



Introduction
The name Lobothallia was introduced as a subgenus of Aspicilia (Clauzade & Roux Reference Clauzade and Roux1984) and was later raised to the genus rank within Aspiciliaceae (Hafellner Reference Hafellner1991). Initially Lobothallia included taxa from the Aspicilia radiosa group and comprised four species with marginal lobes: L. alphoplaca, L. melanaspis, L. praeradiosa and L. radiosa. However, subsequent molecular studies broadened the concept of the genus to also include non-lobate crustose species (Nordin et al. Reference Nordin, Savić and Tibell2010; Kou et al. Reference Kou, Li and Ren2013) as proposed by Esnault (Reference Esnault1985) using morphological and anatomical characters.
Lobothallia is now recognized within Megasporaceae (Schmitt et al. Reference Schmitt, Yamamoto and Lumbsch2006; Nordin et al. Reference Nordin, Savić and Tibell2010) and is characterized by relatively small ascospores, rarely exceeding 18 μm in length, and short conidia, 3–8 μm long. Additional morpho-anatomical characters are: a low hymenium not exceeding 100 μm; an epihymenium lacking, or with small amounts, of Caesiocinerea-green (Meyer & Printzen Reference Meyer and Printzen2000) and consequently a reaction with N that is negative to slightly greenish (vs. strong N+ (emerald green) reaction in other Megasporaceae); a distinct sub-hypothecial algal layer (Ryan Reference Ryan, Nash, Ryan, Diederich, Gries and Bungartz2004; Kou et al. Reference Kou, Li and Ren2013) visible after staining with lactophenol blue (Clauzade & Roux Reference Clauzade and Roux1985) but which may be poorly visible unless stained (Roux et al. Reference Roux, Nordin and Bertrand2016). Species of Circinaria and Megaspora with short conidia differ from Lobothallia in having larger spores, usually more than 20 μm in length. Species of the Aspicilia polychroma group (Aspicilia candida (Anzi) Hue, A. polychroma Anzi, A. polychromoides (J. Steiner) Hue and A. thjanschanica Oxner), with comparable spore length, have much longer conidia and contain substictic acid, which is currently not known in Lobothallia (Nordin et al. Reference Nordin, Savić and Tibell2010). Only Aspicilia brucei Owe-Larss. & A. Nordin, from the A. cinerea group, has spores and conidia which are as small as those in Lobothallia (Owe-Larsson et al. Reference Owe-Larsson, Nordin, Tibell, Nash, Gries and Bungartz2007; Roux et al. Reference Roux, Nordin, Tibell and Sohrabi2011).
The genus currently comprises 13 accepted species: Lobothallia alphoplaca (Wahlenb.) Hafellner, L. cernohorskyana (Clauzade & Vězda) A. Nordin et al., L. chadefaudiana (Cl. Roux) A. Nordin et al., L. cheresina (Müll. Arg.) A. Nordin et al., L. controversa Cl. Roux & A. Nordin, L. crassimarginata X. R. Kou & Q. Ren, L. helanensis X. R. Kou & Q. Ren, L. hydrocharis (Poelt & Nimis) Sohrabi & Nimis, L. melanaspis (Ach.) Hafellner, L. praeradiosa (Nyl.) Hafellner, L. pruinosa X. R. Kou & Q. Ren, L. radiosa (Hoffm.) Hafellner and L. recedens (Taylor) A. Nordin et al. (Nordin et al. Reference Nordin, Savić and Tibell2010; Roux Reference Roux2012; Kou et al. Reference Kou, Li and Ren2013; Nimis Reference Nimis2016; Roux et al. Reference Roux, Nordin and Bertrand2016). Some taxa (viz. Lobothallia cheresina (Müll. Arg.) A. Nordin et al. chemotype justii, L. cheresina (Müll. Arg.) A. Nordin et al. chemotype microspora, Lobothallia parasitica (B. de Lesd.) ined., and Lobothallia radiosa (Hoffm.) Hafellner chemotype subcircinata) are regarded as only chemotypes (Roux Reference Roux2012; Roux et al. Reference Roux, Monnat, Gonnet, Gonnet, Poumarat, Esnault, Bertrand, Gardiennet, Masson and Bauvet2017). Further species, which probably belong in Lobothallia from western North America, along with their validly published combinations, are likely to be forthcoming (Wheeler Reference Wheeler2017).
During the study of lichen material collected by the authors in the Astrakhan, the Orenburg region of Russia, Altai (Russian and Chinese parts) and Kazakhstan, together with herbarium specimens, we recognized that some samples belong to previously undescribed species and that four taxa known as Aspicilia and Lecanora should be placed correctly in Lobothallia. Here we describe three species new to science and propose new combinations in Lobothallia.
Materials and Methods
Specimens and phenotype studies
The core material for this study was collected by the authors and deposited in herbaria ALTB, UFU and UPS. Additionally, type specimens were examined in G, H-Nyl, KW, LE, MARSSJ, S and W. Morphological observations were made using a dissecting microscope. Cross-sections of apothecia and thalli were cut by hand with a razor blade and observed after mounting in water, 10% potassium hydroxide KOH (K), 10% water solution of nitric acid HNO3 (N), calcium hypochlorite (C) and iodine solutions (I). Measurements of spores and conidia are presented as follows: (smallest value recorded) ( $\bar{x}$ − SE) –
$\bar{x}$ − SE) –  $\bar{x}$ – (
$\bar{x}$ – ( $\bar{x}$ + SE) (largest value recorded), where
$\bar{x}$ + SE) (largest value recorded), where  $\bar{x}$ is the (arithmetic) sample mean, and SE the sample error of the mean. The measurements were made with a precision of 0·5 μm.
$\bar{x}$ is the (arithmetic) sample mean, and SE the sample error of the mean. The measurements were made with a precision of 0·5 μm.
Secondary products were analyzed by applying standard thin-layer chromatography techniques (Culberson & Kristinsson Reference Culberson and Kristinsson1970). Solvent systems A (toluene: 1,4-dioxane: acetic acid, 180: 45: 5), B (hexane: diethyl ether: formic acid, 140: 72: 18) and C (toluene: acetic acid, 170: 30) were used for the TLC analysis.
Sequences and phylogenetic reconstructions
To test phylogenetic relationships with other species, nuclear internal transcribed spacer and 5.8S rDNA (ITS) and mitochondrial small subunit (mtSSU) sequences of our fresh material and other sequences retrieved from the NCBI database (GenBank) were used for molecular phylogenetic analysis. Our sampling comprised 14 species of Lobothallia including two putative new species, species of Aspicilia and Circinaria, as well as Megaspora verrucosa as an outgroup. This selection was based on the studies of Nordin et al. (Reference Nordin, Savić and Tibell2010) and a five-gene analysis by Miadlikowska et al. (Reference Miadlikowska, Kauff, Högnabba, Oliver, Molnár, Fraker, Gaya, Hafellner, Hofstetter and Gueidan2014), in which Megaspora forms a sister clade to Lobothallia and Circinaria. Information on the samples together with the GenBank Accession numbers are given in Table 1.
Table 1. Species of lichens used in the phylogenetic analyses in this study together with specimen information and GenBank Accession numbers. New specimens and associated sequences are in bold.

