this variability does not correlate with the geographic origins of the samples (Olivier et al., Reference Olivier, van de Pas, Lepp, Yoder and Relman2001). It has been demonstrated that this ITS sequence variability occurs at the individual-genome level and approaches or exceeds the variability observed among oocysts (Riner et al., Reference Riner, Nichols, Lucas, Mullin, Cross and Lindquist2010).
No genetic polymorphism has been observed in regions of the 70 kilodalton heat shock protein (HSP70) locus characterized in a previous study (Sulaiman et al., Reference Sulaiman, Torres, Simpson, Kerdahi and Ortega2013). These results also support the lack of geographic segregation and the existence of genetically homogeneous population of C. cayetanensis parasites at this genetic locus (Sulaiman et al., Reference Sulaiman, Ortega, Simpson and Kerdahi2014).
Genome characteristics
Tracing the source of infection is facilitated by the genomic comparison of isolates. Cyclospora can also be clearly identified and differentiated from other protozoan parasites involved in foodborne or waterborne outbreaks by their genomic differences. The mitochondrial genome of C. cayetanensis is ~6200 bp in length, with 33% GC content (Cinar et al., Reference Cinar, Gopinath, Jarvis and Murphy2015; Ogedengbe et al., Reference Ogedengbe, Qvarnstrom, da Silva, Arrowood and Barta2015; Tang et al., Reference Tang, Guo, Zhang, Rowe, Roellig, Frace, Li, Liu, Feng and Xiao2015). It contains three protein-coding genes (cytb, cox1 and cox3) and 14 large subunit (LSU) and nine SSU fragmented rRNA genes (Cinar et al., Reference Cinar, Gopinath, Jarvis and Murphy2015; Ogedengbe et al., Reference Ogedengbe, Qvarnstrom, da Silva, Arrowood and Barta2015). The mitochondrial genome of C. cayetanensis has a linear concatemeric or circular mapping topology (Tang et al., Reference Tang, Guo, Zhang, Rowe, Roellig, Frace, Li, Liu, Feng and Xiao2015). A comparative genomic analysis showed strong similarity between the C. cayetanensis and E. tenella genomes, with 90.4% nucleotide sequence similarity and complete synteny in gene organization (Tang et al., Reference Tang, Guo, Zhang, Rowe, Roellig, Frace, Li, Liu, Feng and Xiao2015). Phylogenetic analyses of the mitochondrial genomic sequences have confirmed the genetic similarities between avian Eimeria spp. and C. cayetanensis (Fig. 2B).
The apicoplast genome of C. cayetanensis is ~34 000 bp in size and encodes ~65 genes, with 22% GC content (Tang et al., Reference Tang, Guo, Zhang, Rowe, Roellig, Frace, Li, Liu, Feng and Xiao2015; Liu et al., Reference Liu, Wang, Zheng, Xu, Roellig, Li, Frace, Tang, Arrowood, Moss, Zhang, Feng and Xiao2016). The apicoplast genome is circular, encodes the complete machinery for protein biosynthesis and contains two inverted repeats that differ slightly in the LSU rRNA gene sequences (Tang et al., Reference Tang, Guo, Zhang, Rowe, Roellig, Frace, Li, Liu, Feng and Xiao2015). A comparative genomic analysis revealed high-nucleotide sequence similarity (85.6%) between C. cayetanensis and E. tenella, and a phylogenetic analysis of apicoplast genomic sequences also confirmed the genetic similarities between avian Eimeria spp. and C. cayetanensis (Fig. 2C).
The whole genome of C. cayetanensis is estimated to have a total length of 44 Mbp, with 52% GC content and ~7500 gene (Liu et al., Reference Liu, Wang, Zheng, Xu, Roellig, Li, Frace, Tang, Arrowood, Moss, Zhang, Feng and Xiao2016). A comparative genomic analysis indicated that C. cayetanensis shares a coccidia-like metabolism and invasion components, but has unique surface antigens (Liu et al., Reference Liu, Wang, Zheng, Xu, Roellig, Li, Frace, Tang, Arrowood, Moss, Zhang, Feng and Xiao2016). There are also some major differences in the amino acid metabolism and the posttranslational modification of proteins between C. cayetanensis and other apicomplexans (Liu et al., Reference Liu, Wang, Zheng, Xu, Roellig, Li, Frace, Tang, Arrowood, Moss, Zhang, Feng and Xiao2016). A multilocus sequence typing tool for C. cayetanensis has been developed based on its whole genome, which involves five microsatellite loci (Guo et al., Reference Guo, Roellig, Li, Tang, Frace, Ortega, Arrowood, Feng, Qvarnstrom, Wang, Moss, Zhang and Xiao2016). Noticeable geographic clustering has been observed in human C. cayetanensis isolates from around the world (Li et al., Reference Li, Chang, Shi, Wang, Fu, Li, Xu, Jia, Guo and Zhang2017). Quantitative polymerase chain reaction (PCR) (Guo et al., Reference Guo, Wang, Wang, Zhang, Ortega and Feng2019) and PCR assays (Nascimento et al., Reference Nascimento, Barta, Whale, Hofstetter, Casillas, Barratt, Talundzic, Arrowood and Qvarnstrom2019), both targeting the polymorphic region in the mitochondrial genome, have been developed to genotype C. cayetanensis isolates. Two novel similarity-based classification algorithms for C. cayetanensis have been developed, including a Bayesian and heuristic component that infer the relatedness of pathogen isolates (Barratt et al., Reference Barratt, Park, Nascimento, Hofstetter, Plucinski, Casillas, Bradbury, Arrowood, Qvarnstrom and Talundzic2019). These useful genotyping tools should be helpful in initial source-tracking studies and in distinguishing different case clusters, especially during cyclosporiasis outbreaks.
