



Introduction
With more than 400 species, the lichen genus Graphis is one of the largest genera of crustose lichens (Lücking et al. Reference Lücking, Archer and Aptroot2009, Reference Lücking, Johnston, Aptroot, Kraichak, Lendemer, Boonpragob, Cáceres, Ertz, Ferraro and Jia2014; Rivas Plata et al. Reference Rivas Plata, Hernández, Jesús, Lücking, Staiger, Kalb and Cáceres2011). The genus traditionally included taxa with lirellate ascomata and hyaline, transversely septate ascospores, but has been recircumscribed based on excipular structure and thallus morphology, as well as ascospore amyloidity (Staiger Reference Staiger2002). According to this concept, Graphis is also now defined as having ascomata with convergent labia, a partly to fully carbonized excipulum, a usually corticated, white-grey thallus, and amyloid ascospores that react I+ violet-blue. Graphis species are commonly found in montane and dry tropical forests in semi-exposed situations. Since the comprehensive work by Staiger (Reference Staiger2002), the number of species described within the genus has grown steadily due to the publication of a worldwide key (Lücking et al. Reference Lücking, Archer and Aptroot2009) and concerted efforts in taxonomic revisions and collection of the currently expanded family Graphidaceae (Sohrabi et al. Reference Sohrabi, Lücking and Lumbsch2014). As the genus appears to include two polyphyletic clades, the phylogenetic relationships within the genus remain an area of active research (Berger et al. Reference Berger, Stamatakis and Lücking2011; Rivas Plata et al. Reference Rivas Plata, Hernández, Jesús, Lücking, Staiger, Kalb and Cáceres2011, Reference Rivas Plata, Parnmen, Staiger, Mangold, Frisch, Weerakoon, Hernandez, Caceres, Kalb and Sipman2013).
The study of the genus Graphis in Thailand has been somewhat sporadic, even though the country contains many suitable habitats for the genus (Rundel & Boonpragob Reference Rundel and Boonpragob2009). Since the earliest report by Vainio in the Koh Chang Flora (Vainio Reference Vainio1909), 59 named species of the genus have been listed from Thailand, mostly as part of larger area-based studies of the family Graphidaceae, or the tribe Graphidoideae (Vainio Reference Vainio1909, Reference Vainio1921; Nakashi et al. Reference Nakashi, Kashiwadani and Moon2001; Aptroot et al. Reference Aptroot, Saipunkaew, Sipman, Sparrius and Wolseley2007; Papong et al. Reference Papong, Boonpragob and Lücking2007; Poengsungnoen et al. Reference Poengsungnoen, Mongkolsuk, Boonpragob and Manoch2010; Mongkolsuk et al. Reference Mongkolsuk, Bauruang, Polyiam, Vongshewarat, Phokaeo, Seeiam, Nirongbutr, Sangwisut and Sodamuk2011). In many cases, the genus represents the largest portion of the lichen diversity. For example, in a study of graphidoid lichens in Phu Luang Wildlife Sanctuary, Graphis constituted the largest genus, with over a third of the species richness (35 out of 91 species) in the family (Poengsungnoen et al. Reference Poengsungnoen, Mongkolsuk, Boonpragob and Manoch2010). Similarly, in a study of Khao Yai National Park, 34 Graphis species were reported, making it the most speciose genus in the study (Lichen Research Unit, Ramkhamhaeng University 2004). Despite this level of species diversity, a country-wide study on the genus has yet to be completed.
In the present paper, we aim to enhance the current understanding of the genus Graphis in the light of recent literature, collections and new molecular data. We describe one new species, along with six new records of Graphis from dry dipterocarp forests in northern and eastern floristic provinces of Thailand. We use morphological, anatomical, chemical, and molecular data to circumscribe the new species, Graphis koratensis Pitakpong, Kraichak & Lücking sp. nov.