Methods used for DNA extraction, amplification and sequencing follow Davydov & Yakovchenko (Reference Davydov and Yakovchenko2017). ITS (531 bp) and mtSSU (725 bp) matrices were aligned separately in Geneious 6.0 (Biomatters Ltd., New Zealand) using the MUSCLE algorithm (Edgar Reference Edgar2004). Visible deviations in position homology were manually optimized. Since the specimens for the two datasets differed, we did not concatenate the matrices. Optimal substitution models were inferred separately for ITS1, 5.8S, ITS2 and mtSSU using PartitionFinder, version 1.1.1 (Lanfear et al. Reference Lanfear, Calcott, Ho and Guindon2012). The Kimura 2-parameter (K80) model was selected for the 5.8S partition, Kimura 2-parameter with gamma distribution (K80 + G) for ITS1 + ITS2, and the Hasegawa-Kishino-Yano parameter with the proportion of invariable sites and gamma-distribution (HKY + I+G) for the mtSSU. Bayesian inference with the Markov chain Monte Carlo (BMCMC) method (Larget & Shimon Reference Larget and Shimon1999) was performed using MrBayes 3.2.3 (Ronquist et al. Reference Ronquist, Teslenko, van der Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012). Three parallel Bayesian analyses were run using six chains and every 200th generation was sampled. Convergence of the chains was inferred by calculating the average standard deviation of split frequencies every 100 000 generations using a burn-in fraction of 0·5, and the runs terminated when the standard deviation of split frequencies dropped below 0·001. This was the case after 7·1 M generations for ITS and 13·9 M for the mtSSU analyses. The first 50% of trees was discarded as burn-in and a 50% majority-rule consensus tree was calculated from the remaining trees of three runs with the sumt command implemented in MrBayes 3.2.3. The most likely tree and 1000 rapid bootstrap replicates were calculated using RAxML 8.0.26 (Stamatakis Reference Stamatakis2014) by raxmlGUI software version 1.3.1 (Silvestro & Michalak Reference Silvestro and Michalak2012), applying the GTRGAMMA model of substitution to the subsets. The tree topologies are taken from Bayesian inference (Figs 1 & 2). Bootstrap support values and BMCMC posterior probability were noted on the best-scoring tree.

Fig. 1. Maximum likelihood (ML) phylogeny of selected Lobothallia ITS sequences. The reliability of each branch was tested by ML and Bayesian methods. Numbers at tree nodes indicate Bayesian inference with the Markov chain Monte Carlo (BMCMC) posterior probabilities (left) and ML bootstrap percentages (right). Thicker branches indicate when the bootstrap value of ML is ≥70% or the BMCMC posterior probability is ≥0·95. The Lobothallia clade is arrowed. GenBank Accession numbers are given to serve as operational taxonomic unit (OTU) names (see Table 1). For ‘Lecanora’ bogdoënsis, the quotation marks indicate the genus name prior to synonymy in the present paper. Originally produced sequences are marked in bold. Megaspora verrucosa was used as an outgroup.
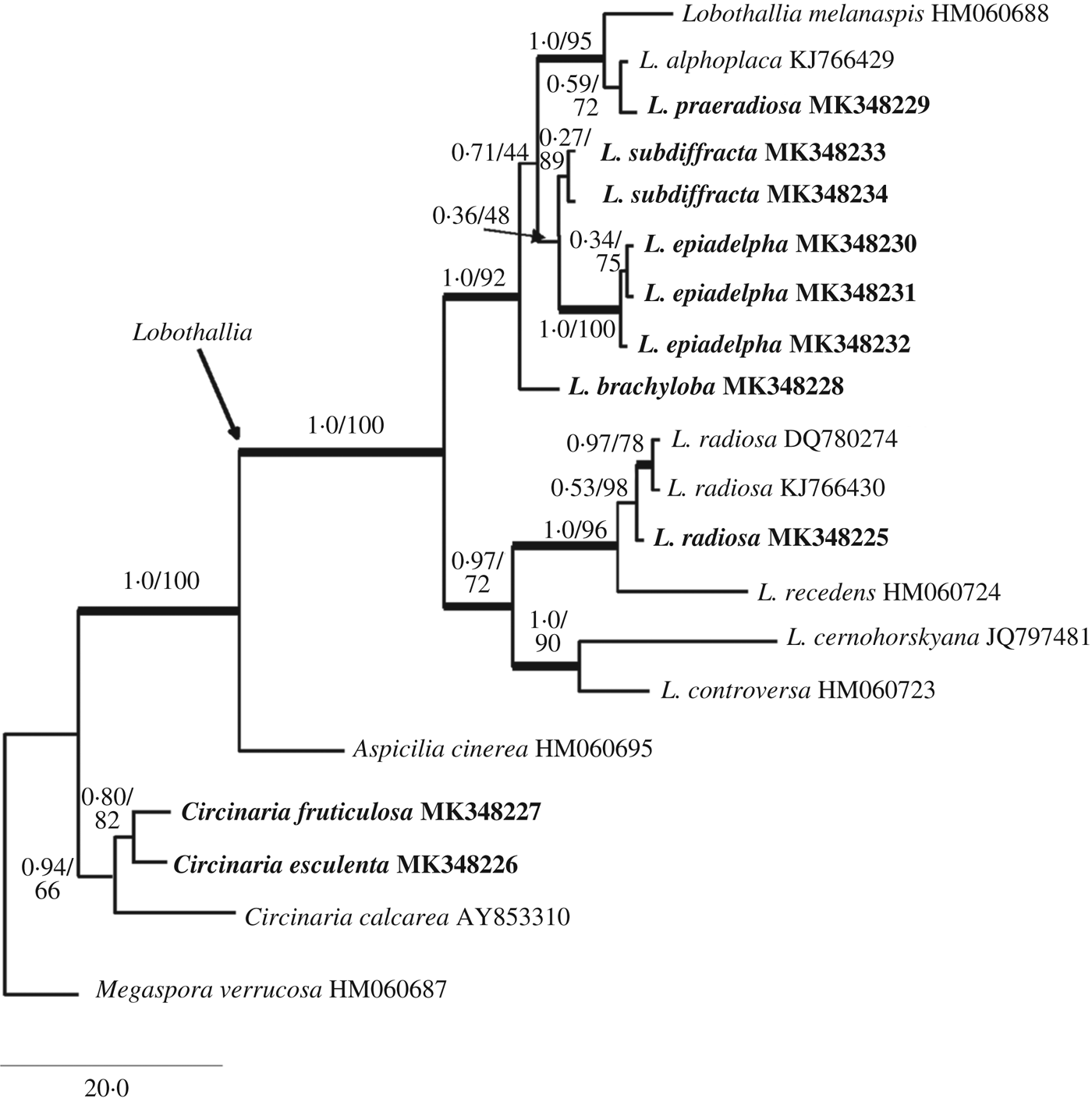
Fig. 2. Maximum likelihood (ML) phylogeny of selected Lobothallia mtSSU sequences. The reliability of each branch was tested by ML and Bayesian methods. Numbers at tree nodes indicate Bayesian inference with the Markov chain Monte Carlo (BMCMC) posterior probabilities (left) and ML bootstrap percentages (right). Thicker branches indicate when the bootstrap value of ML is ≥70% or the BMCMC posterior probability is ≥0·95. Lobothallia clade is arrowed. GenBank Accession numbers are given to serve as operational taxonomic unit (OTU) names (see Table 1). Originally produced sequences are marked in bold. Megaspora verrucosa was used as an outgroup.
Results
Both ITS and mtSSU sequences were successfully obtained from Aspicilia subdiffracta and two putative new Lobothallia species, described below as Lobothallia brachyloba and L. epiadelpha. The material of Lecanora bogdoënsis gave ITS sequences only. The Bayesian 50% majority-rule consensus tree had the same topology as the maximum likelihood tree generated by RAxML for both ITS and mtSSU and the major taxon grouping was similar in the phylogenetic reconstructions of single ITS and mtSSU (Figs 1 & 2).
Both the ITS and mtSSU phylograms combine two well-supported major clades for Lobothallia. The first major clade includes Lobothallia radiosa and L. recedens in the ITS tree with 1·00/93 support values (BMCMC posterior probability/bootstrap value, respectively); in the mtSSU phylogram this clade (0·97/72) additionally contains L. cernohorskyana and L. controversa. The second major clade (ITS: 0·99/75, mtSSU: 1·00/92) combines the remaining species from our selection and includes Lobothallia brachyloba and L. epiadelpha. In the ITS phylogram, both species cluster with low statistical support (0·89/66) as paraphyletic lineages to a well-supported clade combining Lobothallia crassimarginata, L. subdiffracta and L. helanensis. In the mtSSU, three sequences of L. epiadelpha form a monophyletic group (1·00/100) but its relation to other species within the second major clade lacks statistical support. Lecanora bogdo ë nsis is nested within the Lobothallia praeradiosa group (ITS: 1·00/100), and Lobothallia subdiffracta is placed sister to L. helanensis (ITS: 1·00/100). In the mtSSU tree (Fig. 2), Lobothallia brachyloba is sister to the second major clade, comprising Lobothallia alphoplaca, L. melanaspis, L. praeradiosa, L. subdiffracta and L. epiadelpha.
