


Introduction
A large number of Tremella s. lat. species grow on lichens (Diederich Reference Diederich1986, Reference Diederich1996, Reference Diederich2003, Reference Diederich2007; Diederich & Marson Reference Diederich and Marson1988; Diederich & Christiansen Reference Diederich and Christiansen1994; Sérusiaux et al. Reference Sérusiaux, Diederich, Ertz and van den Boom2003; Zamora et al. Reference Zamora, Pérez-Ortega and Rico2011, 2016; Millanes et al. Reference Millanes, Westberg, Wedin and Diederich2012, Reference Millanes, Diederich, Westberg, Knutsson and Wedin2014a ; Diederich et al. Reference Diederich, Millanes and Wedin2014). These taxa are, just like many other Tremella species, mycoparasites, and no interactions with the host lichen photobiont algae have been observed (Grube & de los Ríos Reference Grube and de los Ríos2001). They often induce the production of galls or deformations on the host thallus or apothecia, although in some species basidiomata develop in the absence of gall-like structures (Diederich Reference Diederich1996; Zamora et al. Reference Zamora, Pérez-Ortega and Rico2011). Diederich (Reference Diederich1996) described the majority of the known lichen-inhabiting species in his monograph of the lichenicolous heterobasidiomycetes, and his study raised the interest of lichenologists and mycologists in these previously much neglected fungi. As a result, the known diversity in the group has increased notably in recent decades, and more taxa will surely be described in the future (Millanes et al. Reference Millanes, Westberg, Wedin and Diederich2012; Zamora et al. Reference Zamora, Millanes, Wedin, Rico and Pérez-Ortegain press).
Species delimitation is often difficult in lichenicolous representatives of Tremella, due to the paucity of morphological characters, but in most cases, the host selection has proved to be a good indicator of species boundaries in this generally very host-specific group of fungi (Diederich Reference Diederich1996; Millanes et al. Reference Millanes, Westberg, Wedin and Diederich2012; Zamora et al. Reference Zamora, Millanes, Wedin, Rico and Pérez-Ortegain press). Tremella cetrariicola is a widespread species and its hitherto known host range includes Cetrariella delisei, Tuckermannopsis americana, T. chlorophylla and T. ciliaris (Diederich Reference Diederich1996). Several additional collections of Tremella on Cetrariella delisei have been made in Finland, Greenland, Norway, Russia, Svalbard and Sweden. Both the morphology and the host selection indicate that these specimens constitute a distinct species, which is described below. We will further assess the phylogenetic relationships of the new taxon with Tremella cetrariicola and with other Tremella species.
Material and Methods
Morphological studies
Herbarium specimens are deposited in OULU, S, TUR, UGDA and UPS, and in the private collection of P. Diederich. External morphology of herbarium specimens was examined and measured using an Olympus SZX16 or a Leica MZ 7.5 dissecting microscope. Macroscopic photographs were either taken using an Olympus DP11 camera on an Olympus SZX16 dissecting microscope, or using a Canon 40D camera with a Nikon BD Plan ×10 microscope objective, StackShot (Cognisys) and Helicon Focus (HeliconSoft) for increasing the depth of field. Microscopical structures were studied using hand-cut sections stained with Phloxin (1% in water) after pretreatment with KOH (5%), following the methods of Diederich (Reference Diederich1996), and observed with an Olympus CX40 microscope or a Leica DMLB. Drawings were performed by direct observation. Microscopic photographs were prepared using an Olympus BX53 microscope fitted with differential interference contrast (DIC), and an Olympus DP11 camera; or a Leica DMLB microscope with DIC, using a Leica EC3 camera, and Helicon Focus. The apiculus was not included in basidiospore measurements. Basidiospore length/width ratio is expressed as Q. Sizes in parentheses represent minimum and maximum observed values. When the number of observations is less than 50, it is indicated within parentheses.
Molecular studies
Choice of additional taxa and outgroup
In addition to the eight specimens studied, 23 specimens representing one Biatoropsis, one Cuniculitrema, 15 Tremella and one Trimorphomyces species were included in the molecular study (Table 1). The additional sampling included five specimens of Tremella cetrariicola for comparison, the type of the genus Tremella (T. mesenterica), terminals of the Aurantia, Indecorata and Fuciformis groups distinguished by Chen (Reference Chen1998), terminals representing three groups of lichenicolous species distinguished by Millanes et al. (Reference Millanes, Diederich, Ekman and Wedin2011), and finally two teleomorphic species within the Tremellales that grouped together with lichenicolous taxa, although without support, in Millanes et al. (Reference Millanes, Diederich, Ekman and Wedin2011), viz. Cuniculitrema polymorpha and Trimorphomyces papilionaceus. Syzygospora effibulata was used as outgroup.
