


Introduction
The Collemataceae (‘jelly lichens’) is a large lichen family having a cyanobacterial photobiont (Nostoc). A revised classification of the family distinguishes 10 genera, of which Leptogium (Ach.) Gray remains the largest, despite extensive species transfer to the re-erected genera Scytinium (Ach.) Gray and Pseudoleptogium Müll. Arg. (Otálora et al. Reference Otálora, Jørgensen and Wedin2014). Leptogium comprises, in its present form, c. 100 mainly epiphytic species from wet tropical to temperate regions of the world (Jørgensen Reference Jørgensen1973a , Reference Jørgensen1975, Reference Jørgensen1997; Otálora et al. Reference Otálora, Aragón, Martinez and Wedin2013).
Leptogium species with a Mallotium-type thallus (i.e. those having a tomentum of spherical- or cylindrical-celled hyphal hairs on one or both thallus surfaces) are retained in Leptogium in the classification of Otálora et al. (Reference Otálora, Jørgensen and Wedin2014), but were recovered as polyphyletic. While species with spherical-celled hyphal hairs, the focal group of the present study, form a monophyletic group, those with cylindrical-celled hairs occur in three distinct groups (Otálora et al. Reference Otálora, Aragón, Martinez and Wedin2013). Over 50 species with a Mallotium-type thallus are currently accepted (Kitaura & Marcelli Reference Kitaura and Marcelli2013), including a large number of recently described taxa (Jørgensen 1973 Reference Jørgensena , Reference Jørgensenb , Reference Jørgensen1997; Awasthi & Akhtar Reference Awasthi and Akhtar1977; Jørgensen & Kashiwadani Reference Jørgensen and Kashiwadani2008; Jørgensen & Olley Reference Jørgensen and Olley2010; Kitaura & Marcelli Reference Kitaura and Marcelli2012, Reference Kitaura and Marcelli2013; Kitaura et al. Reference Kitaura, Käffer, Marcelli and Martins2014, Reference Kitaura, Marcelli, de Hora and Jungbluth2015). Among these, only seven species with spherical-celled hairs are known to date (Kitaura & Marcelli Reference Kitaura and Marcelli2013), of which L. burgessii (L.) Mont., L. digitatum (A. Massal.) Zahlbr., L. hibernicum M. E. Mitch. ex P. M. Jørg., and L. laceroides B. de Lesd. are included in Macrolichens of East Africa (Swinscow & Krog Reference Swinscow and Krog1988). Leptogium ethiopicum C. W. Dodge (Dodge 1964) is treated as a possible synonym of L. burgessii in Jørgensen (1973 Reference Jørgensena ) and Aragón et al. (Reference Aragón, Otálora and Martínez2005), but has been recently accepted as a distinct species (Kitaura & Marcelli Reference Kitaura and Marcelli2013). The remaining two, L. mantiqueirense Kitaura & Marcelli and L. nylanderi Kitaura & Marcelli, were recently described from South America, whereupon the former was additionally reported for South Africa (Kitaura & Marcelli Reference Kitaura and Marcelli2013). The similarly neotropical L. velutinum P. M. Jørg. has hairs of an intermediate type, described as isodiametric at the base and short cylindrical towards the tips.
In lichen-forming fungi, a situation with high levels of homoplasy, few discrete morphological characters working at species level, and insufficient material available for study, has frequently resulted in broadly circumscribed and morphologically variable species with wide distributions. In recent years, a growing number of publications have demonstrated the presence of high genetic diversity and pseudocryptic lineages in such widespread species (e.g. the reviews by Crespo & Perez-Ortega Reference Crespo and Pérez-Ortega2009; Crespo & Lumbsch Reference Crespo and Lumbsch2010; Lumbsch & Leavitt Reference Lumbsch and Leavitt2011). Moreover, genetic divergence is often seen to be positively correlated with geographical distance or dispersal barriers (e.g. Argüello et al. Reference Argüello, Del-Prado, Cubas and Crespo2007; Sérusiaux et al. Reference Sérusiaux, Villarreal, Wheeler and Goffinet2011; Leavitt et al. Reference Leavitt, Esslinger, Divakar and Lumbsch2012, Reference Leavitt, Fernandez-Mendoza, Lumbsch, St. Clair, Sohrabi and Divakar2013; Amo de Paz et al. Reference Amo de Paz, Cubas, Crespo, Elix and Lumbsch2012; Del-Prado et al. Reference Del-Prado, Blanco, Lumbsch, Divakar, Elix, Molina and Crespo2013; Moncada et al. Reference Moncada, Reidy and Lücking2014; Alors et al. Reference Alors, Lumbsch, Divakar, Leavitt and Crespo2016).
