









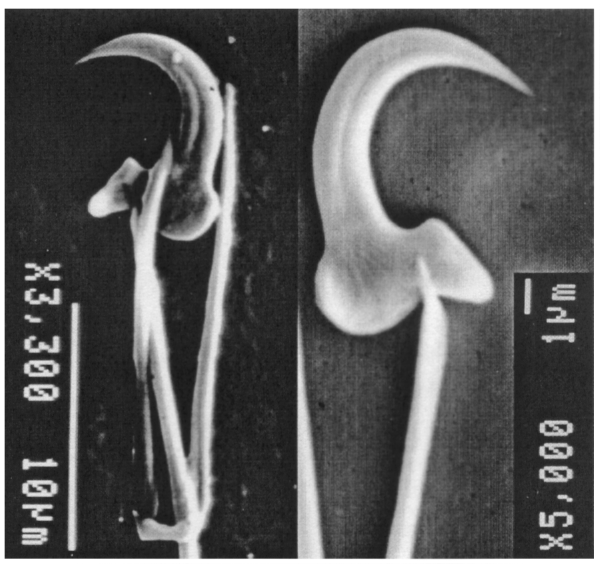










Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Hansen, Haakon
Bachmann, Lutz
and
Bakke, Tor A
2003.
Mitochondrial DNA variation of Gyrodactylus spp. (Monogenea, Gyrodactylidae) populations infecting Atlantic salmon, grayling, and rainbow trout in Norway and Sweden.
International Journal for Parasitology,
Vol. 33,
Issue. 13,
p.
1471.
Olson, Peter D.
and
Tkach, Vasyl V.
2005.
Vol. 60,
Issue. ,
p.
165.
Makhrov, A. A.
and
Bolotov, I. N.
2006.
Dispersal routes and species identification of freshwater animals in Northern Europe: A review of molecular evidence.
Russian Journal of Genetics,
Vol. 42,
Issue. 10,
p.
1101.
S. ZIĘTARA, MAREK
KUUSELA, JUSSI
and
LUMME, JAAKKO
2006.
Escape from an evolutionary dead end: a triploid clone of Gyrodactylus salaris is able to revert to sex and switch host (Platyhelminthes, Monogenea, Gyrodactylidae).
Hereditas,
Vol. 143,
Issue. 2006,
p.
84.
Bakke, T.A.
Cable, J.
and
Harris, P.D.
2007.
Advances in Parasitology Volume 64.
Vol. 64,
Issue. ,
p.
161.
Hansen, Haakon
Bakke, Tor A.
and
Bachmann, Lutz
2007.
Mitochondrial haplotype diversity of Gyrodactylus thymalli (Platyhelminthes; Monogenea): extended geographic sampling in United Kingdom, Poland, and Norway reveals further lineages.
Parasitology Research,
Vol. 100,
Issue. 6,
p.
1389.
OLSTAD, K.
ROBERTSEN, G.
BACHMANN, L.
and
BAKKE, T. A.
2007.
Variation in host preference within Gyrodactylus salaris (Monogenea): an experimental approach.
Parasitology,
Vol. 134,
Issue. 4,
p.
589.
Collins, Catherine M.
Olstad, Kjetil
Sterud, Erik
Jones, Catherine S.
Noble, Leslie R.
Mo, Tor Atle
and
Cunningham, Carey O.
2007.
Isolation of a novel fish thymidylate kinase gene, upregulated in Atlantic salmon (Salmo salar L.) following infection with the monogenean parasite Gyrodactylus salaris.
Fish & Shellfish Immunology,
Vol. 23,
Issue. 4,
p.
793.
Heinecke, R D
Martinussen, T
and
Buchmann, K
2007.
Microhabitat selection of Gyrodactylus salaris Malmberg on different salmonids.
Journal of Fish Diseases,
Vol. 30,
Issue. 12,
p.
733.
Ziętara, Marek S.