Materials and Methods
Specimen collection and identification
A total of 1704 specimens belonging to Graphidaceae were collected during 2013 and 2014 from dry evergreen and dry dipterocarp forests in ten national parks and two other natural areas in Thailand. The national parks included 1) Khun Khan, 2) Doi Inthanon, 3) Phu Hing Rong Kla, 4) Phu Pha Terb, 5) Phu Toei, 6) Thong Pha Phum, 7) Phu Chong Na Yoi, 8) Pang Sida, 9) Keang Krachan, and 10) Hat Chao Mai. The two other natural areas included Sakaerat Environmental Research Station and Tadton waterfall in Mukdahan Province. The new species is described from specimens mostly collected at Sakaerat Environmental Research Station (SERS) in Nakhon Ratchasima (14°26' to 14°32'N, 101°50' to 101°57'E). SERS is located in mountainous terrain at 280–762 m above sea level, 300 km from Bangkok and 60 km from downtown Nakhon Ratchasima. All voucher specimens are deposited in the herbarium of Suranaree University of Technology (SUT), with some duplicates at the Field Museum (F).
The morphological and anatomical characteristics of the thallus and reproductive structures were studied using a low magnification stereomicroscope (Olympus-SZX12, Tokyo, Japan) at magnifications of ×7 to ×90. Sections of thalli and ascomata were examined with a light compound microscope (Olympus-BH2, Tokyo, Japan) at magnifications of ×40 to ×1000. All measurements were made on material mounted in water; Lugol’s iodine and 10% KOH solutions were used for the colour reactions of asci. Secondary products were identified by thin-layer chromatography in solvent C (170 ml toluene, 30 ml acetic acid), according to standardized methods (Elix & Ernst-Russell Reference Elix and Ernst-Russell1993; Orange et al. Reference Orange, James and White2001). The specimens were also examined under UV light (254 nm) for UV-reactive secondary metabolites.
Molecular data
DNA extraction, amplification, and sequencing of the mitochondrial small subunit (mtSSU), and nuclear large subunit (nuLSU) were performed using apothecia from four specimens of the new species. DNA was extracted using the Red-Extract Sigma Kit (Sigma Aldrich, USA). Dilutions of 9:1 of the genomic extracts were used in PCR reactions. Primers for PCR amplifications included a) mrSSU1 and mrSSU3R (Zoller et al. Reference Zoller, Scheidegger and Sperisen1999) for the mtSSU, and b) AL2R (Mangold et al. Reference Mangold, Martin, Kalb, Lücking and Lumbsch2008) and LR3 (Vilgalys & Hester Reference Vilgalys and Hester1990) for the nuLSU. PCR reactions contained 2·5 μl Sigma RED-Extract-N-Amp™ PCR, 0·5 μl of each primer (10 μM), 2 μl genomic DNA extract and 4 μl distilled water for a total of 10 μl. Thermal cycling conditions were as follows: a) for mtSSU: initial denaturation for 5 min at 95°C, followed by 35 cycles of 45 s at 94°C, 1 min at 50°C, 1 min 30 s at 72°C, and a final elongation for 10 min at 72°C; and b) for nuLSU: initial denaturation for 5 min at 94°C, followed by 35 cycles of 30 s at 95°C, 30 s at 58°C, 1 min at 72°C, and a final elongation for 10 min at 72°C. Amplification products were visualized on 1% agarose gels stained with ethidium bromide. The product was purified by the cutting of the target bands and digesting of the gel materials at 70°C for 10 min or until it turned liquid. Then it was cooled to 45°C-heat block for 5 min, treated with 1 μl of gelase (Epicentre Biotechnologies, Madison, WI, USA) and incubated at 45°C for at least 3 h or left overnight. The 10-μl cycle sequencing reactions followed standard protocols from a previous study (Rivas Plata et al. Reference Rivas Plata, Parnmen, Staiger, Mangold, Frisch, Weerakoon, Hernandez, Caceres, Kalb and Sipman2013). The products were then sequenced, using Applied Biosystems 3730 DNA Analyzer (Foster City, California, USA) automatic sequencer. Sequence fragments obtained were assembled with Geneious 8.0.3 (Drummond et al. Reference Drummond, Ashton, Buxton, Cheung, Cooper, Duran, Field, Heled, Kearse and Markowitz2014), manually inspected and adjusted. From this process, two mtSSU and three nuLSU sequences were obtained from the new species and submitted to GenBank (Table 1).