Several specimens identified as L. hibernicum from extra-European countries were sequenced and compared with Norwegian and Western European specimens during a population genetic study of Leptogium hibernicum (M. Bendiksby, T. Bjelland, A. Frisch, M. H. Jørgensen & M. Görts, unpublished data). Initial analyses showed that specimens identified as L. hibernicum from East Africa form a strongly supported phylogenetic clade, clearly distinct from the species in Europe and eastern North America. After careful morphological examination, the East African material is described here under the name Leptogium krogiae and compared with other Leptogium species with spherical-celled hairs. We show the phylogenetic position of L. krogiae within the Collemataceae by Bayesian and maximum likelihood phylogenetic analyses of DNA sequence alignments containing both mitochondrial and nuclear DNA sequence data.
Material and Methods
Lichen sampling
Sequence data of the nuclear ribosomal internal transcribed spacer (nrITS) and large subunit (nrLSU), the mitochondrial ribosomal small subunit (mrSSU) and the DNA replication licensing factor mini-chromosome maintenance complex (Mcm7) were generated for 23 specimens identified as L. hibernicum from East Africa (Ethiopia, Kenya, Tanzania), Europe, and North and South America, and for one specimen of L. saturninum (Dicks.) Nyl. from Norway. The material was received on loan from the following herbaria: BG, BM, E, O, and S. Additional sequence data were added from GenBank, most of which were also used in previous phylogenies of the Collemataceae by Otálora et al. (Reference Otálora, Aragón, Martinez and Wedin2013, Reference Otálora, Jørgensen and Wedin2014; Table 1). Based on their results, we selected all specimens of taxa presumed to be close relatives of L. hibernicum (i.e. section Mallotium with spherical-celled hairs). These include L. hibernicum, L. digitatum, L. laceroides and L. velutinum. No DNA sequence data are yet available for L. burgessii, L. ethiopicum, L. mantiqueirense and L. nylanderi which are clearly differentiated morphologically from both L. hibernicum and L. krogiae. Our DNA sequencing effort was therefore focused on the putative new species and its closest relative, L. hibernicum. We also included a broad sampling from the Collemataceae and, based on Otálora et al. (2013, 2014), used Staurolemma omphalarioides (Anzi) P. M. Jørg. & Henssen (Pannariaceae) as the ultimate outgroup.