Clinical features
The clinical symptoms of cyclosporiasis in humans typically manifest as periodic profuse watery diarrhea, together with malaise, nausea, anorexia, cramping and periods of apparent remission (Shields and Olson, Reference Shields and Olson2003). Mild-to-moderate self-limiting diarrhea is common among healthy individuals who have ingested sporulated oocysts (Mansfield and Gajadhar, Reference Mansfield and Gajadhar2004). However, patients with immune dysfunction can experience severe intestinal injury and prolonged diarrhea (Shields and Olson, Reference Shields and Olson2003; Mansfield and Gajadhar, Reference Mansfield and Gajadhar2004). In some cases, low-grade fever and the malabsorption of d-xylose may be present (Shields and Olson, Reference Shields and Olson2003). Asymptomatic infections also occur frequently in disease-endemic areas.
Striking intestinal histological changes are observed during C. cayetanensis infection, including acute or chronic inflammation, disruption of the surface epithelium, villous atrophy, crypt hyperplasia (Connor et al., Reference Connor, Shlim, Scholes, Rayburn, Reidy and Rajah1993) and intense lymphocytic infiltration within the lamina propria and epithelial cells (Ortega et al., Reference Ortega, Nagle, Gilman, Watanabe, Miyagui, Quispe, Kanagusuku, Roxas and Sterling1997; Wiwanitkit, Reference Wiwanitkit2006). The inflammatory changes associated with C. cayetanensis infection may persist beyond the eradication of the parasite (Connor et al., Reference Connor, Reidy and Soave1999). Reactive hyperaemia with vascular dilatation and congestion of the villous capillaries has also been observed (Ortega et al., Reference Ortega, Nagle, Gilman, Watanabe, Miyagui, Quispe, Kanagusuku, Roxas and Sterling1997).
In addition to gastrointestinal symptoms, C. cayetanensis can infect the biliary tract (Sifuentes-Osornio et al., Reference Sifuentes-Osornio, Porras-Cortés, Bendall, Morales-Villarreal, Reyes-Terán and Ruiz-Palacios1995), resulting in acalculous cholecystitis in people with acquired immunodeficiency syndrome (AIDS), and the presence of oocysts in gallbladder epithelial cells (Zar et al., Reference Zar, El-Bayoumi and Yungbluth2001). Although no C. cayetanensis respiratory infection has yet been identified, C. cayetanensis oocysts were detected in the sputum of two patients with tuberculosis (Di Gliullo et al., Reference Di Gliullo, Cribari, Bava, Cicconetti and Collazos2000; Hussein et al., Reference Hussein, Abdul-Manaem and El-Attary2005). Cyclospora cayetanensis infection has been associated with a variety of sequelae, including reactive arthritis syndrome, Reiter syndrome and Guillain-Barre syndrome (Connor et al., Reference Connor, Johnson and Soave2001; Shields and Olson, Reference Shields and Olson2003; Abanyie et al., Reference Abanyie, Harvey, Harris, Wiegand, Gaul, Desvignes-Kendrick, Irvin, Williams, Hall, Herwaldt, Gray, Qvarnstrom, Wise, Cantu, Cantey, Bosch, DA Silva, Fields, Bishop, Wellman, Beal, Wilson, Fiore, Tauxe, Lance, Slutsker and Parise2015).