Table 1 Voucher information and GenBank accessions for the sequences used in the phylogenetic analysis. The accessions in bold are sequences generated in this study. The missing sequences are indicated with a dash (–)

Phylogenetic analysis
In order to determine the genetic identity of the new species, the newly obtained sequences were aligned with 42 other samples of lichens from the genus Graphis and two outgroup taxa of the genus Diorgyma available from GenBank (Table 1). For each locus, the alignment was performed in the software Geneious 8.0.3 (Drummond et al. Reference Drummond, Ashton, Buxton, Cheung, Cooper, Duran, Field, Heled, Kearse and Markowitz2014) with the built-in MUSCLE algorithm with eight iterations. The resulting alignment was further adjusted manually to remove ambiguity. The 956-bp concatenated sequences were used to reconstruct phylogenetic relationships among the samples with maximum likelihood (ML) and Bayesian approaches. The ML analysis was performed with RaXML BlackBox 8.1.1 (Stamatakis Reference Stamatakis2006), with two partitions for each of the two loci and 1000 pseudoreplicates. For the Bayesian approach, the dataset was also partitioned into two for each of the two loci and then analyzed using MrBayes 3.1.2 (Huelsenbeck & Ronquist Reference Huelsenbeck and Ronquist2001). A run with 10 000 000 generations, starting with a random tree and employing four simultaneous chains, was executed. Heating of chains was set to 0·2. Posterior probabilities were approximated by sampling trees, using a variant of the Markov chain Monte Carlo (MCMC) method. Every 1000th tree was sampled to avoid sample autocorrelation. The first 4000 trees were discarded as burn-in. For the remaining trees, a majority-rule consensus tree with average branch lengths was calculated using the sumt option of MrBayes. Posterior probabilities were obtained for each clade. Only clades with bootstrap support equal or above 70% under ML and posterior probabilities equal or above 0·95 in a Bayesian framework were considered as supported. Both ML and Bayesian analyses were performed on the CIPRES supercomputer cluster (Miller et al. Reference Miller, Pfeiffer and Schwartz2010). The topology from the ML analysis was illustrated, using the R-package ape (Paradis et al. Reference Paradis, Claude and Strimmer2004). The alignment and tree were submitted to TreeBase (http://purl.org/phylo/treebase/phylows/study/TB2:S17170).
Results and Discussion
After reviewing the literature, we found that 11 of the 59 reported names of Graphis from Thailand represented synonyms. Five of these were transferred to other genera, four with already reported Graphis species, and two with Graphis species new to Thailand (see Taxonomic Treatment below). From our recent collection of 1704 specimens, a total of 536 were identified as Graphis spp. Thirty-seven species of Graphis were found, including one new species, Graphis koratensis Pitakpong, Kraichak & Lücking sp. nov., and six new records from Thailand (see Taxonomic Treatment below). Graphis koratensis was collected from two dry dipterocarp forests in north-eastern Thailand. While morphologically similar to G. caesiella, the new species differs in a number of apothecial characters. It is also phylogenetically distinct from G. caesiella, with strong support from both the ML and Bayesian analyses (98 and 0·91, respectively; Fig. 1). Similar to a previous study (Berger et al. Reference Berger, Stamatakis and Lücking2011; Rivas Plata et al. Reference Rivas Plata, Hernández, Jesús, Lücking, Staiger, Kalb and Cáceres2011), we recovered two main clades, the Graphis and “Allographa” clades, within the genus in its current circumscription. While the monophyly of the genus Graphis is still somewhat uncertain (Rivas Plata et al. Reference Rivas Plata, Parnmen, Staiger, Mangold, Frisch, Weerakoon, Hernandez, Caceres, Kalb and Sipman2013), the analyses placed G. koratensis in the Graphis s. str. clade along with the type species, G. scripta.