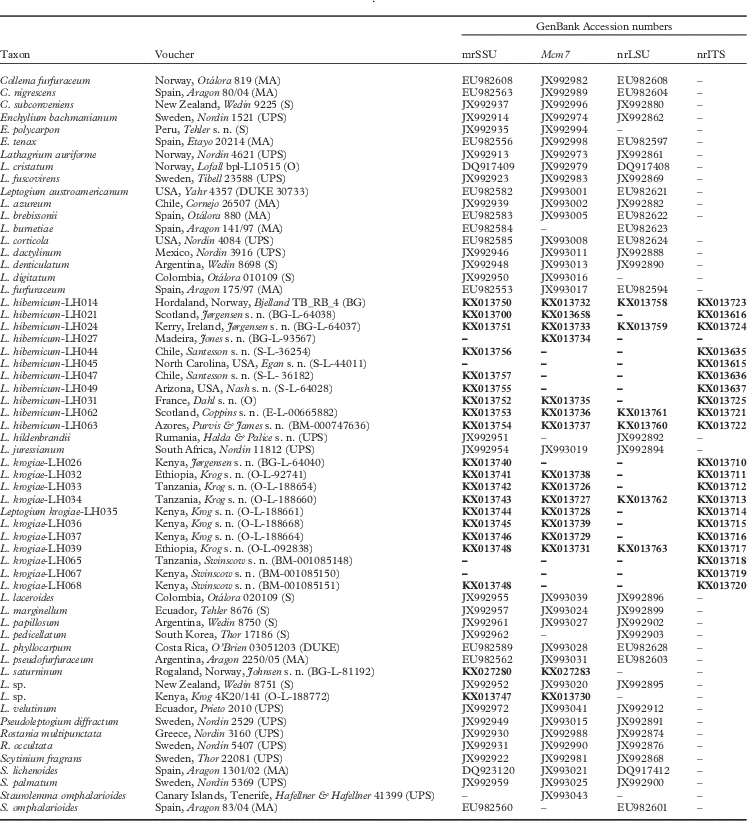
Table 1 Specimens used in this study with voucher information and GenBank Accession numbers. New sequences are shown in bold
DNA extraction and PCR amplification
The lichen samples were crushed using a Retsch TissueLyser II. Total genomic DNA was extracted using the E.Z.N.A. SP Plant DNA Mini Kit (Omega Bio-tek, Inc., Norcross, Georgia, USA) following the manufacturer’s instructions. The DNA regions were PCR amplified using PuReTaq Ready-To-Go PCR beads (GE Healthcare Life Sciences, Little Chalfont, Buckinghamshire, UK), in accordance with the manufacturer’s instructions. The following primer sets were used: 1) nrITS: ITS4 and ITS5 (White et al. Reference White, Bruns, Lee and Taylor1990), 2) nrLSU: LIC24R (Miadlikowska & Lutzoni Reference Miadlikowska and Lutzoni2000) and nrLSU-hypR2 (Bendiksby & Timdal Reference Bendiksby and Timdal2013), 3) mrSSU: mtSSU1 and mtSSU3R (Zoller et al. Reference Zoller, Scheidegger and Sperisen1999) and 4) Mcm7: our newly designed primers MCM7-FLH (GTCACTACGAAACAGTTCACC) and MCM7-RLH (GACGCCACCGATCAGGAGTAG). In older specimens, higher PCR success was achieved by amplifying shorter fragments using the following internal primers: 1) mrSSU: mtSSU-hypF and mtSSU-hypR (Bendiksby & Timdal Reference Bendiksby and Timdal2013) and 2) nrITS: ITS-lichF and ITS-lichR (Bendiksby & Timdal Reference Bendiksby and Timdal2013).
The samples were run on a BIO RAD T100 Thermal Cycler with the following settings: initial denaturation at 95°C for 10 min, and 31 (nrITS, nrLSU, mrSSU) or 35 (Mcm7) cycles at 95°C for 30 s, 60°C (nrITS, nrLSU, mrSSU) or 56°C (Mcm7) for 30 s and 72°C for 30 s, followed by a final extension at 72°C for 5 min. All PCR products including negative controls were visualized by electrophoresis on a 2% agarose gel. PCR products were purified with ExoProStar 1-step, Enzymatic PCR and Sequence Reaction Clean-up Kit (GE Healthcare Life Sciences) and sent for sequencing to Eurofins Genomics GmbH (Ebersberg, Germany) with the same primers as used for the PCR reaction.
Phylogenetic analyses
Sequences were aligned using the general MAFFT settings as implemented in the Guidance Web Server (Penn et al. Reference Penn, Privman, Landan, Graur and Pupko2010) and manually corrected. The nrITS region was included only for Leptogium hibernicum s. lat. (i.e. the specimens from Europe, the northern Atlantic and the Americas) and L. krogiae. This region is highly variable across the Collemataceae and available for only a small number of the species included in our phylogeny. By excluding all nrITS sequences outside the L. hibernicum-L. krogiae clade, we were able to make use of the full phylogenetic signal of the nrITS for our target species.