Epidemiology
Outbreaks of human cyclosporiasis
Cyclospora cayetanensis infections in humans have been documented in over 56 countries worldwide, distributed across all five human-inhabited continents (Fig. 3; Table 1S). The first recorded outbreak of C. cayetanensis infection (called an ‘alga-like organism’ at the time) occurred among 55 British expatriates with prolonged diarrhea in Nepal between June and November, 1989 (Shlim et al., Reference Shlim, Cohen, Eaton, Rajah, Long and Ungar1991). The first reported outbreak of diarrheal illness associated with Cyclospora infection in the USA was in 1990 (Huang et al., Reference Huang, Weber, Sosin, Griffin, Long, Murphy, Kocka, Peters and Kallick1995).
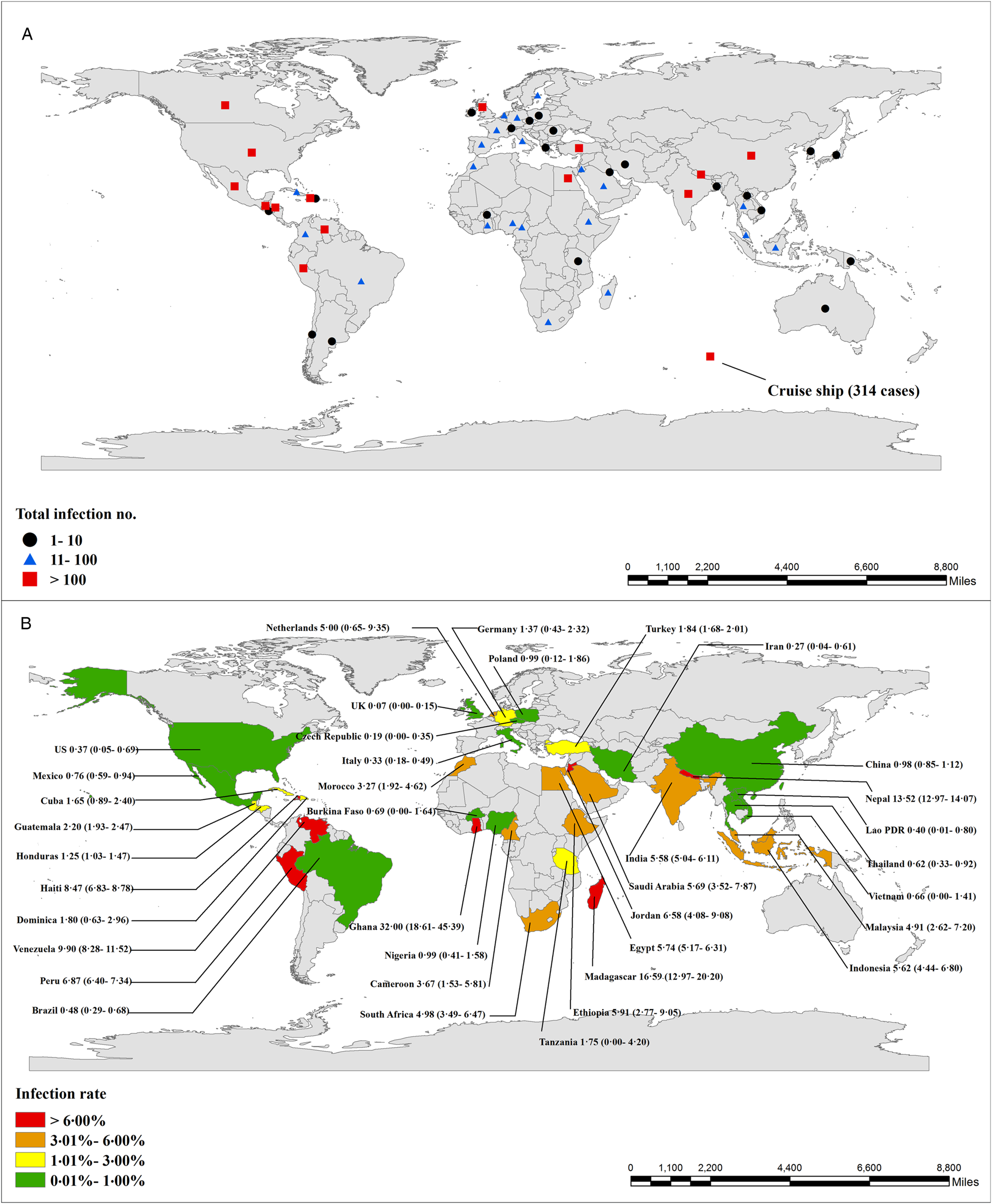
Fig. 3. Number of documented human C. cayetanensis infections and prevalence worldwide. Number of documented infection cases (A) and prevalence (B) worldwide (95% confidence intervals are shown in brackets).
