


Introduction
Sphinctrina Fr. was described by E. Fries (Reference Fries1825, Reference Fries1828) and placed among the fungi. Later, Th. M. Fries (Reference Fries1861) relocated the genus, including it with other genera of Caliciales s. lat. such as Calicium Ach. or Chaenotheca (Th. Fr.) Th. Fr. Currently, Sphinctrina is placed in Sphinctrinaceae, together with Pyrgidium, according to phylogenetic analyses using molecular markers (Wedin & Tibell Reference Wedin and Tibell1997; Tibell & Wedin Reference Tibell and Wedin2000; Tibell & Vinuesa Reference Tibell and Vinuesa2005).
Sphinctrina was characterized by the shape of the capitulum, size of asci and spores, and the lack of a well-developed thallus. Löfgren & Tibell (Reference Löfgren and Tibell1979) recognized five species, four of them with non-septate spores [S. anglica Nyl., S. leucopoda Nyl., S. tubiformis A. Massal. and S. turbinata (Pers. ex Fr.) de Not.] and one with 1-septate spores (S. prorrectula Nyl.). Recently, three additional species of Sphinctrina with 1-septate spores have been recognized: S. ophioparmae Kalb, S. benmargana Selva, and S. palidella (Willey) Selva (Kalb Reference Kalb2001; Selva Reference Selva2004).
The four species of Sphinctrina hitherto known from the Iberian Peninsula have non-septate spores (Hladun & Llimona Reference Hladun and Llimona2007). However, several specimens previously identified as S. tubiformis or S. turbinata differ in terms of ecology and morphology. In addition to Iberian samples, similar specimens from other Mediterranean areas do not fit into currently described species of Sphinctrina. Therefore, a new species, Sphinctrina paramerae, is proposed to include all these samples.
Materials and Methods
The material of Sphinctrina examined includes Mediterranean and non-Mediterranean specimens. Fresh material was collected in several localities from the Iberian Peninsula, and is kept in the herbarium BCN. Dry material from several European herbaria (GZU, LEB, MACB, MA-LICH, MU, SALAF, SANT, UPS and VAB-LICH) was examined.
Thin and ultrathin sections of apothecia were examined using light microscopy. Thin sections of apothecia in calcium alginate were obtained using a freezing microtome. The inclusion in calcium alginate facilitates handling and orientation of the apothecia. Some sections were stained with lactic blue and cleaned with distilled water. Sections were dehydrated and fixed in DPX. Ultrathin sections were prepared at the Serveis Científicotècnics, Parc Científic, Universitat de Barcelona. Sections were stained with a 1% solution of methylene blue and 1% solution of sodium tetraborate and then heated for 30–60 s at 90°C.
Scanning electron microscope (SEM) examinations were carried out after fixing the samplers on a glue surface and coating with gold.
Chemical tests of apothecia were made on squash preparations mounted in water, applying a 10% aqueous solution of potassium hydroxide (K). Other reacting agents (nitric acid or bleach) were also used, but no reactions were obtained.
Measurements of apothecium height, capitulum width, stalk width, ascus length and width, and ascospore length and width follow Löfgren & Tibell (Reference Löfgren and Tibell1979): (absolute minimum) a-b (absolute maximum), where a represents arithmetic mean minus one standard deviation, and b represents arithmetic mean plus the standard deviation. Extreme values, arithmetic mean (χ), standard deviation (σ) and number of measurements (n) are given within parentheses.
Measurements of height and width of apothecia were made under a binocular lens on dry material. At least ten measurements of each sample were recorded when the material allowed.