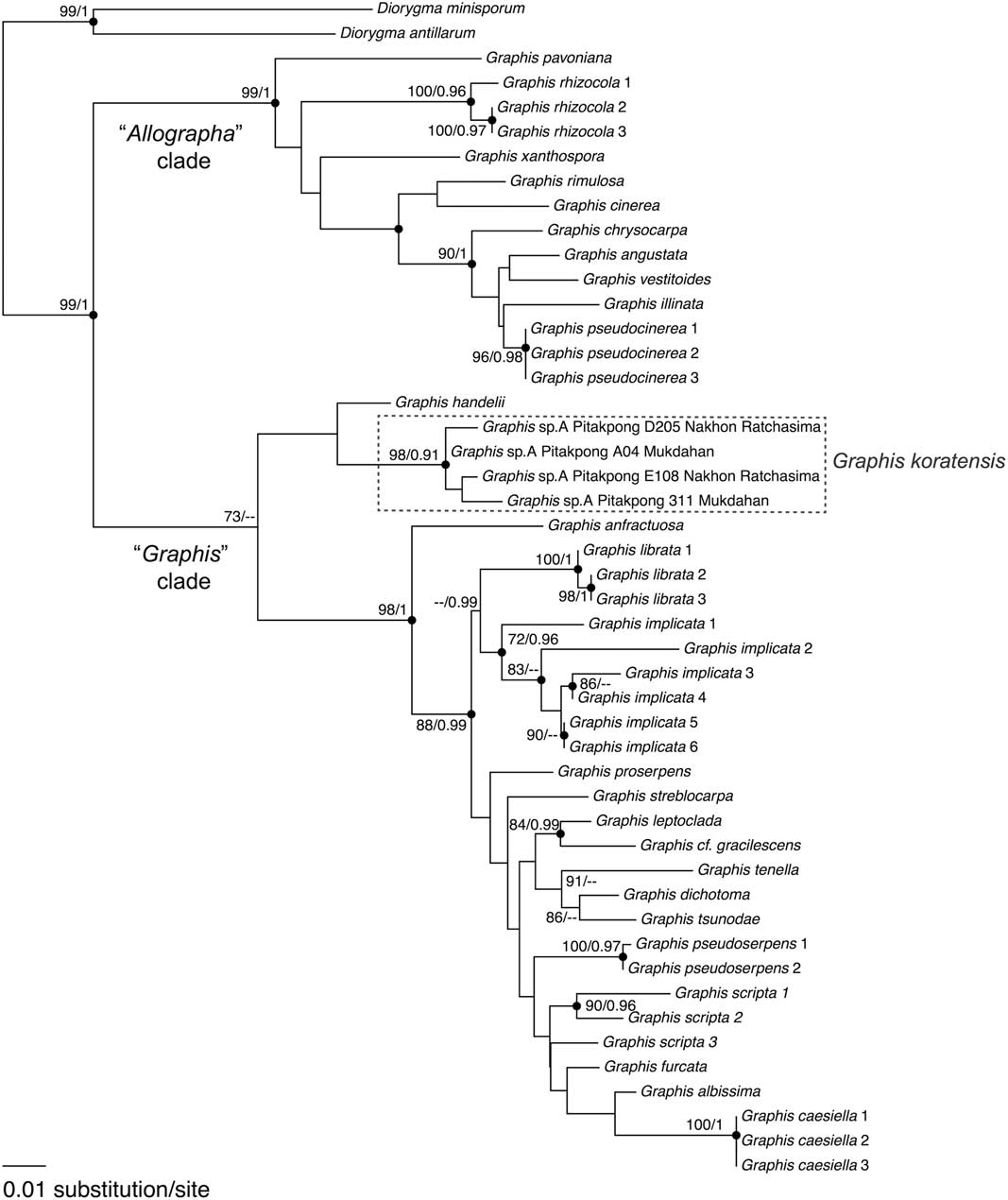
Fig. 1 Phenogram illustrating phylogenetic relationships among lichens in the genus Graphis. The topology follows the bipartitioned tree from a maximum likelihood (ML) analysis. The numbers at each node represent the bootstrap support value from the ML analysis, followed by the posterior probability (PP) from the Bayesian analysis (ML/PP). Only the ML support>70 and PP>0·95 are considered strong supports and reported here. Circles indicate nodes with strong support from both analyses.