Table 1 GenBank accession numbers for the species used in this study with the outgroup Syzygospora effibulata. Newly obtained sequences are in bold

Species names, voucher information, and GenBank accession numbers are given in Table 1.
DNA extraction and amplification
DNA was extracted directly from the specimens examined (Table 1). The outer surface of the selected galls, in which most of the tremellalean hyphae and hymenial components are located, was sectioned and separated with a scalpel in order to minimize the lichen tissue in the DNA extraction. Approximately three or four basidiomata were selected from each specimen for DNA extraction. Total DNA was extracted using the Qiagen DNeasy Plant MiniKit, according to the manufacturer’s instructions, but using 50 μl of water in each of the last two steps of final elution.
For PCR amplification, we used general fungal primers in combination with primers designed to selectively amplify the DNA from tremellalean fungi (Millanes et al. Reference Millanes, Diederich, Ekman and Wedin2011). The primers ITS1F (Gardes & Bruns Reference Gardes and Bruns1993), BasidLSU3-3 and BasidLSU1-5 (Millanes et al. Reference Millanes, Diederich, Ekman and Wedin2011), and LR5 (Vilgalys & Hester Reference Vilgalys and Hester1990) were used to amplify the internal transcribed spacer I, the 5.8 rDNA gene, the internal transcribed spacer II and a fragment of c. 1000 bp in the nLSU rDNA gene.
PCR amplifications were performed using Illustra™ Hot Start PCR beads, according to the manufacturer’s instructions, with the following settings: for the primer pair ITS1F/BasidLSU3-3, we used initial denaturing at 95°C for 3 min, four cycles (95°C for 40 s, 53°C for 40 s and 72°C for 90 s), four cycles (95°C for 30 s, 50°C for 30 s and 72°C for 90 s), and finally 32 cycles (95°C for 30 s, 47°C for 30 s and 72°C for 90 s) with a final extension at 72°C for 480 s. For the primer pair BasidLSU1-5/LR5, we used initial denaturing at 95°C for 3 min, four cycles (95°C for 40 s, 56°C for 40 s and 72°C for 90 s), four cycles (95°C for 30 s, 53°C for 30 s and 72°C for 90 s) and finally 32 cycles (95°C for 30 s, 50°C for 30 s and 72°C for 90 s) with a final extension at 72°C for 420 s. Before sequencing, the PCR products were purified using the PCR-M® Clean-up System of Viogene or the enzymatic method Exo-sap-IT© provided by USB Corporation. In all PCR reactions, we used 2 μl of DNA extraction, and 0·5 μl of each primer (primer concentration 10 μM).
The PCR-products were sequenced using the DYEnamic ET terminator cycle sequencing kit (Amersham Biosciences, Freiburg, Germany), with the following settings: 25 cycles (95°C for 20 s, 50°C for 15 s, and 60°C for 60 s). Post-reaction clean-up was carried out following DYEnamic ET terminator cycle sequencing kit protocols (Amersham Biosciences). The purified samples were run on an automated sequencer (ABI Prism 377) located in the Molecular Systematic Laboratory at the Swedish Museum of Natural History, or on an automated sequencer (ABI Prism 3100-Avant) in the Genomic Unit at Rey Juan Carlos University.
Sequence alignment and phylogenetic analyses
Sequences were aligned using the Q-INS-i algorithm (Katoh & Toh Reference Katoh and Toh2008a ) of the multiple sequence alignment software MAFFT version 6.611 (Katoh et al. Reference Katoh, Misawa, Kuma and Miyata2002; Katoh & Toh Reference Katoh and Toh2008b ). Major insertions and ambiguous regions were identified and eliminated with Gblocks version 0.91b (Castresana Reference Castresana2000).
Dataset congruence was assessed manually by analyzing the datasets separately by maximum likelihood bootstrapping. Conflict among clades was considered as significant if a significantly supported clade (bootstrap support ≥ 70%; Hillis & Bull Reference Hillis and Bull1993) for one marker was contradicted with significant support by another. No incongruence was found and the data were concatenated into a single dataset.