Ascospore length, width, and length-breadth ratio were recorded from 38 specimens of S. turbinata, 9 of S. anglica, 10 of S. leucopoda, 12 of S. tubiformis, and 8 of S. paramerae, material permitting. All measurements were made with a×100 (1·80 NA) oil immersion lens on water-mounted squash material. At least twenty spores of each specimen were drawn with the aid of a drawing tube, avoiding irregular morphology and immature spores with a thick gelatinous coat. All the measurements were made using the ImageJ program (Abramoff et al. Reference Abramoff, Magelhaes and Ram2004). The mean of these measurements were used in all subsequent statistics to represent the specimens. Data did not fulfil requirements of normality and homogeneity of variance. A Kruskal-Wallis test including all species, as implemented in SPSS for Windows (version 14, SPSS Inc., Chicago, IL), was performed for each three characters to check the differences between species. As the Kruskal-Wallis tests show differences between the species for each character, a pairwise Mann-Whitney test was carried out for the three variables between species. It was assumed that individual specimens had been classified to species a priori. This classification was achieved using excipular structure, K reaction, presence or absence of a stalk, habitat, spore ornamentation and spore shape, among other characteristics. Graphical representations were carried out using R (R Development Core Team 2004).
The Species
Sphinctrina paramerae Muñiz & Hladun sp. nov.
MycoBank No: MB 564705
Lichenum parasiticum insidens Pertusaria paramerae. Apothecia exigua, 0·15–0·50 mm alta. Stipia non manifesta. Capitula sphaeroidea, parum depressa, nitida nigra praeter orbis margino albo. Excipulum spadiceum vel hyalinum cum margino xerampelino. Hypothecium brunneum. Asci (57·6) 63·4–74·2(80·6)×(7·2)8·1–9·5(10·1) µm lati, cylindrici. Sporae non septatae, fusciceae, sphaeroidae vel ellipsoideae, (7·9)8·9–11·7(14·4)×(6·5)7·8–9·3(10·7) µm, ornamento parum rugoso, involucro gelatinoso parum manifesto.
Typus: Spain, Catalonia, Tarragona, Terra Alta, Serra dels Ports, Arnes, Barranc del Grevolar, 31TBF7125, 800–900 m, on Pertusaria paramerae on Juniperus phoenicea, 20 October 1990, M. Boqueras (holotypus—BCC-Lich 6174).
(Fig. 1)

Fig. 1. Sphinctrina paramerae. A, habitus (BCN 2933); B, section of apothecium (UPS 41694); C, ultrathin section of exciple stained with methylene blue (MA 11266); D, detail of excipulum (UPS 41694); E, section of apothecium (SEM) (BCN 6174, holotype); F, detail of hypothecium (UPS 143231); G, spore ornamentation (SEM) (BCN 6174, holotype); H, spore ornamentation (light microscope) (BCN 6174, holotype). Scales: A=1 mm; B, E=100 µm; C, D & F=50 µm; G=10 µm; H=20 µm.
Apothecia sessile, (0·15)0·21–0·42(0·50) mm high, black or seldom dark brown. Capitulum (0·21)0·32–0·36(0·41) mm broad, spherical to vertically compressed, shining black, usually with a colourless excipular rim caused by loosely interwoven hyaline hyphae protruding beyond the coloured tissues. Mazedium black, well-developed. Excipulum with an outer scleroplectenchymatous layer (Yoshimura & Shinada Reference Yoshimura and Shimada1980), consisting of isodiametric to ovoid cells with thick gelatinous walls reddish brown to pale brown, almost hyaline inside; inner layer prosoplectenchymatous of periclinally arranged hyaline hyphae, 6–10 µm wide. Hypothecium medium to pale brown in section, formed by an upper layer of periclinally arranged thin hyphae and paraplectenchymatous below, cells with thin walls. Asci cylindrical, 8-spored, (57·6)63·4–74·2(80·6)×(7·2)8·1–9·5(10·1) µm. Spores uniseriate, non-septate, dark brown, elliptical to subspherical, (7·9)8·9–11·7(14·4)×(6·5)7·8–9·3(10·7) µm, (length: χ=10·02, σ=1·05, n=98; width: χ=8·57, σ=0·76, n=98). The distal and proximal ascospores have an ovoid shape. Ornamentation indistinct under the light microscope, slightly rugose in SEM. Spore coat very thin, seldom distinct in water.
Pycnidia not observed.
Chemistry
All reddish pigmented structures react with K intensifying the reddish brown colour. The pigment dissolves and stains the adjacent hyaline structures with a yellowish tinge.
Etymology
The name paramerae refers to the host, Pertusaria paramerae Crespo & Vêzda.