The Species
Lobothallia brachyloba Paukov & I. V. Frolov sp. nov.
MycoBank No.: MB 827340
Lobothallia with a thin, lobate, grey thallus, containing norstictic acid. Lobes flat, firmly attached to the substratum, apothecia immersed, 1–7 per areole, not crowded, lacking distinct thalline margin.
Type: Russia, Republic of Altai, Shebalino District, c. 1 km NE of village of Topuchaya, gneiss rocks on SW-slope above River Sarlyk, elev. c. 1200 m a.s.l., on gneiss, 7 July 2012, I. Frolov 357 (UFU–L3254—holotype; UPS—isotype).
(Fig. 3A & B)

Fig. 3. A & B, Lobothallia brachyloba, holotype: A, part of a thallus; B, central part of the thallus showing apothecia. C–F, Lobothallia zogtii, holotype: C & D, part of a thallus; E, central part of a thallus with apothecia; F, part of a thallus from a shady habitat. Scales = 1 mm. In colour online.
Life habit lichenized, not lichenicolous.
Thallus light grey, 0·3–1·0 mm thick, placodioid, lobate in the periphery and areolate in the central part. Lobes 0·9–2·0 × 0·6–1·0 mm (length × width), flat to slightly convex, not overlapping, closely adnate to the rocky substratum, not detaching in the outer parts. Central areoles 0·5–2·0 mm, irregular in form, flat when sterile, moderately convex when fertile. Upper cortex paraplectenchymatous, (25·0–)32·3–33·4–34·4(–42·5) μm (n = 20) over algal stacks, opaque, light brownish throughout, translucent in K, cells 4–8 μm, epinecral layer (7·5–)11·7–12·3–12·8(–17·5) μm (n = 20). M edulla (75·0–)87·5–89·4–91·2(–105·0) μm (n = 20) opaque with numerous crystals. Photobiont layer interrupted by fungal tissue, forming separate groups 50–65 μm tall and 25–45 μm wide; almost continuous, or in groups visible without staining under the hypothecium, algae 6–14 μm diam. Prothallus absent. Vegetative propagules absent.
Apothecia cryptolecanorine, 1–7 per areole in the outer part of the thalli, 1–2 in the central part, immersed, only slightly projecting when mature, (0·30–)0·39–0·41–0·43(–0·60) mm (n = 20); disc initially dot-like, later wider, slightly concave to flat, sparingly pruinose, dark brown, with a bulge of sterile tissue in some mature apothecia; thalline margin absent. Exciple of radiating hyphae, 10–12 μm thick, widening to 20–25 μm in the uppermost part, scarcely discernible. Hymenium hyaline, (65·0–)72·3–73·5–74·7(–82·5) μm high (n = 20), yellowish blue in I; paraphyses simple, submoniliform; epihymenium brownish, unchanged in N, (12·5–)13·5–14·0–14·5(–20·0) μm high (n = 20). Hypothecium hyaline, I+ bluish, (30·0–)37·8–38·8–39·7(–45·0) μm high (n = 20). Asci clavate, Aspicilia-type; ascospores broadly ellipsoid, to almost spherical, hyaline, aseptate (10·0–)11·5–11·9–12·0(–13·0) × (7·0–)9·5–9·8–10·0(–12·0) μm (n = 25).
Pycnidia common, with punctiform ostiole; conidia short, bacilliform, hyaline, aseptate (5·0–)5·7–5·8–5·9(–7·0) μm (n = 30).
Chemistry
Thallus K+ red, C−; medulla K+ red, C−; norstictic acid by TLC.
Etymology
The name reflects the shorter lobes of the species compared to Lobothallia radiosa, which can be similar in appearance.
Ecology
Lobothallia brachyloba was found on exposed acid rocks at an elevation of 1200 m a.s.l. in the boreal forest belt dominated by Picea obovata Ledeb. and Pinus sibirica Du Tour. The following species co-occurred with L. brachyloba: Aspicilia cinerea, Bellemerea cupreoatra (Nyl.) Clauzade & Cl. Roux, Lecanora cenisia Ach. and Protoparmeliopsis muralis (Schreb.) M. Choisy.
Distribution
The species is known from one locality in the Altai mountains (Republic of Altai, Russia).
Notes
Lobothallia brachyloba is a species with a placodioid, light grey thallus and norstictic acid as a secondary metabolite. This combination of characters renders it similar to other Lobothallia species, such as L. crassimarginata, L. praeradiosa and to some extent L. alphoplaca. Lobothallia brachyloba differs from these species by its flat to slightly convex, closely adnate lobes and permanently immersed apothecia without a thalline margin. Lobothallia radiosa chemotype radiosa is externally similar to L. brachyloba but differs in having a K− thallus. The norstictic acid-containing chemotype of Lobothallia radiosa (=Aspicilia subcircinata (Nyl.) Coppins) can be separated from L. brachyloba by its apothecia which are crowded in the centre of the thallus, and finally form visible thalline margins (thalline margin absent in L. brachyloba). The central areoles of Lobothallia radiosa are uneven and crossed by linear depressions which divide the areoles into smaller units, while L. brachyloba has smooth, un-cracked central areoles.
Lobothallia epiadelpha Paukov & A. Nordin sp. nov.
MycoBank No.: MB 827338
Lobothallia with a thick, lobate, brownish thallus, containing stictic acid, initially developing on species of Circinaria, later free-living. Lobes are smooth, flat or slightly convex, loosely attached to the substratum, apothecia sessile.
Type: Russia, Orenburgskaya Oblast’, Gaiskiy District, 17 km to the west of Novotroitsk Town (via Akkermanovka), hills around ‘The King's Spring’, 51°08′56·1″N, 58°02′32·0″E, elev. 299 m a.s.l., on serpentine outcrops in a dry stony steppe, 17 June 2016, A. Paukov 1881 (UFU–L3189—holotype; G, LE, M, UPS—isotypes).
(Fig. 4A–F)
Life habit lichenized, initially lichenicolous, developing mainly on Circinaria spp., later overgrowing the host and free-living. Thallus brown, up to 1·5 mm thick, lobate in the periphery and squamulose in the central part. Lobes relatively short and wide, 1·0–2·3 × 0·5–2·7 mm (length × width), flat to moderately convex, white-bordered, smooth or with irregular cracks, widened paw-like and incised in the outermost part to form 2–3 lobules or entire, overlapping, attached to the rocky substratum only by their base, but when parasitic usually firmly attached to the host, lower side whitish to light brownish. Central squamules 0·8–2·7 mm, irregular in form, constricted at the base, slightly overlapping, moderately convex. Upper cortex paraplectenchymatous, (22·5–)30·1–31·3–32·6(–41·5) μm high (n = 15) over algal stacks, brownish in its outermost part (Subdepressa-brown; see Roux et al. Reference Roux, Nordin, Tibell and Sohrabi2011), cells 7–9 μm, epinecral layer (8·5–)12·3–12·8–13·4(–16·0) μm high (n = 15). M edulla I−, K+ yellow. Lower cortex whitish. Photobiont layer interrupted by fungal tissue, forming algal stacks (Vondrák & Kubásek Reference Vondrák and Kubásek2013), 100–125 μm tall and 28–66 μm wide; spherical algal groups (21·0–)31·0–32·7–34·4(–41·5) μm (n = 10) present under hypothecium. Photobiont chlorococcoid, algae 7–20 μm diam. Prothallus absent. Vegetative propagules absent.