Bayesian analyses were performed by Markov chain Monte Carlo (MCMC) sampling as implemented in the software MrBayes 3.2.4 (Ronquist et al. Reference Ronquist, Teslenko, van der Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012). We selected substitution models for each of the four gene regions (i.e., ITS1, 5.8S, ITS2, and nLSU of the nuclear ribosomal DNA), using the Akaike Information Criterion (AIC) as implemented in jModeltest (Guindon & Gascuel Reference Guindon and Gascuel2003; Posada Reference Posada2008). We used full likelihood optimization, six discrete gamma categories, and selected only among the 24 models implemented in MrBayes. A SYM+Γ model was selected for the ITS1, a SYM+I+Γ for the 5.8S and ITS2, and finally a GTR+I for the nuclear LSU rDNA. Analyses settings were as those described in Millanes et al. (Reference Millanes, Westberg, Wedin and Diederich2012), but using three runs for the MCMC search. Maximum likelihood analyses were performed in RAxMLGUI 1.3 (Silvestro & Michalak Reference Silvestro and Michalak2012), a graphical front-end for RAxML (Stamatakis Reference Stamatakis2006), using the GTRGAMMAI model of nucleotide substitution applied to all partitions. We performed a thorough ML search with a total of 100 runs and assessed node support by thorough bootstrap using 1000 bootstrap pseudoreplicates.
Results
Phylogenetic results
We generated 21 new sequences (11 ITS and 10 nLSU rDNA), which were aligned together with sequences already available in GenBank (Table 1). Two data matrices were produced, one including ITS and one including nLSU rDNA.
The combined matrix contained 1306 characters (ITS1: 1–104; 5.8S: 105–261; ITS2: 262–375; nLSU: 376–1306). The best tree obtained from the ML analysis had an ln-likelihood value of −7560·0728. The Bayesian analysis halted after 400000 generations, at which time the average standard deviation of split frequencies across runs was 0·009, which indicates that the three runs had converged (<0·01). A majority-rule consensus tree was constructed from the 6000 trees of the stationary tree sample. Since the topologies of the ML and the Bayesian trees were congruent, only the 50% majority-rule consensus tree from the Bayesian analysis is shown in Fig. 1.

Fig. 1 50% majority-rule Bayesian consensus tree with average branch lengths from the combined analyses of ITS and nLSU datasets. Bayesian PP values≥0·95 are indicated above the branches, and maximum likelihood bootstrap values≥70% below the branches. New species are indicated in bold font and the type specimen is indicated with (T). Branch lengths are scaled to the expected number of nucleotide substitutions per site.
The eight specimens of T. cetrariellae formed a single clade supported by both ML bootstrap (99%) and Bayesian posterior probabilities (1·0). This clade is divided into two subclades, which are also strongly supported by both methods: ML bootstrap (100%) and Bayesian posterior probabilities (1·0) (Fig. 1). Tremella cetrariellae forms a sister clade to Tremella cetrariicola, and this relationship is supported by both ML bootstrap (72%) and Bayesian posterior probabilities (1·0). Otherwise, the topology of the tree is congruent with previous phylogenies including similar taxon sampling (Millanes et al. Reference Millanes, Diederich, Ekman and Wedin2011, Reference Millanes, Westberg, Wedin and Diederich2012).
The Species
Tremella cetrariellae Millanes, Diederich, M. Westb., Pippola & Wedin sp. nov.
MycoBank No.: MB 814481
Lichenicolous on Cetrariella delisei, inducing pale to dark brown or black, convex, often tuberculate galls, 0·1–1·5 mm diam., on the thallus surface. Basidia 2-, 3- or 4-celled, with transverse, longitudinal, or oblique septa. Basidiospores ellipsoid to subglobose, 4–8×4–8 μm, Q=0·83–1·50. The species is phylogenetically closely related to Tremella cetrariicola, from which the new species differs by its predominantly tuberculate galls, longer, often transversely septate basidia, and typical tremelloid spores.
Type: Norway, Finnmark, Sør Varanger, Balgami, area with siliceous rocks, Empetrum and Betula nana, next to the sea and to a small lake, 69°58·757'N, 29°34·883'E, alt. 21 m, 4 July 2014, A. Millanes 1130 (S F264653––holotype).