Up to 1996, more than 1400 cases of cyclosporiasis were recorded in multistate outbreaks in the USA and Canada (Herwaldt and Ackers, Reference Herwaldt and Ackers1997). The most recent large outbreaks were documented in 2013 and 2018 concerning multistate outbreaks in the USA (Abanyie et al., Reference Abanyie, Harvey, Harris, Wiegand, Gaul, Desvignes-Kendrick, Irvin, Williams, Hall, Herwaldt, Gray, Qvarnstrom, Wise, Cantu, Cantey, Bosch, DA Silva, Fields, Bishop, Wellman, Beal, Wilson, Fiore, Tauxe, Lance, Slutsker and Parise2015; Casillas et al., Reference Casillas, Bennett and Straily2018). Up to December 2018, at least 13 countries documented cyclosporiasis outbreaks, involving ~6557 cases (Table 2S). Among these countries, cyclosporiasis has mainly been documented in the Americas and Europe, including Peru, Mexico, the USA, Canada and the United Kingdom (Table 2S).
Prevalence and case reports of C. cayetanensis in humans
A total of 13 845 C. cayetanensis cases have been recorded in humans, either during epidemiological studies (5478), during outbreak investigations (6557), or in case reports (1810) (Table 2S; Table 3S; Table 4S). The overall prevalence of C. cayetanensis among humans worldwide is 3.55% (5478/1 54 410). Asia (5.63%, 2771/49 254) and Africa (5.33%, 554/10 401) have shown greater prevalence than the Americas (3.03%, 1625/53 775) and Europe (1.28%, 528/41 186). A high prevalence of C. cayetanensis and large numbers of cases have been recorded in Nepal (13.68%) and India (5.58%) in Asia; Madagascar (16.59%) and Egypt (5.74%) in Africa and Venezuela (9.90%), Peru (6.87%) and Haiti (8.47%) in the Americas (Fig. 3).
Transmission risk factor assessment
A marked seasonality (rainy season or summer) has been observed in human C. cayetanensis infections in the northern hemisphere, including in China (Zhou et al., Reference Zhou, Lv, Wang, Wang, Jian, Zhang, Ning, Fu, Wang, Qi, Yao, Zhao, Zhang, Sun, Shi, Arrowood and Xiao2011; Jiang et al., Reference Jiang, Yuan, Zang, Li, Wang, Zhang, Liu, Cao and Shen2018), Nepal (Sherchand and Cross, Reference Sherchand and Cross2001; Kimura et al., Reference Kimura, Rai, Rai, Insisiengmay, Kawabata, Karanis and Uga2005; Bhandari et al., Reference Bhandari, Tandukar, Parajuli, Thapa, Chaudhary, Shrestha, Shah, Sherchan and Sherchand2015), Turkey (Ozdamar et al., Reference Ozdamar, Hakko and Turkoglu2010), Honduras (Kaminsky et al., Reference Kaminsky, Lagos, Raudales Santos and Urrutia2016) and Mexico (Orozco-Mosqueda et al., Reference Orozco-Mosqueda, Martínez-Loya and Ortega2014). The consistent pattern of the seasonal distribution of C. cayetanensis infections probably reflects the optimal environmental conditions (temperature and humidity) that are required for oocysts to sporulate. The major risk factors for Cyclospora transmission are probably the consumption of or contact with oocysts in contaminated food, water or soil; contact with animals and poor sanitation. These findings are typically documented in Peru (contaminated water sources) (Burstein Alva, Reference Burstein Alva2005), Nepal (contaminated drinking water) (Bhattachan et al., Reference Bhattachan, Sherchand, Tandukar, Dhoubhadel, Gauchan and Rai2017), Venezuela (contact with soil contaminated with human faeces) (Chacín-Bonilla et al., Reference Chacín-Bonilla, Barrios and Sanchez2007), Nepal (livestock kept near households and the consumption of raw vegetables and fruits) (Bhandari et al., Reference Bhandari, Tandukar, Parajuli, Thapa, Chaudhary, Shrestha, Shah, Sherchan and Sherchand2015) and Turkey (consumption of tap water or eating in unsanitary establishments) (Erdogan et al., Reference Erdogan, Kurt, Mandiracioglu, Ahmet, Mucide and Hande2012), among others. In summary, the epidemiological determinants and risk factors for human cyclosporiasis are shown in Table 1.