Ecology and habitat
Sphinctrina paramerae grows on Pertusaria paramerae. The host grows as an epiphyte on Juniperus thurifera in the most exposed areas of juniper forests of the Iberian high plateau (paramera). These habitats are characterized by a high environmental summer humidity and winter drought, with temperatures between 8–17°C at altitudes ranging between 800–1300 m. Pertusaria paramerae also occurs on J. phoenicea and J. oxycedrus at lower altitudes (Crespo & Vězda Reference Crespo and Vězda1985).
Distribution
Sphinctrina paramerae ranges from the temperate mesomediterranean to cold supramediterranean belts of the Iberian Peninsula, Morocco, Libya, Turkey and the FYROM.
Additional specimens examined
Libya: Cyrenaica: 65 km WSW of Derna, 24 km SW of Lamludah, on stems of Juniperus phoenicea in open J. phoenicea forest, 13 iii 1983, G. Thor [BCN 2933, UPS 41694 sub. Sphinctrina turbinata (Pers. Fr.) de Not.].—Morocco: Subiendo El Tigri-Mighkit, on Pertusaria sp., 11 iv 1987, A. Gómez-Bolea (BCN).—Spain: Cuenca: Serranía de Cuenca, Cotillas, 30TWK8738, 1200 m, on Pertusaria paramerae on Juniperus thurifera, 14 iii 1998, G. Aragón & I. Martínez (MA-Lichen 11249 sub. Sphinctrina tubiformis A. Massal.); Hoya de la Madre de las Latas, 30TWK9238, 1350 m, on Pertusaria paramerae on Juniperus thurifera, 14 iii 1998, G. Aragón & I. Martínez (MA-Lichen 11266 sub. Sphinctrina tubiformis A. Massal.). Tarragona: Terra Alta, Serra dels Ports, Arnes, Barranc del Grevolar, 31TBF7125, 800–900 m, on Pertusaria paramerae on Juniperus phoenicea, 20 x 1990, M. Boqueras [BCN 6174 sub. Sphinctrina turbinata (Pers.) De Not.].—Turkey: Antalya: Kaşa, transitus Sinekcibeli Geçidi in montibus Akdaglari, alt. 1500 m s.m., ad lignum trunci Juniperus sp.¸in thallo Pertusariae sp. vigens, 11 iv 1996, A. Vězda (UPS 143231).—FYROM: Makedonien: Galiĉica, Paß zw. Oteševo und Trpejca, Paßhöhe, um 1600 m, W-exponierte, auf Juniperus sp., 26 viii 1976, H. Mayrhofer (GZU sub. Sphinctrina cf. turbinata).
Discussion
Sphinctrina paramerae differs from other species of Sphinctrina with non-septate ascospores by having non-stalked apothecia. In addition, morphological features of ascospores supply good diagnostic characters for distinguishing between Mediterranean species of Sphinctrina (Table 1). Sphinctrina paramearae externally resembles S. turbinata and S. tubiformis (Fig. 1). However, S. paramerae can be easily separated from these species by anatomical characteristics of the apothecia, spore shape and spore size (Table 1). The main feature distinguishing S. paramerae from S. tubiformis is spore shape, the latter having almond-shaped verrucose ascospores. Many of the specimens of S. paramerae were previously misidentified as S. turbinata. Both species have a reddish pigment in the exciple that reacts with KOH; however, it turns reddish brown in S. paramerae and purple in S. turbinata. Sphinctrina paramerae has elliptical to subspherical spores, longer and wider than in S. turbinata (Fig. 2, Table 2). Sphinctrina paramerae also differs in the lack of a well-developed outer gelatinous spore coat which is present in S. turbinata. In addition, ascospores of S. turbinata have a distinct verrucose ornamentation, while S. paramerae has an irregularly ridged ornamentation.

Fig. 2. Comparison of spore length and width of Sphinctrina species with non-septate spores, represented as the mean value (circle) and 95% of confidence for each variable (whiskers).
Table 1. Comparison of main morphological characters of non-septate Sphinctrina species.