Fig. 4. A–F, Lobothallia epiadelpha: A, part of a thallus with wide lobes, holotype; B, young specimen overgrowing Circinaria maculata, isotype; C, Lobothallia epiadelpha on C. maculata, isotype; D, section of thallus, holotype; E, section of apothecium coloured by iodine solution, holotype; F, type locality in the Orenburg Oblast’. G & H, Lobothallia hedinii, holotype: G, outer part of a thallus; H, apothecia. Scales: A, B, G & H = 1 mm; C = 1 cm; D & E = 50 μm. In colour online.
Apothecia lecanorine, 1–4 per squamule, initially immersed, later sessile, constricted at the base, rounded or elliptic in outline, (0·5–)0·8–0·9–1·0(–1·5) mm diam. (n = 25); disc initially dot-like, later wide, flat to slightly convex, pruinose when young, black to chestnut brown in older apothecia; thalline margin appearing in dot-like apothecia as a dark zone around the disc, later well developed, thick, 0·15–0·25 mm, projecting, dark, concolorous with the disc, in older apothecia becoming lighter and receding. Exciple of radiating hyphae, 12–20 μm, widening to 45–50 μm in the uppermost part. Hymenium hyaline, (60·5–)66·8–68·2–69·6(–77·0) μm high (n = 15), unchanged in I; paraphyses moniliform with 3–7 apical cells 5·0–6·5 μm thick; epihymenium brownish, N+ weakly greenish, giving an impression of a paraplectenchymatous tissue because of swollen upper cells of the paraphyses, (13·5–)17·3–18·0–18·6(–22·5) μm high (n = 15). Hypothecium hyaline, I+ weakly bluish, (26·0–)30·0–31·1–32·0(–40·0) μm high (n = 15). Asci clavate, Aspicilia-type; ascospores broadly ellipsoid, hyaline, aseptate (10·0–)11·9–12·1–12·4(–14·5) × (6·0–)9·0–9·2–9·5(–11·8) μm (n = 30).
Pycnidia common, with punctiform ostiole; conidia short, bacilliform, hyaline, aseptate (5·0–)5·8–5·9–6·0(–7·0) μm long (n = 32).
Chemistry
Thallus K−, C−; medulla K+ yellow, C−; stictic acid complex by TLC, norstictic acid as a minor substance found in one specimen.
Etymology
The name refers to the parasitic life habit of the species, which grows on representatives of the same family Megasporaceae.
Ecology
Lobothallia epiadelpha was found in arid conditions on exposed siliceous rocks (schistose and serpentine) in steppe communities at elevations 200–1200 m a.s.l. It grows predominantly as a free-living saxicolous lichen but younger thalli commonly overgrow species of Circinaria, mainly Circinaria maculata (H. Magn.) Q. Ren (Fig. 4C). The following species co-occurred with Lobothallia epiadelpha: Acarospora irregularis H. Magn., Aspicilia cinerea (L.) Körb., Bellemerea cupreoatra, Candelariella vitellina (Ehrh.) Müll. Arg., Circinaria spp., Lecanora argopholis (Ach.) Ach., Protoparmeliopsis muralis, Rusavskia spp. and Xanthoparmelia delisei (Duby) O. Blanco et al.
Distribution
The species is known from three localities in the Orenburg Oblast’ of Russia, and Central and Eastern Kazakhstan.
Notes
Lobothallia epiadelpha may be confused with Lobothallia radiosa chemotype parasitica due to its lichenicolous habit and stictic acid as a secondary metabolite. The latter taxon differs by its thinner thallus with closely adnate not overlapping lobes, and the immersed apothecia with smaller discs (up to 0·6 mm), which are numerous in each areole. It is known only from Mediterranean and sub-Mediterranean regions and grows on other hosts (Bouly de Lesdain Reference Bouly de Lesdain1931; Loppi & Mariotti Reference Loppi and Mariotti1995) or may be free-living (Roux et al. Reference Roux, Monnat, Gonnet, Gonnet, Poumarat, Esnault, Bertrand, Gardiennet, Masson and Bauvet2017).
Additional material examined. Russia: Orenburgskaya Oblast’: Gaiskiy District, vicinity of Novotroitsk Town, 9·5 km to the SW of Khabarnoye settlement, 51°06′28·7″N, 58°06′35·6″E, 205 m, serpentine outcrops in steppe, 2013, A. Paukov AGP20130528-06 (UFU–L1705); 1 km to the east of Karagay-Pokrovka Village, steep slope on a bank of a brook confluent to Guberlya River, 51°37′54·8″N, 57°55′45·8″E, 413 m, slate outcrops in steppe, 2013, A. Paukov AGP20130529-21 (UFU–L1882).—Kazakhstan: Shyghys Kazakhstan: along the road NE of Alekseyevka, 48°29′N, 85°52′E, 1200 m, on schistose rocks, 1993, R. Moberg & A. Nordin K22:19 (UPS–L078045); along the road S of Ajaguz, 48°35′N, 80°39′E, 850 m, on open rocks, 1993, R. Moberg & A. Nordin K28:07 (UPS–L078051). Karagandinskaya Oblast’: Shetskiy District, rocky outcrops in steppe along the road Zharyk-Unrek, 48°51′N, 72°55′E, 2018, I. Frolov K1:2145 (UFU).
Lobothallia zogtii Paukov & Davydov sp. nov.
MycoBank No.: MB 827339
Lobothallia with a thick, lobate, brownish, non-parasitic thallus, containing stictic acid. Lobes with deep cracks, slightly to strongly convex, firmly attached to the substratum; apothecia sessile, pruinose; disc brownish black to jet black, convex, surrounded by a receding thalline margin.
Type: China, Dzhungarian Gobi, Baitag-Bogd-Ula, elev. 1800 m a.s.l., on rocks, 19 July 1981, U. Zogt 5259 (LE—holotype, isotype). Determined as Aspicilia melanaspis (Ach.) Poelt & Leuckert.
(Fig. 3C–F)
Life habit lichenized, not lichenicolous.
Thallus brown, 1·5–2·5 mm thick, lobate in the periphery and squamulose-areolate in the central part. Lobes 1·5–3·5 × 1·0–2·5 mm (length × width), moderately to strongly convex, white-bordered, with irregular deep depressions forming a reticulate pattern (smooth in shade), not overlapping, transversely cracking towards the centre of the thallus to form areoles, closely adnate to the rocky substratum, with outer part (c. 0·5–1·0 mm) unattached, lower side whitish to light brownish. Lower cortex indefinite. Central areoles 1·0–1·7 mm, irregular in form, moderately convex, with irregular depressions forming a reticulate pattern, sometimes even forming cracks subdividing the areoles (in shade the areoles are convex and smooth). Upper cortex paraplectenchymatous, (62·5–)76·9–79·3–81·8(–100·0) μm (n = 15) over algal groups, outer 10 μm of cortex brownish, with ‘Subdepressa-brown’ cells 5–8 μm, epinecral layer (17·5–)21·3–23·7–26·0(–42·5) μm (n = 20). M edulla I−, K+ yellow. Photobiont layer interrupted by a fungal tissue, forming separate columns 125–300 μm tall and 75–125 μm wide; algal layer under hypothecium is visible without staining as spherical algal groups (25·0–)35·5–37·2–38·8(–50·0) μm (n = 21) usually but not consistently present under the hypothecium. Photobiont chlorococcoid, algae 8–17 μm diam. Prothallus absent. Vegetative propagules absent.
Apothecia lecanorine, 1–2 per squamule, but sometimes forming 3–4 unclearly separated apothecia due to cracking of the primary discs, initially immersed, soon sessile, constricted at the base, rounded or elliptic in outline, (0·5–)1·2–1·3–1·4(–2·5) mm (n = 20); disc initially dot-like, later wide, slightly convex, pruinose, brownish black to jet black, chestnut brown when wet; thalline margin 0·10–0·20 mm thick, projecting when young, concolorous with the thallus, in older apothecia becoming dark and receding, not higher than the disc, sometimes with white pruina. Exciple of radiating hyphae, 17–25 μm, widening to 45–50 μm in the uppermost part. Hymenium hyaline, (70·0–)81·1–82·7–84·2(–95·0) μm high (n = 20), unchanged in I; paraphyses moniliform, with 3–7 apical cells 7–9 μm thick; epihymenium brownish, unchanged in N, (12·5–)16·6–17·3–18·1(–22·5) μm high (n = 20). Hypothecium hyaline, I+ weakly bluish, (25·0–)34·2–35·3–36·5(–45·0) μm high (n = 20). Asci clavate, Aspicilia-type; ascospores broadly ellipsoid, hyaline, aseptate (10·0–)11·2–11·4–11·7(–14·0) × (7·0–)7·9–8·0–8·1(–9·0) μm (n = 25).