Table 1. Epidemiological determinants and risk factors for human cyclosporiasis

Susceptible populations and risk factors
Cyclospora cayetanensis is recognized as an opportunistic protozoan pathogen of humans (Wiwanitkit, Reference Wiwanitkit2006). Immunodeficiency and diarrhea in the host are two major risk factors for C. cayetanensis infection. Notable distributions of infection have been documented in Nigeria (human immunodeficiency virus (HIV) patients with diarrhea) (Alakpa et al., Reference Alakpa, Fagbenro-Beyioku and Clarke2002), Mexico (patients with diarrhea) (Jiménez-González et al., Reference Jiménez-González, Martínez-Gordillo, Caballero-Salazar, Peralta-Abarca, Cárdenas-Cardoz, Arzate-Barbosa and Ponce-Macotela2012), Honduras (patients with diarrhea or liquid stools) (Kaminsky et al., Reference Kaminsky, Lagos, Raudales Santos and Urrutia2016) and Turkey (immunosuppressed patients) (Karaman et al., Reference Karaman, Daldal, Ozer, Enginyurt and Erturk2015), among others.
The statistics for C. cayetanensis infection in different human populations demonstrate that diarrhea is a major risk factor for Cyclospora infection: immunocompromised and immunocompetent individuals with diarrhea (7.38 vs 9.14%, respectively) both had a significantly higher prevalence of infection than patients with other symptoms (4.91 vs 2.09%, respectively; P = 0.0001).
Poor sanitation conditions are another risk factor for infection with C. cayetanensis. It should be noted that people from low-income communities living in areas with poor sanitation have the highest prevalence of infection. Remarkably high-prevalence rates have been reported in Peru (54.88 and 41.58%), Venezuela (24.20%) and India (22.27%), together with poor sanitary conditions (Burstein Alva, Reference Burstein Alva2005; Nundy et al., Reference Nundy, Gilman, Xiao, Cabrera, Cama, Ortega, Kahn and Cama2011; Cazorla et al., Reference Cazorla, Acosta, Acosta and Morales2012; Jeevitha et al., Reference Jeevitha, Pushparaj and Kanchana2014). In one study in Nepal, the members of a family that kept livestock at home had higher Cyclospora infection rates than families who did not (Bhandari et al., Reference Bhandari, Tandukar, Parajuli, Thapa, Chaudhary, Shrestha, Shah, Sherchan and Sherchand2015).
Age may be another factor that affects the occurrence of cyclosporiasis in humans. Many studies have reported that children show a higher prevalence of C. cayetanensis infection than the general populations, including in Guatemala, Nepal, Turkey and Honduras, among others (Bern et al., Reference Bern, Hernandez, Lopez, Arrowood, de Mejia, de Merida, Hightower, Venczel, Herwaldt and Klein1999; Kimura et al., Reference Kimura, Rai, Rai, Insisiengmay, Kawabata, Karanis and Uga2005; Erdogan et al., Reference Erdogan, Kurt, Mandiracioglu, Ahmet, Mucide and Hande2012; Bhandari et al., Reference Bhandari, Tandukar, Parajuli, Thapa, Chaudhary, Shrestha, Shah, Sherchan and Sherchand2015; Kaminsky et al., Reference Kaminsky, Lagos, Raudales Santos and Urrutia2016). However, unexpectedly, children had a lower infection rate than the general population (4.90 vs 9.36%, respectively) of immunocompetent individuals with diarrhea, according to epidemiological statistics (P < 0.0001) (Table 2). This may be because the general population has more opportunity to consume raw produce than children.
Table 2. Cyclospora cayetanensis prevalence in different human population groups

Note: Summarized in ‘Table 3S: Epidemiology investigation of Cyclospora cayetanensis prevalence in humans’.
Cyclospora cayetanensis is also an important pathogen causing traveler's diarrhea, especially in industrialized regions (Shields and Olson, Reference Shields and Olson2003; Mansfield and Gajadhar, Reference Mansfield and Gajadhar2004). International travel or expatriate relocation to developing countries with disease-endemic areas or poor sanitation might be a risk factor for cyclosporiasis in humans (Fryauff et al., Reference Fryauff, Krippner, Prodjodipuro, Ewald, Kawengian, Pegelow, Yun, von Heydwolff-Wehnert, Oyofo and Gross1999; Pandey et al., Reference Pandey, Bodhidatta, Lewis, Murphy, Shlim, Cave, Rajah, Springer, Batchelor, Sornsakrin and Mason2011; Kłudkowska et al., Reference Kłudkowska, Pielok, Frąckowiak and Paul2017).
Animal reservoirs
Several Cyclospora species or Cyclospora-like organisms have been reported in various animals (Table 5S), including five Cyclospora species identified in primates (Eberhard et al., Reference Eberhard, da Silva, Lilley and Pieniazek1999, Reference Eberhard, Njenga, DaSilva, Owino, Nace, Won and Mwenda2001; Ortega and Sanchez, Reference Ortega and Sanchez2010; Li et al., Reference Li, Ye, Arrowood, Ma, Wang, Xu, Feng and Xiao2015). Cyclospora-like organisms have been documented in dogs, cattle, chickens, rats, house mice, birds and even shellfish. The Asian freshwater clam (Corbicula fluminea) can recover the oocysts of C. cayetanensis during artificial contamination, and could therefore be used as a biological indicator of water contaminated with oocysts (Graczyk et al., Reference Graczyk, Ortega and Conn1998).