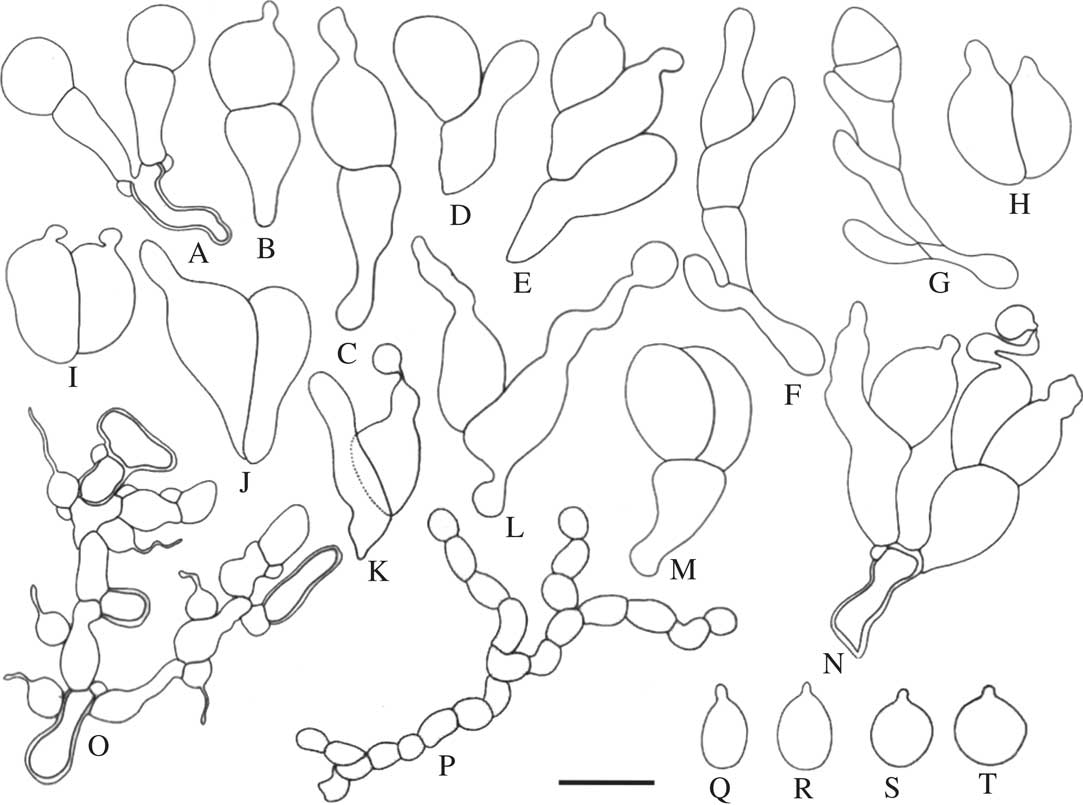
Fig. 2 Tremella cetrariellae. A–N, basidia; O, hyphae with clamps and haustorial branches; P, catenulate conidia; Q–T, basidiospores (A–E, H–M, O–S, holotype; N, T & S, F264651; F & G, OULU L002848). Scale=10 μm.

Fig. 3 Tremella cetrariellae. A–F, morphological variation in gall morphology (A, holotype; B, S F264669; C & D, OULU L002848; E, S F264651; F, S F264652); G–M, basidia showing different septation patterns; N, haustorial cells; O, catenulate conidia; P–S, basidiospores (G–J, N–R, holotype; K–M, OULU L002848; S, S F264651). Scales: A–F=0·5 mm; G–O=10 μm; P–S=5 μm. In colour online.
Sexual morph: Basidiomata superficial, waxy gelatinous, inducing the formation of galls on the thallus surface (Fig. 3A–F). Galls pale to dark brown or black, at first regularly convex to subglobose, tuberculate when mature, 0·1–1·5 mm diam., very rarely with a central depression. Context hyphae thin- to thick-walled, often with clamp connections, 1·5–4·0 μm diam. (Fig. 2O); haustorial branches frequent, mother cells spherical to subspherical, (2·5–)3·0–4·0×3·0–4·0(–5·0) μm, haustorial filament 1 μm diam., up to 15 μm long (Figs 2O & 3N). Hymenium hyaline, containing numerous probasidia; probasidial initials clavate, proliferations occurring through the basal clamp (Figs 2A, 2N, 3J). Basidia, when mature, 2-, 3-, or 4-celled, with transverse, longitudinal, or oblique septa (Figs 2A–L, 3G–K); a combination of transverse and longitudinal or oblique septa can also be observed in the same basidium (Figs 2M & N, 3L & M); transversely, obliquely and longitudinally septate basidia are often found within the same gall; when transversely septate, constricted at the septum, the lower cell with an attenuated stalk-like base, often longer than the upper cell, 10–31(–33)×4–11 μm (incl. stalk-like base; excl. epibasidia), lower part of the stalk-like base 2–4 μm diam.; when longitudinally or obliquely septate, (8–)9–13×11–18 μm; epibasidia subcylindrical, up to 28 μm long, (1–)2–5 μm diam. (Figs 2B–L, 2N, 3H–I, 3K–M). Basidiospores ellipsoid to subglobose, (3–)4–8×(3–)4–8 μm, Q=0·83–1·50(–1·60) (n=44) with a distinct apiculus, c. 1 μm diam. (Figs 2Q–T, 3P–S).