Pycnidia common, with punctiform ostiole; conidia short, bacilliform, hyaline, aseptate (5·0–)5·8–6·0–6·1(–7·0) μm (n = 35).
Chemistry
Thallus K−, C−; medulla K+ yellow, C−; stictic acid complex by TLC.
Etymology
The name refers to the Mongolian botanist U. Zogt, who collected the type specimen.
Ecology
Lobothallia zogtii was found in arid steppe communities on exposed siliceous rocks at elevations 1200–1800 m a.s.l. Accompanying species are Anaptychia desertorum (Rupr.) Poelt, Candelariella rosulans (Müll. Arg.) Zahlbr., Lecidea tessellata Flörke and Protoparmeliopsis peltata (Ramond) Arup et al.
Distribution
The species is known from two localities in the Xinjiang Autonomous Region of China.
Notes
Lobothallia zogtii might be confused with L. epiadelpha due to its general appearance, the colour of the thallus and the presence of stictic acid as its main secondary metabolite. However, it has not been found overgrowing other lichens and it has narrower and convex, adnate, non-overlapping lobes with a definite reticulate pattern, formed by depressions of the upper cortex. This pattern, however, is poorly visible or even absent in thalli from shady microhabitats although it can be found in lobes of the same specimens on an exposed side of the stone substratum. Compared to L. epiadelpha, apothecia of Lobothallia zogtii are larger, usually 1–2 per areole, and have blackish (brown in shade) convex discs with receding margins, projecting in younger apothecia only.
Additional material examined. China: Xinjiang: foothills of Mongolian Altai range at the 40th km of the Qinghe-Altai road (30 km SW of Qinghe), desert steppe slopes, on rocks, Achnatherum splendens community in meso-depressions, 46°30′46·5″N, 90°03′31·5″E, 1280 m, 2007, E. A. Davydov 16027 (ALTB).
New combinations
In his works on the lichens of China, Magnusson (Reference Magnusson1940, Reference Magnusson1944) described among others two species, Lecanora hedinii H. Magn. and L. subdiffracta H. Magn., which were subsequently moved to Aspicilia (Oxner Reference Oxner1972; Wei Reference Wei1991). A year before Magnusson, Oxner himself described Aspicilia lacteola Oxner from Kyrgyzstan (Oxner Reference Oxner1939). Cretzoiu (Reference Cretzoiu1941) was one of the lichenologists who treated Aspicilia as a section in the genus Lecanora. For this reason, he proposed the name L. oxneri Cretz. when transferring A. lacteola into Lecanora to avoid a homonymy with L. lacteola Müll. Arg. Our study of the types of these species in S and KW has convinced us that their correct placement is in Lobothallia.
Lobothallia hedinii (H. Magn.) Paukov, A. Nordin & Sohrabi comb. nov.
MycoBank No.: MB 827675
Lecanora hedinii H. Magn. Lichens from Central Asia 1: 98 (1940).—Aspicilia hedinii (H. Magn.) Oxner, Nov. Sist. Nizsh. Rast. 9: 288 (1972); type: China, Kansu Prov., Eh-ma-ta-ch'üan, 1931, Bohlin 55a (S—holotype!).
Lecanora hedinii f. pruinosa H. Magn., Lichens from Central Asia 1: 100 (1940).—Aspicilia hedinii f. pruinosa (H. Magn.) J. C. Wei, Enumeration of Lichens in China (Beijing) 1991: 32 (1991); type: China, Kansu Prov., Eh-ma-ta-ch'üan, 1931, Bohlin 55b (S—holotype!), syn. nov.
(Fig. 4G & H)
The species can be recognized by the following set of characters: brown, white-pruinose thallus with norstictic acid; straight, parallel, simple or dichotomous convex lobes; immersed apothecia 1–2(3) per areole. Lobothallia pruinosa most closely corresponds to this set of characters but has flat lobes, not arranged in parallel. Other Lobothallia species with convex lobes differ from Lobothallia hedinii in their appressed apothecia.
Lobothallia lacteola (Oxner) Şenkardeşler, Paukov, Davydov & Sohrabi comb. nov.
MycoBank No.: MB 827678
Aspicilia lacteola Oxner, Zhurnal Institutu Botaniki AN URSR 20(28): 118 (1939); type: Kyrgyzstan (Kirgizskaya SSR), Dzhelalabadskiy kanton, spurs of Fergana ridge, vicinity of Arslan-Boba, 1928, Lazarenko (KW 23609!—lectotype, designated here, MycoBank typification MBT 386683). HPTLC: norstictic acid. (Other syntype traced: Kyrgyzstan (Kirgizskaia SSR), Dzhelalabadskyi kanton, spurs of Fergana ridge, limestones in the vicinity of Gava Village, 1928, Lazarenko (KW 23608!). HPTLC: norstictic and connorstictic acids).
Lecanora oxneri Cretz., Bull. Grǎd. Bot. Univ. Cluj 21: 139 (1941), syn. nov. (KW 23609!—type).
(Fig. 5D)
Lobothallia lacteola differs from other species of the genus by the following set of characters: calcareous, circular, white, farinose, distinctly rimose-areolate thallus with indefinitely radially cracked or plicate margins; immersed apothecia with black, pruinose disc and prominent thalline margin; norstictic acid as a major secondary metabolite, which is occasionally accompanied by connorstictic acid as a minor component. Additional characters are simple and non-moniliform paraphyses and 6–8-spored asci with shortly ellipsoid to globose ascospores 10·0–11·5 × 7·5–11·5 μm in size.

Fig. 5. A, holotype of Lobothallia subdiffracta, part of a thallus (S); B, L. recedens, Sweden, Dalarna, A. Nordin 6587 (UPS); C, L. controversa, Russia, Altaiskiy Krai, E. A. Davydov 17606 & L. S. Yakovchenko (ALTB); D, lectotype of L. lacteola (KW); E, L. radiosa chemotype parasitica, isotype of Lecanora parasitica (LE); F, L. radiosa chemotype radiosa, Russia, Orenburg Oblast’, A. Paukov AGP20120606-11 (UFU); G & H, L. radiosa chemotype subcircinata, France, Pyrénées-Orientales (photograph: S. Poumarat). Scales = 1 mm. In colour online.
The species is similar to Lobothallia controversa Cl. Roux & A. Nordin but differs chemically in the absence of terpenes. The thalline margin of L. lacteola projects clearly in mature apothecia while it is absent in L. controversa.
Lobothallia lacteola is known from two localities in Kyrgyzstan and one locality in Russia (Altaiskiy Krai), where it inhabits exposed limestone rocks. The species is regarded here as new to Russia.
Additional material examined. Russia: Altaiskiy Krai: Tigirekskiy Range, right bank of Inya River, 2 km upstream from Tigirek settlement, Inskaya mountain, SE slope, limestone, 51°09′N, 83°04′E, 1996, E. A. Davydov 5008 (ALTB).
Lobothallia subdiffracta (H. Magn.) Paukov comb. nov.
MycoBank No.: MB 827676
Lecanora subdiffracta H. Magn. Lichens from Central Asia 2: 40 (1940).—Aspicilia subdiffracta (H. Magn.) J. C. Wei, Enumeration of Lichens in China (Beijing) 1991: 34 (1991); type: China, Inner Mongolia, Beli-Miao, 41°30′N, 110°10′E, 1929, Bohlin (S—holotype!).
Lobothallia helanensis X. R. Kou & Q. Ren, Mycotaxon 123: 245 (2013); type: China, Inner Mongolia, Mt. Helan, elev. 1500 m, 2011, D. B. Tong 20122517 (SDNU—holotype, not seen), syn. nov.