Taxonomic Treatment
Graphis koratensis Pitakpong, Kraichak & Lücking sp. nov.
MycoBank No.: MB811731
Lirelline ascocarps with whitish grey or grey-green pruina along the slit with a completely carbonized expiculum.
Type: Thailand, Nakhon Ratchasima Province, Sakaerat Environmental Research Station, dry dipterocarp forest, located at a height of 380 m a.s.l., tree trunks, 2014, Pitakpong D205 (SUT—holotype; F—isotype).
(Fig. 2)

Fig. 2 Graphis koratensis Pitakpong, Kraichak, Lücking sp. nov. A & B, thallus and ascomata showing lirellae from Pitakpong D205; C, from Pitakpong A04; D & E, ascospores from Pitakpong D205. Scales: A–C=1 mm; D & E=10 µm.
Thallus corticolous, 3–6 cm diam., 150–250 µm thick in cross-section, continuous; surface smooth to uneven, pale whitish grey or grey-green, cortex distinct 10–15 µm; algal layer continuous 50–75 µm, medulla 100–150 µm thick, with clusters of calcium oxalate crystals.
Ascomata lirelliform, emergent to prominent, straight to curved, sparsely branched, with an apically thin thalline margin, 2–7 mm long, 0·3–0·5 mm wide, 0·10–0·15 mm high; labia thick, entire, appearing greyish black but with pruina along the slit. Proper exciple completely carbonized, 100–140 µm wide; hymenium clear, 100–150 µm high; asci 20–25 µm long, 110–140 µm wide. Ascospores 8 per ascus, oblong to narrowly fusiform, transversely 11–19 septate, 70–115 µm long, 9–15 µm wide, colourless.
Chemistry
Norstictic acid (thallus in section with K+ yellow efflux forming red, needle-shaped crystals, C−, P+ orange, UV−).
Etymology
The epithet was derived from the Thai word Korat, referring to the traditional name of the type locality (Nakhon Ratchasima).
Notes
This new species is similar to Graphis caesiella Vain. in having white-pruinose labia and elongate and irregularly branched lirellae, plus an identical chemistry, but differs in the completely carbonized excipulum and the larger ascospores (20−40×6−8 μm in G. caesiella). The only other species in the genus with a completely carbonized excipulum, large, transversely septate ascospores and norstictic acid is G. marginata Raddi, which has strongly prominent lirellae with sharply delimited, black labia, and belongs in the “Allographa” clade (Berger et al. Reference Berger, Stamatakis and Lücking2011).
In reference to the worldwide key (Lücking et al. Reference Lücking, Archer and Aptroot2009), G. koratensis belongs to Group 4 (labia entire, excipulum laterally carbonized, hymenium clear, ascospores transversely septate, p. 391–397). This species can be placed in couplet 6 along with Graphis erythrocardia Müll. Arg., for having a similar chemistry (norstictic acid), but differing in having larger ascospores.
Selected specimens examined. Thailand: Nakhon Ratchasima: dry dipterocarp forest in Sakaerat Environmental Research Station, 2014, Pitakpong E108, D202 (SUT). Mukdahan: dry dipterocarp forest in Phu Pha Terb, 16°26'7"N, 104°48'21"E, 2014, Pitakpong A04, 311 (SUT).
New records of Graphis from Thailand
Graphis cincta (Pers.) Aptroot
Habitat
Dry dipterocarp forest, Tadton waterfall, Mukdahan Province, SUT-211.
Graphis jejuensis K. H. Moon et al.
Habitat
Dry dipterocarp forest, Tadton waterfall, Mukdahan Province, SUT-315.
Graphis nigrocarpa Adaw. & Makhija
Habitat
Dry dipterocarp forest, Phu Pha Terb National Park, Mukdahan Province, SUT-005.
Graphis renschiana (Müll. Arg.) Stizenb.