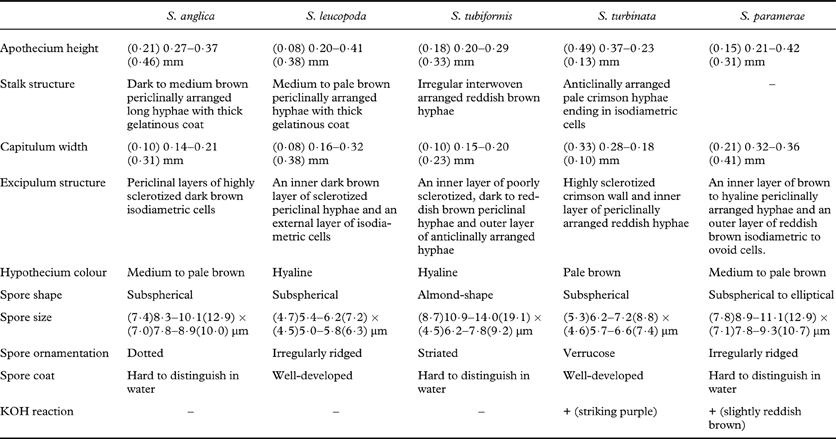
Table 2. Pairwise comparison of the species included in the non-septate group of Sphinctrina based on ascospore characters, using a Mann-Whitney test.

*significant at P<0·05; ns indicates no significant differences.
Sphinctrina paramerae could also be mistaken for S. anglica since spore dimensions are rather similar, although they are significantly longer in S. paramerae (Fig. 2, Table 2). The presence of a dotted ornamentation in the spores of S. anglica also distinguishes this species from S. paramerae. Additionally, S. paramerae differs from S. anglica in that the latter has stalked apothecia with a very different anatomy (Table 1). Sphinctrina paramerae differs from S. leucopoda in all the considered characters except the ornamentation of the spore, which consists of irregular ridges as seen in SEM and is almost smooth under light microscopy in both species.
Features on the morphology of ascospores of the species of Sphinctrina are useful as diagnostic characters. The presence or absence of a septum distinguishes two main groups of species. Among the species with non-septate spores, ascospore shape distinguishes S. tubiformis, with almond-shape ascospores from the species with subspherical to ellipsoidal ascospores: S. anglica, S. leucopoda, S. turbinata and the new species S. paramerae (Table 1). The comparison of spore size features of species with non-septate ascospores showed significant differences in terms of length and width (Table 2), the ascospores of S. paramerae being largest among the species with subspherical to ellipsoidal ascospores (Fig. 2). Among the species of Sphinctrina with non-septate ascospores, S. leucopoda and S. turbinata are the closest in terms of ascospore size (Fig. 2, Table 2). Both species can be separated by the colour of the apothecium. Sphinctrina turbinata has an unidentified crimson pigment reacting strikingly purple with K.
Key to the Mediterranean non-septate species of Sphinctrina
-
1 Ascospores almond-shaped ... S. tubiformis
Ascospores subspherical to ellipsoid ... 2
-
2(1) Ascospores 4·0–6·5 µm in length ... 3
Ascospores 7–12 µm in length ... 4
-
3(2) Apothecia K+ purplish ... S. turbinata
Apothecia K− ... S. leucopoda
-
4(2) Apothecia stalked, ascospores with a well-developed dotted ornamentation; exciple brown to dark brown in section, highly sclerotized ... S. anglica
Apothecia sessile; ascospores smooth or with irregularly ridged ornamentation; exciple colourless to pale brown in section, not sclerotized ... S. paramerae
We are grateful to the curators of the GZU, LEB, MACB, MA-LICH, MU, SALAF, UPS and VAL-LICH herbaria for loans of specimens cited in this investigation, and especially to our curator in BCN herbarium, A. Sánchez-Cuixart, for his infinite patience. We would like to express our gratitude to Dr A. Gómez-Bolea and Dra. G. Figueras for their help in the uncanny mission of finding fresh material. We are indebted to A. Romero and Dr J. M. Blanco for their valuable help with the statistical treatment of our data. The authors acknowledge support from the Spanish Ministerio de Educación y Ciencia (BOS 2001-0869-C04-02).