(Fig. 5A)
The holotype of Lecanora subdiffracta kept in S is a small specimen. Nonetheless it bears all the characters typical of this species, namely a thick, non-lobate squamulose thallus, dark, white-bordered squamules and apothecia with incised margins, which agree with Lobothallia helanensis. On the basis of the similarity of the species in the ITS and mtSSU regions, as well as their morphological and anatomical similarity, we conclude that these two taxa represent the same species, with subdiffracta as an earlier epithet. In addition to the type localities of Lobothallia subdiffracta and L. helanensis in China, the species is widely distributed in arid conditions in Russia (Republic of Altai) and Mongolia.
Material examined. Russia: Republic of Altai: Kosh-Agachskiy District, SE part of Kuray Ridge, NE of village of Chagan-Uzun, 2000 m, 2012, I. Frolov 105 (UFU); 10 km S of Tarkhata, valley of Tarkhata brook, schistose rocks at the brook, 2200 m, 2012, I. Frolov 178 (UFU); Chuiskaya Steppe, right bank of the Tydtuyaryk River near its junction with Chuya River, rock outcrops, dry steppe, 50°04′18″N, 88°24′50″E, 1760 m, on rocks, 2016, A. Paukov 1952 (UFU).—Mongolia: Dschungarian Gobi: Argalant mountains, near Ubchu-Bulak, 1700 m, on rocks, 1973, N. S. Golubkova & U. Zogt (LE–L406). Gobi-Altai Aimak: Gobi-Alyk-Nuur steppe, Adgi-Bogdo ridge, 1500 m, on granite, 1973, N. S. Golubkova & U. Zogt (LE–L432); Ih-Bogdo-Ula ridge, Dzhargatantyn-Ama canyon, 2600 m, on rocks, 1973, N. S. Golubkova & U. Zogt (LE–L555). South- Gobi Aimak: Servey ridge, on rocks, 1973, N. S. Golubkova & U. Zogt (LE–L763).
New synonym
Lobothallia praeradiosa (Nyl.) Hafellner
MycoBank No.: MB 354542
Acta Bot. Malacit. 16: 138 (1991).—Lecanora praeradiosa Nyl., Flora (Regensburg) 67: 389 (1884); type: Hungary, prope Budapest, 1882, Lojka (G—isotype!).
Lecanora bogdoënsis Tomin, Pririoda i sel'skoye khozyaystvo zasushlivykh oblastey SSSR 1–2: 48[4] (1927), syn. nov.—Placolecanora bogdoënsis (Tomin) Kopach., Nov. Sist. Nizsh. Rast. 9: 295 (1972), syn. nov.—Protoparmeliopsis bogdoënsis (Tomin) S. Y. Kondr., Ukr. Bot. J. 69: 876 (2012), syn. nov. (W-L282—isotype!).
(Fig. 6F & G)
Lecanora bogdoënsis Tomin was segregated from lobate Lecanora alphoplaca on the grounds of its wider and more flattened lobes and wider conidia (Tomin Reference Tomin1927). The species was later moved to Placolecanora (Kopaczevskaja Reference Kopaczevskaja1972) and Protoparmeliopsis (Kondratyuk et al. Reference Kondratyuk, Zarei-Darki and Khajeddin2012). However, the absence of usnic acid, isousnic acid or xanthones along with small spores and short conidia imply an affiliation with Lobothallia. ITS sequences as well as the morphology and conidial width of the material collected by the authors in the locus classicus agree with those of Lobothallia praeradiosa, with which we synonymize Lecanora bogdoënsis here.

Fig. 6. A, Lobothallia chadefaudiana, Vězda, Lich. Sel. Exs. 1638 (W); B, L. cernohorskyana, Vězda, Lich. Sel. Exs. 895 (W); C, L. cheresina chemotype cheresina, Greece, Rhodos, I. Pišút s. n. (SAV); D. L. cheresina chemotype justii, type of Lecanora justii (W); E, L. alphoplaca, Russia, Bashkortostan, A. Paukov AGP20100525-37 (UFU); F, L. praeradiosa, isotype of Lecanora praeradiosa (G); G, isotype of Lecanora bogdoënsis (W); H, L. melanaspis, Russia, Altaiskiy Krai, E. A. Davydov 5907 (ALTB). Scales = 1 mm. In colour online.
Notes on Lobothallia radiosa (incl. Aspicilia parasitica B. de Lesd.), stictic acid chemotype
Aspicilia parasitica B. de Lesd.
Bull. Soc. Bot. France 78: 728 (1931); type: Italia, Genova, val Bisagno, Bosco del Serina, sur roche siliceuse, 1930, Sbarbaro (LE—isotype!).
(Fig. 5E)
Aspicilia parasitica has been the subject of two different interpretations: Esnault (Reference Esnault1985) and Loppi & Mariotti (Reference Loppi and Mariotti1995) consider it as a species distinct from Lobothallia radiosa due to its chemistry (stictic acid) and its parasitism on various species of Aspicilia (Loppi & Mariotti Reference Loppi and Mariotti1995). The parasitic nature of the species was considered to be an essential attribute because of the existence in Algeria of a non-parasitic chemotype of L. radiosa with stictic acid (Esnault Reference Esnault1985).
Roux et al. (Reference Roux, Monnat, Gonnet, Gonnet, Poumarat, Esnault, Bertrand, Gardiennet, Masson and Bauvet2017) considered Aspicilia parasitica to be a chemotype of L. radiosa with a K+ yellow thallus containing stictic acid. Even though the parasitism of this chemotype is quite common, it is not at all constant (especially in the Southern Alps or in young thalli which often begin their development unattached to other lichens). Otherwise, the chemotype radiosa, which is devoid of secondary chemistry, parasitizes other Aspicilia s. lat. (especially in le Var and in Corsica).
Lobothallia radiosa with stictic acid (chemotype parasitica), which is also known from the Pyrénées-Orientales (Roux et al. Reference Roux, Monnat, Gonnet, Gonnet, Poumarat, Esnault, Bertrand, Gardiennet, Masson and Bauvet2017), has a more southern distribution (Mediterranean and sub-Mediterranean) than chemotype radiosa, but their ranges widely overlap. Esnault (Reference Esnault1985: 116) provided two other distinguishing characters of the chemotype parasitica, namely a darker colour of the thallus and the presence of an amorphous cortical coating (epinecral layer). The examination of several specimens of Lobothallia radiosa (chemotype radiosa (Fig. 5F), without or with a trace amount of norstictic acid; chemotype subcircinata with norstictic acid (Fig. 5G & H); chemotype parasitica, including isotype material, which parasitizes Aspicilia proluta, collected by Sbarbaro) has shown unambiguously that these two characters are not informative. The colour of the thallus is variable and the presence of an epinecral layer occurs in all the specimens examined but its thickness varies, depending on the part of the thallus examined, from 0 to 30 μm. Hence, we follow here the view of Roux et al. (Reference Roux, Monnat, Gonnet, Gonnet, Poumarat, Esnault, Bertrand, Gardiennet, Masson and Bauvet2017) and include Aspicilia parasitica in Lobothallia radiosa.
Selected specimens examined.
Сhemotype radiosa. France: Pyrénées-Orientales: Jujols, beginning of the Garrigue trail, 1025 m, on inclined or subvertical surfaces (10–80°), non-calcareous schist, 2007, C. Roux 25139 (MARSSJ); Nohèdes, in the immediate vicinity of the Nohèdes Nature Reserve, 170 m to the NW of Cortal, 1030 m, on small non-calcareous shale walls, 2009, C. Roux 25614 (MARSSJ).—Italy: Liguria Occidentale: Spotorne (Savona), in regione ‘Castelo’, ad rupes schistosas, 1962, C. Sbarbaro (Vězda, Lichenes Selecti Exsiccati no 167; sub ‘Lecanora parasitica (B. de Lesd.) Zahlbr.’), but the specimen in MARSSJ is not parasitic and K−.