Another study attempted to develop an animal model of C. cayetanensis in which to study human cyclosporiasis. Various types of animals (various strains of mice, rats, sand rats, chickens, ducks, rabbits, birds, hamsters, ferrets, pigs, dogs, owl monkeys, rhesus monkeys and cynomolgus monkeys) were inoculated with human C. cayetanensis oocysts by gavage. None of the animals had developed patent infection or signs of infection 4–6 weeks after inoculation. It was concluded that none of the mammals tested are susceptible to infection by C. cayetanensis (Graczyk et al., Reference Graczyk, Ortega and Conn1998). Combined with the unpublished observation and personal communication data, great efforts had been made to attempts to infect various animals, the animal models of C. cayetanensis infections were still unsuccessfully.
A pilot study sought to infect human volunteers with C. cayetanensis, but no oocysts were detected in any stool sample from any of the seven volunteers during the 16-week trial (Alfano-Sobsey et al., Reference Alfano-Sobsey, Eberhard, Seed, Weber, Won, Nace and Moe2004). These results suggest that the conditions necessary for Cyclospora to become infectious were not maintained during the preparation or storage of the oocysts. Future studies are required to assess the effects of temperature, humidity, storage conditions and disinfection on the survival, viability and infectivity of stored Cyclospora oocysts.
Food, water and soil sample contamination
In industrialized countries or regions, cyclosporiasis is most often linked to foodborne outbreaks (Rose and Slifko, Reference Rose and Slifko1999). In developing countries or disease-endemic areas, recorded C. cayetanensis infections have been associated with contact with contaminated food, water or soil (Burstein Alva, Reference Burstein Alva2005; Chacín-Bonilla, Reference Chacín-Bonilla2008; Bhandari et al., Reference Bhandari, Tandukar, Parajuli, Thapa, Chaudhary, Shrestha, Shah, Sherchan and Sherchand2015). In a community in Venezuela, a strong association between environmental contact with faecal-contaminated soil and the occurrence of cyclosporiasis was detected, suggesting that contact with soil may be an important mode of transmission (Chacín-Bonilla, Reference Chacín-Bonilla2008).
There are many records of vegetables, fruits, water and soil contaminated with Cyclospora oocysts in countries as diverse as Italy (Giangaspero et al., Reference Giangaspero, Marangi, Koehler, Papini, Normanno, Lacasella, Lonigro and Gasser2015), Malaysia (Bilung et al., Reference Bilung, Tahar, Yunos, Apun, Lim, Nillian and Hashim2017), Peru (Sturbaum et al., Reference Sturbaum, Ortega, Gilman, Sterling, Cabrera and Klein1998), Nepal (Sherchand and Cross, Reference Sherchand and Cross2001) and Vietnam (Tram et al., Reference Tram, Hoang, Cam, Chung, Fyfe, Isaac-Renton and Ong2008), among others (Table 6S). Numerous methods have been developed for the recovery and analysis of Cyclospora oocysts in contaminated food, water and soil samples (Robertson et al., Reference Robertson, Gjerde and Campbell2000; Shields et al., Reference Shields, Lee and Murphy2012).
Detection methods
A laboratory diagnosis of C. cayetanensis infection can be made simply by examining wet-mount preparations of faeces under light microscopy or by the autofluorescence of oocysts under UV epifluorescence microscopy. A more-automated flow-cytometric detection assay for C. cayetanensis in human faecal specimens was developed based on the morphology and autofluorescence characteristics of oocysts (Dixon et al., Reference Dixon, Bussey, Parrington and Parenteau2005). Modified Ziehl–Neelsen acid-fast staining is recommended for the detection of Cyclospora oocysts (Brennan et al., Reference Brennan, MacPherson, Palmer and Keystone1996; Clarke and McIntyre, Reference Clarke and McIntyre1996). Some other staining methods, such as (modified) Kinyoun acid-fast staining (Gonçalves et al., Reference Gonçalves, Uemura, Castilho and Corbett2005; Hussein, Reference Hussein2007; Behera et al., Reference Behera, Mirdha, Makharia, Bhatnagar, Dattagupta and Samantaray2008; Dillingham et al., Reference Dillingham, Pinkerton, Leger, Severe, Guerrant, Pape and Fitzgerald2009; Bhandari et al., Reference Bhandari, Tandukar, Parajuli, Thapa, Chaudhary, Shrestha, Shah, Sherchan and Sherchand2015), trichrome staining (Turgay et al., Reference Turgay, Yolasigmaz, Erdogan, Zeyrek and Uner2007; Al-Megrin, Reference Al-Megrin2010), carbol fuchsin staining (Alakpa et al., Reference Alakpa, Fagbenro-Beyioku and Clarke2002; Chacín-Bonilla et al., Reference Chacín-Bonilla, Barrios and Sanchez2007), (modified) safranin staining (Visvesvara et al., Reference Visvesvara, Moura, Kovacs-Nace, Wallace and Eberhard1997) and lactophenol cotton blue staining (Parija et al., Reference Parija, Shivaprakash and Jayakeerthi2003), have been used in the past to identify Cyclospora oocysts in faecal smears, with variable degree of sensitivity and specificity. However, these morphology-based detection methods need more parasites burden, and may lead to frequent false positive results or false negatives. There are large differences in the performance between the different microscopy techniques. Direct detection using epifluorescence is actually the very best option, followed by the safranin-stain. In practice, two or more techniques could be used together to detect the presence of parasites.