Asexual morph: Conidia catenulate, initially hyaline, later becoming brownish, individual conidia (2–)3–8×3–10 μm have been observed within the basidioma (Figs 2P, 3O).
Etymology. In reference to the host, Cetrariella delisei.
Host. Cetrariella delisei (thallus).
Distribution. Known from Finland, Greenland, Norway, Russia, Svalbard and Sweden.
Additional specimens examined (all on Cetrariella delisei). Finland: Inarin Lappi: c. 8 km NW of Viitaniemi, 5 ix 1936, S. Ahlner (S F264686). Enontekiön Lappi: Enontekiö, Kilpisjärvi, 23 vii 1948, A. J. Huuskonen, Lichenotheca Fennica 692 (OULU L002848, TUR 6655). Perä-Pohjanmaa: Simo, Kivalot, Ala-Penikka, 1988, P. Halonen 43 (OULU F073604). Oulun Pohjanmaa: Ylikiiminki, Kontionsuo, 23 vii 2007, T. Rämä (OULU 0036202).—Greenland: Qeqqata: Kangerluqssuaq, Husvika, 22 viii 1931, J. K. Tornrøe (S F264655). Ilulissat: Disko Bay, close to the Arctic Station, 29 ix 1927, M. P. Porsild (S F264660).—Norway: Finnmark: Båtsfjord, Hill W of Adamsvann, 70°32·88'N, 29°26·33'E, 2014, A. Millanes 1108 (S F264667); Nesseby municipality, 11 km SW the village Nesseby in the Varanger Peninsula, between the villages Karlebotn and Reppen, 70°06·146'N, 28°37·183'E, 4 vii 2014, M. Westberg (S F264652); Sør Varanger, Balgami, 69°58·757'N, 29°34·883'E, 4 vii 2014, G. Amo (S F264668).—Russia: Murmansk: Khibiny Mts., SE slope of Lovchor Mt., 4·5 km SE of Kirovsk, 9 viii 2000, M. Kukwa (UGDA).—Svalbard: Blomstrandhalvøya, 23 vii 2010, D. Fontaneto (S F178310).—Sweden: Jämtland: Åre par., Storlien, 30 vii 1909, E. P. Vrang (UPS). Torne Lappmark: Jukkasjärvi par., in the valley Gearggevággi (Kärkevagge) between Vásseèohkka (Vassitjåkko) and Gearggeèorru (Kärketjårro), c. 10 km ESE of Riksgränsen, 5 viii 2013, M. Westberg & U. Arup (S F264651); Jukkasjärvi par., Abisko, 68°25·94'N, 18°31·21'E, 2013, A. Millanes 874 (S F264669).