Сhemotype parasitica (parasitic). France: Alpes-Maritimes: Saorge, bottom of the Mérim Valley, 930 m, on a steeply sloping slab of non-calcareous Permian red sandstone, on Aspicilia viridescens (A. Massal.) Hue, 2011, C. Roux 26926 (MARSSJ).—Italy: Liguria occidentale: Val Bisagno, S. Eusebio sub castaneis, on Aspicilia proluta, C. Sbarbaro (hb. Bouly de Lesdain in MARSSJ, topotypus or isotypus).
Сhemotype parasitica (not parasitic). France: Pyrénées-Orientales: Jujols, beginning of the Garrigue trail, 1025 m, on inclined or subvertical surfaces (10–80°), non-calcareous schist, 2007, C. Roux 25154 (MARSSJ).
Сhemotype subcircinata. France: Alpes-de-Haute-Provence: Uvernet-Four, bottom of the Bachelard Gorges to the N of Uvernet-Four and the Courriers bridge, along the D902 road, 1250 m, on calcareous rock block (flysch du Pelat), shaded by Pinus sylvestris, 2010, C. Roux 26930 (MARSSJ); Thoard, near the Siron Peak, on exposed limestone outcrops, 1650 m, 1965, G. Clauzade (hb. G. Clauzade in MARSSJ). Alpes-Maritimes: Saint-Sauveur-sur-Tinée, a little to the S of the village, on the edge of the road (D2205), 483 m, on large rocks in a red pelites location, 2013, C. Roux 6337 (MARSSJ).
Additional species reported
Lobothallia controversa Cl. Roux & A. Nordin
Herzogia 29: 588 (2016); type: France, Bas-Vivarais, Ardèche, Rochecolombe, 2008, Bricaud, Bauvet & Roux (UPS-L205562—holotype).
(Fig. 5C)
The systematic position of this species was clarified only recently (Roux et al. Reference Roux, Nordin and Bertrand2016). It is the only known representative of Megasporaceae containing terpenes. Lobothallia controversa belongs to the group of species with a thick, whitish, rimose-areolate thallus. It differs from morphologically similar taxa by its apothecia which are pruinose, irregular in outline and without margins. Lobothallia controversa is known from Algeria and France (Roux et al. Reference Roux, Nordin and Bertrand2016) and has been recently collected in the Altai region. The species is reported here as new to Russia and Asia.
Material examined. Russia: Altaiskiy Krai: Krasnoshchëkovskiy District, Tigirekskiy Range, Tigirekskiy Nature Reserve, Khankharinskiy cluster, left bank of the ‘Dragunskiy klyuch’ stream, 5·8 km NW of the community of Tigirek, 51°11′09″N, 82°58′30″E, 870 m, 2014, E. A. Davydov 17606 & L. S. Yakovchenko (ALTB).
Discussion
Lobothallia is the third largest genus within Megasporaceae, currently comprising 18 species. It is widely distributed in arid and mountainous regions of the Northern Hemisphere but the number of species reported in different parts of Eurasia varies. The highest diversity of Lobothallia (12 species) is currently known from the Altai Mountains (Sedelnikova Reference Sedelnikova1990; Davydov Reference Davydov2014; Paukov et al. Reference Paukov, Shiryaeva and Davydov2018) and adjacent territories of China (Magnusson Reference Magnusson1940, Reference Magnusson1944; Wei Reference Wei1991; Kou et al. Reference Kou, Li and Ren2013). Six of these, viz. Lobothallia brachyloba, L. crassimarginata, L. hedinii, L. pruinosa, L. subdiffracta and L. zogtii, are known only in a small number of localities in this region. The Alps, together with the Mediterranean and sub-Mediterranean regions, have 10 species of Lobothallia (Roux Reference Roux2012; Roux et al. Reference Roux, Nordin and Bertrand2016; Nimis et al. Reference Nimis, Hafellner, Roux, Clerc, Mayrhofer, Martellos and Bilovitz2018), while territories situated north of latitude 50°N maintain a much lower diversity of Lobothallia. Only five species are known both from Fennoscandia and the Ural Mountains, with none endemic to these regions (Paukov & Trapeznikova Reference Paukov and Trapeznikova2005; Paukov Reference Paukov2009; Nordin et al. Reference Nordin, Moberg, Tønsberg, Vitikainen, Dalsätt, Myrdal, Snitting and Ekman2011a).
The highest known diversity of Lobothallia in Eurasia occurs in areas which belong to the Mediterranean and to a lesser extent the temperate and boreal bioclimates, according to Rivas-Martínez et al. (Reference Rivas-Martínez, Rivaz-Sáenz, Penas and Díaz2004). Equivalent regions which could be similarly high in diversity of this genus are south-western Asia and arid parts of southern Asia, which lie between the Mediterranean region and Altai and belong to the same type of bioclimate. However, seven and six species are currently known from Turkey and Iran, respectively (John Reference John1996; Seaward et al. Reference Seaward, Sipman and Sohrabi2008; Kinalioğlu Reference Kinalioğlu2010; Sohrabi et al. Reference Sohrabi, Stenroos, Myllys, Søchting, Ahti and Hyvönen2013b). This may be connected with the Cenozoic history of the continent during the Oligocene and middle Miocene epochs, when western Asia was continuously being modified by the rise and subsequent retreat of the Tethys Sea (Popov et al. Reference Popov, Rögl, Rozanov, Steininger, Shcherba and Kovac2004). These geological events, which also appear to correlate with diversification times in some other lichen genera (Leavitt et al. Reference Leavitt, Esslinger, Divakar and Lumbsch2012; Cornejo & Scheidegger Reference Cornejo and Scheidegger2018), might have played a role in the lower diversity of Lobothallia in western Asia.
Key to species of Lobothallia
The characters in the following key are based on our observations and measurements, and on additional information from Clauzade & Roux (Reference Clauzade and Roux1985), Ryan (Reference Ryan, Nash, Ryan, Diederich, Gries and Bungartz2004) and Kou et al. (Reference Kou, Li and Ren2013).