Habitat
Mixed deciduous forest, Khun Khan National Park, Chiang Mai Province, SUT-501.
Graphis seminuda Müll.Arg.
Habitat
Dry dipterocarp forest, Tadton waterfall, Mukdahan Province, SUT-317.
Graphis subserpentina Nyl.
Habitat
Dry dipterocarp forest, Phu Chong Na Yoi National Park, Ubon Ratchathani Province, SUT-008.
Synonyms and nomenclatural notes
The following names were reported from Thailand, but have since been synonymized with other names and/or transferred to other genera. The names in bold are currently accepted names.
Graphis albidolivens Vain., Ann. Acad. Sci. Fenn. Ser. A, 15(6): 217 (1921), reported in Aptroot et al. (Reference Aptroot, Saipunkaew, Sipman, Sparrius and Wolseley2007)=Diorygma hieroglyphicum (Pers.) Kalb et al. in Symb. Bot. Ups. 34(1): 151 (2004).
Graphis aphanes Mont. & Bosch, Plant. Junghuhn. 4: 474 (1855), reported by Nakashi et al. (Reference Nakashi, Kashiwadani and Moon2001)=Hemithecium aphanes (Mont. & Bosch) M. Nakan. & Kashiw., Bull. Ntl. Sci. Mus. Tokyo 29: 88 (2003).
Graphis ceylanica Zahbr., Cat. Lich. Univ. 2: 297 (1923), reported in Papong et al. (Reference Papong, Boonpragob and Lücking2007)=Graphis dendrogramma Nyl. in Lücking et al., Lichenologist 41: 363–452 (2009).
Graphis chondroplaca (Redinger) Lücking et al., Fieldiana, Bot. 38: 64 (2008), reported by Poengsungnoen et al. (Reference Poengsungnoen, Mongkolsuk, Boonpragob and Manoch2010)=Graphis handelii Zahlbr. in Lücking et al., Lichenologist 41: 363–452 (2009).
Graphis concolor Nyl., Mem. Soc. Science. Nat. Cherbourg 5 (1857), reported in Vainio (Reference Vainio1909)=Graphina boschiana var. concolor (Nyl.) D.D. Awasthi & Kr.P. Singh, Current Science 42: 656 (1973); possibly a species of Carbacanthographis or Diorygma.
Graphis persimilis Vain., Bot. Tidsskr. 29: 125 (1909), reported in Vainio (Reference Vainio1909)=Phaeographis hypoglauca (Kremp.) Zahlbr., Cat. Lich. Univ. 2: 374 (1923).
Graphis pyrrhocheila Vain., Hedwigia 46: 179 (1907) nom. illeg., reported in Alava (1988)=Graphis pyrrhocheiloides Zahlbr. in Lücking et al., Lichenologist 41: 363–452 (2009).
Graphis siamensis Vain., Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo 1(3): 52 (1921), reported in Vainio (Reference Vainio1921); we were unable to locate the type material, but from the original description, this species does not belong to the genus Graphis, because of its ascoma shape and hymenium chemistry.
Graphis simplex Vain., Hedwigia 46: 77 (1907), reported in Vainio (Reference Vainio1907)=Graphis analoga Nyl. in Lücking et al., Lichenologist 41: 363–452 (2009).
Graphis subcinerea Staiger, Bibl. Lichenol. 85: 258 (2002) nom. inval., reported in Aptroot et al. (Reference Aptroot, Saipunkaew, Sipman, Sparrius and Wolseley2007)=Graphis phaeospora Vain. in Lücking et al., Lichenologist 41: 363–452 (2009).
Graphis tenuis Vain., Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo 1(3): 52 (1921), reported in Vainio (Reference Vainio1921)=Graphis hossei Vain. in Lücking et al., Lichenologist 41: 363–452 (2009).
This study was supported by grants from the Suranaree University of Technology, National Research Council of Thailand to AP, and a National Foundation of Science grant (DEB-1025861) to The Field Museum (Co-PIs: HTL and RL). All new sequences were generated in the Pritzker Laboratory for Molecular Systematics and Evolution at the Field Museum (Chicago).