Discussion
Our morphological and phylogenetic results confirm that Tremella cetrariellae is a distinct species, different from Tremella cetrariicola. Firstly, the galls of the new species are predominantly tuberculate when mature (Fig. 3A–E) and the central depression often present in Tremella cetrariicola (Diederich Reference Diederich1996; this study, Fig. 4A) is only rarely, if at all, observed in T. cetrariellae (Fig. 3F). Secondly, the basidia of the new species are longer than those of Tremella cetrariicola, which reach a maximum 22 μm in length. Basidia of Tremella cetrariellae can be 2–4-celled, with septa that can be longitudinal, oblique or transverse (Figs 2A–N, 3G–M), whereas basidia of Tremella cetrariicola are predominantly 2-celled, and only rarely show transverse septa. The combination of longitudinal, oblique and transverse septa in the same basidia (Figs 2M & N, 3L & M) has not been observed in Tremella cetrariicola. One of the main diagnostic characters of T. cetrariicola mentioned by Diederich (Reference Diederich1996) was the somewhat limoniform spores, with a refractive apiculus situated at one of the narrow ends of the spore (Fig. 4B & C). This contrasts with most other Tremella species, in which ellipsoid to subspherical spores usually present a lateral apiculus. Figure 2S & T and Fig. 3R & S represent Tremella cetrariellae with typical Tremella spores with a lateral apiculus, while spores in Figs 2Q & R and 3P & Q are similar to those of T. cetrariicola. The presence of typical Tremella spores is thus characteristic of the new species, whereas the presence of limoniform spores, strongly developed in T. cetrariicola and occasionally in T. cetrariellae, characterizes the clade formed by both species. The presence of catenulate conidia observed in Tremella cetrariellae (Figs 2P, 3O) has never been reported in T. cetrariicola. Finally, the new species is confined to Cetrariella delisei. Material growing on this host had been included in the description of Tremella cetrariicola (Diederich Reference Diederich1996), and Urbanavichus et al. (Reference Urbanavichus, Motiejūnaitė, Kukwa and Urbanavichene2007) later reported an additional specimen from Russia. After our further studies, we can now confirm that this material must also be included within T. cetrariellae.

Fig. 4 Tremella cetrariicola for comparison (isotype—hb. Diederich). A, central depression typically observed in many galls; B & C, ellipsoid spores with a refractive basal apiculus. Scales: A=0·5 mm; B & C=5 μm. In colour online.
Our topology suggests that Tremella cetrariellae might contain two cryptic species (subclades 1 and 2, Fig. 1). With the data at hand, however, it is not possible to distinguish the subclades based on morphological or ecological data, and sampling within subclade 1 is very scant. For the time being, we consider the entire material to belong to a single species, although the infraspecific genetic variation is higher than that of its sister species Tremella cetrariicola (Fig. 1).
Millanes et al. (Reference Millanes, Diederich, Ekman and Wedin2011) showed that most Tremella species growing on Parmeliaceae grouped together, although this relationship was recovered without strong support. In our analyses, the new species is included in a monophyletic clade that only includes other species growing on Parmeliaceae. Tremella cetrariicola is the sister taxon of T. cetrariellae. The hosts of both species belong to the cetrarioid core group of the Parmeliaceae, but whereas Tuckermannopsis species are placed in the ‘Nephromopsis’ clade, Cetrariella delisei belongs to the ‘Cetraria’ clade (Nelsen et al. Reference Nelsen, Chavez, Sackett-Hermann, Thell, Randlane, Divakar, Rico and Lumbsch2011). Moreover, the host of the new species, Cetrariella delisei, contains gyrophoric and hiascic acids, which are absent in Tuckermannopsis species (Kärnefelt et al. Reference Kärnefelt, Mattson and Thell1993). It has been shown that the occurrence of lichenicolous fungi can be influenced by the chemistry of their lichenized hosts (Lawrey Reference Lawrey1995; Merinero et al. Reference Merinero, Bidussi and Gauslaa2015). This would support a scenario in which the separation of T. cetrariicola and T. cetrariellae would have occurred through a host switch, as has been shown to occur in other lichenicolous tremellalean species (Millanes et al. Reference Millanes, Truong, Westberg, Diederich and Wedin2014b ), followed by an adaptation to a different chemical environment. There are, moreover, other factors that could have hindered genetic flow and therefore favoured the separation of the two species, after a hypothetical host switch. These are the different geographical distribution of the hosts (arctoalpine in Cetrariella delisei, while more widespread in Tuckermannopsis) and the different habitat (epigeic in Cetrariella delisei vs. epiphytic in Tuckermannopsis). Further studies are needed, however, to assess hypotheses related to the mode of speciation, but these are beyond the scope of this work.
We thank Guillermo Amo, Ulf Arup, Diego Fontaneto, and Martin Kukwa for collecting material for the present study, and the staff of the Molecular Systematic Laboratory at the Swedish Museum of Natural History for excellent technical support. Marcos Méndez provided much-appreciated field guidance in Torne Lappmark. This paper was financially supported by The Swedish Taxonomy Initiative (Svenska Artprojektet, administered by the Swedish Species Information Centre/ArtDatabanken) through a grant to M. Wedin, and by the Spanish Ministry of Economy and Competitiveness (CGL2012–40123) through a grant to A. Millanes and M. Wedin. Further support for fieldwork was received from the Finnish Ministry of Environment and from the Swedish Taxonomy Initiative, through grants to E. Pippola and M. Westberg, respectively.