Kuusela, Jussi
Veselov, Alexei
and
Lumme, Jaakko
2007.
Molecular faunistics of accidental infections of Gyrodactylus Nordmann, 1832 (Monogenea) parasitic on salmon Salmo salar L. and brown trout Salmo trutta L. in NW Russia.
Systematic Parasitology,
Vol. 69,
Issue. 2,
p.
123.
Kania, P W
Jørgensen, T R
and
Buchmann, K
2007.
Differentiation between a pathogenic and a non‐pathogenic form of Gyrodactylus salaris using PCR‐RFLP.
Journal of Fish Diseases,
Vol. 30,
Issue. 2,
p.
123.
Von Gersdorff Jørgensen, L
Heinecke, R D
Kania, P
and
Buchmann, K
2008.
Occurrence of gyrodactylids on wild Atlantic salmon, Salmo salar L., in Danish rivers.
Journal of Fish Diseases,
Vol. 31,
Issue. 2,
p.
127.
Jørgensen, Thomas R.
Larsen, Thomas B.
and
Buchmann, Kurt
2009.
Parasite infections in recirculated rainbow trout (Oncorhynchus mykiss) farms.
Aquaculture,
Vol. 289,
Issue. 1-2,
p.
91.
Jørgensen, Thomas R.
Raida, Martin K.
Kania, Per W.
and
Buchmann, Kurt
2009.
Response of rainbow trout (Oncorhynchus mykiss) in skin and fin tissue during infection with a variant of Gyrodactylus salaris (Monogenea: Gyrodactylidae).
Folia Parasitologica,
Vol. 56,
Issue. 4,
p.
251.
Harris, Phil D.
Bachmann, Lutz
and
Bakke, Tor A.
2010.
Atlantic Salmon Ecology.
p.
221.
Shinn, A.P.
Collins, C.
García-Vásquez, A.
Snow, M.
Matějusová, I.
Paladini, G.
Longshaw, M.
Lindenstrøm, T.
Stone, D.M.
Turnbull, J.F.
Picon-Camacho, S.M.
Rivera, C. Vázquez
Duguid, R.A.
Mo, T.A.
Hansen, H.
Olstad, K.
Cable, J.
Harris, P.D.
Kerr, R.
Graham, D.
Monaghan, S.J.
Yoon, G.H.
Buchmann, K.
Taylor, N.G.H.
Bakke, T.A.
Raynard, R.
Irving, S.
and
Bron, J.E.
2010.
Multi-centre testing and validation of current protocols for the identification of Gyrodactylus salaris (Monogenea).
International Journal for Parasitology,
Vol. 40,
Issue. 12,
p.
1455.
Heneberg, Petr
and
Literák, Ivan
2013.
Molecular phylogenetic characterization of Collyriclum faba with reference to its three host-specific ecotypes.
Parasitology International,
Vol. 62,
Issue. 3,
p.
262.
Firdaus-Nawi, Mohd
Zamri-Saad, Mohd
Nik-Haiha, Nik Yusoff
Zuki, Md Abu Bakar
and
Effendy, Abd Wahid Mohd
2013.
Histological assessments of intestinal immuno-morphology of tiger grouper juvenile, Epinephelus fuscoguttatus
.
SpringerPlus,
Vol. 2,
Issue. 1,
Rakers, Sebastian
Niklasson, Lars
Steinhagen, Dieter
Kruse, Charli
Schauber, Jürgen
Sundell, Kristina
and
Paus, Ralf
2013.
Antimicrobial Peptides (AMPs) from Fish Epidermis: Perspectives for Investigative Dermatology.
Journal of Investigative Dermatology,
Vol. 133,
Issue. 5,
p.
1140.
Ramírez, Raúl
Bakke, Tor A.
and
Harris, Philip D.
2014.
Same barcode, different biology: differential patterns of infectivity, specificity and pathogenicity in two almost identical parasite strains.
International Journal for Parasitology,
Vol. 44,
Issue. 8,
p.
543.