Several PCR-based detection methods that amplify specific genes of C. cayetanensis have been developed. The first PCR method used for the clinical identification of C. cayetanensis, based on SSU rRNA gene sequences, was developed by Relman et al. (Reference Relman, Schmidt, Gajadhar, Sogin, Cross, Yoder, Sethabutr and Echeverria1996). Many other different PCR assays have since been developed. The real-time PCR based on the SSU rRNA gene has been optimized to specifically detect DNA from as few as one C. cayetanensis oocyst (Varma et al., Reference Varma, Hester, Schaefer, Ware and Lindquist2003; Verweij et al., Reference Verweij, Laeijendecker, Brienen, van Lieshout and Polderman2003). Another method uses the real-time quantitative PCR with a melting curve analysis to detect, identify and differentiate C. cayetanensis from other coccidian species of concern in animal health, zoonotic diseases and food safety (Lalonde and Gajadhar, Reference Lalonde and Gajadhar2011). Several other assays have been developed based on sequences other than the SSU rRNA gene, such as a PCR-based ITS assay, which is highly sensitive in oocyst detection (Olivier et al., Reference Olivier, van de Pas, Lepp, Yoder and Relman2001; Lalonde and Gajadhar, Reference Lalonde and Gajadhar2008), and an hsp70-gene-based nested PCR protocol for the detection of C. cayetanensis, which was developed in 2013 (Sulaiman et al., Reference Sulaiman, Torres, Simpson, Kerdahi and Ortega2013). Many molecular methods have also been used to recover and detect Cyclospora oocysts in environmental water samples and agricultural products (Quintero-Betancourt et al., Reference Quintero-Betancourt, Peele and Rose2002; Steele et al., Reference Steele, Unger and Odumeru2003; Murphy et al., Reference Murphy, Cinar, Gopinath, Noe, Chatman, Miranda, Wetherington, Neal-McKinney, Pires, Sachs, Stanya, Johnson, Nascimento, Santin, Molokin, Samadpour, Janagama, Kahler, Miller and da Silva2018). Generally speaking, molecular-based detection methods can reliably detect a smaller parasites burden than other methods, even a single oocyst, and they thus overcome many of the limitations of microscopic diagnoses (Lalonde and Gajadhar, Reference Lalonde and Gajadhar2008).
Serological screening tests for Cyclospora would support epidemiological studies, and would be especially useful in the investigation of outbreaks (Ortega and Sanchez, Reference Ortega and Sanchez2010). However, no serological assays to determine human exposure to Cyclospora are yet available. Specific antibodies for the diagnosis of C. cayetanensis infection are not easily obtained, which greatly restricts immunological testing. Another serious limitation of serological assays is the lack of a laboratory culture method with which Cyclospora can be propagated in vitro (Eberhard et al., Reference Eberhard, Ortega, Hanes, Nace, Do, Robl, Won, Gavidia, Sass, Mansfield, Gozalo, Griffiths, Gilman, Sterling and Arrowood2000; Cinar et al., Reference Cinar, Gopinath, Jarvis and Murphy2015).