1 Marginal lobes absent or indistinct; thalli may have incised, plicate margins or tiny,
infrequent and irregular lobules. In radial specimens the ‘lobes’ are formed by cracking of the peripheral zone by splits moving from the central parts of thalli ………2
Marginal lobes constant and well developed. In closely adnate specimens the splits originate mostly from the outside of the thalli………10
2(1) On calcareous rocks. Thallus off-white, partly with yellowish or greyish tinge………3
Оn siliceous rocks. Thallus light grey, dark grey, olive grey to brownish………9
3(2) Thallus ±continuous with granules on the surface or consisting of areoles with multiple cracks and having granulate appearance. Norstictic acid or no lichen substances………4
Thallus rimose-areolate to distinctly areolate at least in the central part. Areoles not granulate, with smooth or farinose upper surface. Terpenes, norstictic, stictic acid or no lichen substances………5
4(3) Lacks lichen substances, K−. Thallus thick, up to 1·5 mm, continuous to rimose, with rough yellowish granules on the surface. Apothecia immersed, separated from the thallus by thin cracks………Lobothallia chadefaudiana (Fig. 6A)
Contains norstictic acid, K+ red. Thallus up to 0·5 mm thick, rimose to areolate; areoles have granulate appearance. Apothecia appressed, becoming constricted at the bases with time………Lobothallia cernohorskyana (Fig. 6B)
5(3) Thallus contains terpenes and ±norstictic or stictic acid, 1–4 mm thick. Apothecia without visible thalline margin………Lobothallia controversa (Fig. 5C)
Thallus contains no terpenes………6
6(5) Thallus off-white, thick, 0·5–2·0 mm, superficial, with ± farinose, continuous or finely cracked, plicate margins, lacking long and straight radial cracks. Thalline margin finally prominent. Contains norstictic and ±connorstictic acids………Lobothallia lacteola (Fig. 5D)
Thallus whitish, commonly with grey tinge, thin to thick, sometimes semi-immersed, not farinose, with definite cracks up to the margins. Orbicular specimens usually with long, straight, open, radially arranged cracks. Lacking prominent thalline margin. Secondary metabolites absent or thallus contains stictic/norstictic acids………7
7(6) Contains no detectable secondary metabolites……… Lobothallia cheresina chemotype cheresina (Fig. 6C)
Norstictic or stictic acid present………8
8(7) Stictic acid as a main secondary metabolite……… Lobothallia cheresina chemotype justii (Fig. 6D)
Norstictic acid as a main secondary metabolite……… Lobothallia cheresina chemotype microspora
9(2) Thallus areolate, light to dark grey to brownish. Thalline margin of apothecia smooth, projecting, later receding. Disc brownish. Europe, Caucasus……… Lobothallia recedens (Fig. 5B)
Thallus squamulose, light to dark grey to olive grey, pruinose. Squamules with deep cracks, outer squamules usually slightly enlarged. Thalline margin incised. Disc blackish. China (Gansu, Inner Mongolia); Russia (Altai), Mongolia……… Lobothallia subdiffracta (Fig. 5A)
10(1) Thallus with stictic acid as a main lichen substance………11
Thallus with norstictic acid as a main lichen substance or secondary metabolites absent………13
11(10) Thallus light grey to dark or brownish grey, growing on non-calciphilous Aspicilia species, closely attached to the host or occasionally not parasitic. Apothecia immersed or slightly projecting, up to 0·6 mm, abundant and crowded in the central parts of thalli. Mediterranean region……… Lobothallia radiosa chemotype parasitica (Fig. 5E)
Thallus olive brownish to shades of brown, without hints of grey, free-living or parasitic in the early stages of development. Mature apothecia larger than 0·8 mm, sessile, constricted at the base, not crowded. South Urals (Russia), Kazakhstan, Xinjiang Autonomous Region of China………12
12(11) Young thalli parasitic on Circinaria maculata, later free-living. Lobes of non-parasitic thalli loosely attached, overlapping, smooth or with unclear cracks. Apothecia with dark brown flat discs. Thalline margin permanent, thick……… Lobothallia epiadelpha (Fig. 4A–E)
Thalli free-living, lobes firmly attached to the substratum, not overlapping, with definite deep cracks forming a reticulate pattern in exposed habitats. Apothecia with brownish black to jet black, finally convex discs. Thalline margin receding in mature apothecia………Lobothallia zogtii (Fig. 3C–F)
13(10) Thalli entirely or partly loosely attached to the substratum or at least outer 1–4 mm of lobes not adherent. Lobes ±overlapping. Mature apothecia sessile with constricted base………14
Thalli closely adnate to the substratum almost up to the margins, lobes not overlapping. Mature apothecia immersed to projecting (but check Lobothallia radiosa in couplets 19 and 21)………16
14(13) Thallus lacks secondary metabolites. Lobes dark grey, greenish when wet, repeatedly branching. In wet habitats………Lobothallia melanaspis (Fig. 6H)
Thallus with norstictic acid. In dry habitats………15
15(14) Thalli loosely attached to the substratum, separate lobes or even whole thalli can be detached almost intact (less evident in younger specimens). Lobes strongly convex to almost cylindrical, whitish grey, rarely with light shades of brown, side margins of lobes never arranged in parallel. Central ‘areoles’ bullate with strongly swollen tips and constricted bases……… Lobothallia alphoplaca (Fig. 6E)
Thalli normally closely adnate to the substratum with only outer 1–4 mm of lobes not adherent (but specimens overgrowing lichens/mosses or older parts of thalli may strongly resemble the previous species as these areas are easily detachable). Lobes flat to moderately convex, grey to distinctly brownish, often with side margins arranged in parallel in the closely adnate parts. Central ‘areoles’ flat to moderately convex or uneven, not bullate………Lobothallia praeradiosa (Fig. 6F)
16(13) On inundated rocks in summer-dry creeks. Thallus lead or bluish grey, without pruina. Areoles angular, apothecia dark, 1–6 on areole, 0·5–1 mm diam., immersed, immarginate or rarely with indistinct dark margins. Sardinia……… Lobothallia hydrocharis
In dry, not inundated habitats. Thalli variously coloured, grey, whitish, brownish, often pruinose. Apothecia with margins, if immarginate, less than 0·6 mm diam. Distribution various………17
17(16) Apothecia immersed to slightly projecting, normally less than 0·6 mm………18
Mature apothecia projecting to sessile (if slightly projecting, then more than 0·6 mm)………20
18(17) Thallus brown, white-pruinose. Lobes strongly convex, simple to dichotomous, with ±straight and parallel margins. Apothecia 1–2 on areole. Contains norstictic acid. China (Gansu)………Lobothallia hedinii (Fig. 4G & H)
Thallus light to dark grey, whitish, rarely brownish, pruinose or not. Lobes flat, apothecia 1–7 on areole. Contains norstictic acid or no secondary metabolites………………19
19(18) Thallus light grey, epruinose, contains norstictic acid. Apothecia immersed, circular, without margins, always smaller than the areole. Central areoles with ±smooth surface………Lobothallia brachyloba (Fig. 3A & B)
Thallus from chalky white to dark grey with brownish tint, pruinose when growing on limestone, without (chemotype radiosa) or with (chemotype subcircinata) norstictic acid. Apothecia immersed to slightly projecting, finally with visible margins, crowded on central areoles and angular. Central areoles with uneven surface and crossed by depressions or cracks which finally divide them into smaller units and, in turn, have single apothecia………Lobothallia radiosa (Fig. 5F–H)
20(17) Lobes strongly convex, short, 1–2 mm, dichotomous or simple, grey, nonpruinose. Apothecia with blackish discs and thick, permanent thalline margin (thickness equal to or more than the radius of the disc). Norstictic acid……… ………Lobothallia crassimarginata
Lobes flat or slightly convex, whitish, greyish to brownish or brownish grey, often pruinose. Apothecia single or crowded, with narrower margins………21
21(20) Lobes flat, grey to brownish, short, 1–2 mm long, 0·7–1 mm wide. Pruina on the margins of areoles or covering whole surface and discs of apothecia. Apothecia solitary, slightly projecting, discs 0·7–1·2 mm. Norstictic acid. China……… ………Lobothallia pruinosa
Lobes flat to moderately convex, chalky white to brownish grey, 3–5 mm long, 0·5– 1·5 mm wide, with or without pruina. Apothecia initially crowded and angular, extended period immersed, later projecting to broadly sessile 0·5–1·5 mm. No lichen substances or norstictic acid. Widespread……… ………Lobothallia radiosa, see also couplet 19 (Fig. 5F–H)
ED thanks Dr Wen-Li Chen, Dr Bing Liu and Dr D. German for organizing the expedition to China. The study by AP, AT and AS was partly financially supported by the Russian Foundation for Basic Research (grants 16-04-01346 and 18-04-00414). The work of AT was additionally supported by the Ministry of Science and Higher Education of the Russian Federation as part of the state assignment number 6.7696.2017/8.9. AP is grateful to the Ministry of Science and Higher Education of the Russian Federation (agreement no. 02.A03.21.0006) for financial support during his work in herbaria, and to Philippe Clerc (G), Anna Guttová, Ivan Pišút (SAV), Anton Igersheim (W), Jens Klackenberg (S), Sergiy Kondratyuk (KW), Leena Myllys, Teuvo Ahti (H) and Harrie Sipman (B) for their hospitality. AP, AT and AS thank Stanislav Glagolev and Nikolai Pirogov (Bogdinsko-Baskunchakskiy Reserve) for permission to visit the reserve and for their help during the fieldwork. Alexander Ivanov (Ural Federal University) kindly provided us with specimens of Lobothallia from Kazakhstan. Ivan Frolov worked in the frame of the national project of the Botanical Garden Institute (Russian Academy of Sciences, Ural Branch). The authors thank Serge Poumarat (Marseille) for providing us with the photographs of Lobothallia radiosa chemotype subcircinata. And finally, we would like to express our gratitude to anonymous reviewers and associate editor Toby Spribille whose valuable comments and meticulous reading allowed us to make substantial improvements to the manuscript.