Treatment
Treatment with trimethoprim–sulfamethoxazole (TMP–SMX) (160 mg trimethoprim, 800 mg sulfamethoxazole) twice daily for 7–10 days is reported to be effective in curing Cyclospora infection (Hoge et al., Reference Hoge, Shlim, Ghimire, Rabold, Pandey, Walch, Rajah, Gaudio and Echeverria1995; Escobedo et al., Reference Escobedo, Almirall, Alfonso, Cimerman, Rey and Terry2009). This is also an effective therapy for Cyclospora infections in HIV patients (Pape et al., Reference Pape, Verdier, Boncy, Boncy and Johnson1994; Verdier et al., Reference Verdier, Fitzgerald, Johnson and Pape2000) and AIDS patients with biliary disease (Sifuentes-Osornio et al., Reference Sifuentes-Osornio, Porras-Cortés, Bendall, Morales-Villarreal, Reyes-Terán and Ruiz-Palacios1995). TMP–SMX (also known as co-trimoxazole) is an effective treatment, and a low recurrence rate has been reported in many studies (Hoge et al., Reference Hoge, Shlim, Ghimire, Rabold, Pandey, Walch, Rajah, Gaudio and Echeverria1995; Madico et al., Reference Madico, McDonald, Gilman, Cabrera and Sterling1997; Goldberg and Bishara, Reference Goldberg and Bishara2012).
Ciprofloxacin is less effective than TMP–SMX, but is suitable for patients who are intolerant of sulfonamide drugs (Verdier et al., Reference Verdier, Fitzgerald, Johnson and Pape2000). Successful treatment of C. cayetanensis infections with nitazoxanide has only been reported in a small number of patients (Diaz et al., Reference Diaz, Mondragon, Ramirez and Bernal2003). However, nitazoxanide is an important treatment option for patients with a sulfa allergy or for whom treatment with sulfa or ciprofloxacin has failed (Zimmer et al., Reference Zimmer, Schuetz and Franco-Paredes2007). However, norfloxacin, metronidazole, tinidazole and quinacrine have been shown to be ineffective in several studies of human cyclosporiasis (Escobedo et al., Reference Escobedo, Almirall, Alfonso, Cimerman, Rey and Terry2009).
Conclusions
Since the earliest reported cases of human Cyclospora infection in Papua New Guinea in 1979, at least 54 countries have documented C. cayetanensis infections (involving 13 845 cases) up to December 2018. Of these countries, more than 13 have recorded cyclosporiasis outbreaks (including 6557 cases). The overall C. cayetanensis prevalence in humans worldwide is 3.55% (5478/1 54 410). Cyclospora cayetanensis infections are commonly reported in developing countries with low-socioeconomic levels or disease-endemic areas, such as Madagascar, Nepal, Indonesia, Peru and Haiti, among others. However, large outbreaks have also been documented in developed countries in Europe and the Americas, and among travelers from these countries and those returning from tropical endemic areas. Among susceptible populations, the highest prevalence has been documented in immunocompetent individuals with diarrhea. The marked seasonality of C. cayetanensis infection, which occurs predominantly during the rainy season or summer, is well documented. Infection with C. cayetanensis is mainly transmitted through the ingestion of food contaminated with oocysts. Cyclospora cayetanensis or Cyclospora-like organisms have also been detected in food, water, soil and faecal material from some animals. Detection methods based on oocyst morphology, staining and molecular testing have been developed. Treatment with TMP–SMX effectively cures C. cayetanensis infection. Ciprofloxacin is less effective than TMP–SMX, but is suitable for patients who cannot tolerate co-trimoxazole.
Despite many recent advances in research, our understanding of human cyclosporiasis is hampered by several technical difficulties. It will be necessary to establish an in vitro or animal model of C. cayetanensis in the near future, in which to study human cyclosporiasis. Rapid, convenient, precise and economic detection methods for its diagnosis and genotype in humans, and effective tracing methods, must also be developed to monitor the transmission of C. cayetanensis. More importantly, the proper disposal of faeces to avoid the contamination of soil and food, boiling and filtering drinking water and improved personal hygiene will go a long way toward preventing enteric parasitic infections.
Search strategy and selection criteria
We searched PubMed, Web of Science, ScienceDirect, Wangfang and the China National Knowledge Infrastructure, with no language restriction, using the following search terms to screen for relevant articles: ‘Cyclospora’ or ‘Cyclospora-like organisms’ or ‘cyclosporiasis’ or ‘cyanobacterium-like body’ or ‘alga-like organism’. For articles without the full text or published in other languages, the titles and abstracts in English were screened for mention of Cyclospora infection. We included articles published up to 31 December 2018, when calculating the epidemiology data and summarizing the cases of infection. Articles published in English, Spanish, Portuguese, French, Turkish, Chinese, Czech, Dutch, Japanese, Rumanian and German were included.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182019001471.
Acknowledgements
We thank Janine Miller, PhD, of Liwen Bianji, Edanz Group China (www.liwenbianji.cn/ac), for editing the English text of a draft of this manuscript.
Financial support
This study was partly supported the National Key Research and Development Program of China (2017|YFD0501305, 2017YFD0500405), the National Natural Science Foundation of China (31330079, 30600603, 31672548) and the Natural Science Foundation of Henan Province (162300410129).
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical standards
Not applicable.