Prior to concatenation, we tested the four single-gene alignments for conflicting tree topologies. Bayesian Inference (BI) and Maximum Likelihood (ML) analyses were performed using the same settings for the single-gene and the concatenated four-gene alignments. Serious conflict was assumed when deviant tree topologies were supported by ≥70% bootstrap values (BS) and ≥0·95 posterior probabilities (PP). A partitioned dataset was used for the final phylogenetic analyses to enable independent parameter estimation for the four DNA regions. The Mcm7 alignment was further partitioned according to codon positions to allow for the higher evolutionary rates of the 3rd codon position.
The BI and ML phylogenetic analyses were performed on the CIPRES Science Gateway (Miller et al. Reference Miller, Pfeiffer and Schwartz2010). For the BI analysis, we used MrBayes 3.2.6 (Ronquist & Huelsenbeck Reference Ronquist and Huelsenbeck2003). A GTR-I-Γ model of sequence evolution was applied to the partitioned dataset and the model parameters were estimated during the run for each gene partition separately starting from a default flat Dirichlet distribution. The analysis was run for 15000000 generations in eight chains and every 500th generation was sampled. The first 50% of trees were discarded as burn-in and the PPs summarized on a 50% majority-rule consensus tree. For the ML analysis, we used the RAxML-HPC black box with rapid bootstrapping and full ML analysis under the GTR+GAMMA approximation (i.e. not allowing for a proportion of invariable sites, I). The analysis was automatically stopped after 252 bootstrap replicates using the bootstrapping option implemented in RAxML 3.2.7 (Pattengale et al. Reference Pattengale, Alipour, Bininda-Emonds, Moret and Stamatakis2009).
Morphology
The morphology of the lichen specimens was examined using a Zeiss stereomicroscope. Sections for anatomical examination were cut by hand, mounted in water, and examined with a Zeiss Axioskop 2 microscope. With the exception of the lobe and apothecium sizes, all measurements were made in water.
Results
Phylogenetic analysis
New sequences generated in this study included 21 nrITS, 6 nrLSU, 20 mrSSU, and 16 Mcm7 (Table 1).
The final alignment of 59 accessions and 2704 nucleotide positions contained 19 nrITS (453 nucleotide positions), 39 nrLSU (959), 55 mrSSU (815), and 48 Mcm7 (477) sequences, including the outgroup, Staurolemma omphalarioides.
There were no supported conflicts between the BI and ML analyses or between the separate single-gene analyses. We therefore performed the final analyses on a concatenated dataset of all four DNA regions which resulted in a well-resolved and overall strongly supported phylogenetic hypothesis (Fig. 1). All included accessions of L. hibernicum s. lat. group with high support. This clade contains two strongly supported subclades that correlate with geographical distance (Europe, the northern Atlantic and the eastern USA vs. Chile and the south-western USA; clades a and b, respectively). There is less genetic variation among the included accessions of L. krogiae, which forms a strongly supported sister (clade c) to L. hibernicum s. lat. Clade d comprises L. digitatum, L. laceroides, L. velutinum and two specimens collected as L. hibernicum from Kenya and New Zealand (Krogh 4K20/141 and Wedin 8751, respectively) that do not group with the other accessions of L. hibernicum or L. krogiae. Clade e, including subclades a, b, and c, together with its sister clade d, consists of species with spherical-celled hyphal hairs, with the exception of L. velutinum which has intermediate hairs.

Fig. 1 Phylogenetic relationships among Leptogium species with spherical-celled tomentum based on the best RAxML tree using a concatenated alignment of four genetic regions (nrITS, nrLSU, mrSSU, Mcm7) with particular focus on the phylogenetic position and genetic delimitation of Leptogium hibernicum s. lat (clades a+b) and the new species, L. krogiae (clade c; grey box). Thickened branches indicate Bayesian posterior probabilities PP ≥0·95 and RAxML bootstrap values BS ≥70%. Wide, light grey branches indicate BS ≥70% and PP <0·95. The dashed branch indicates PP ≥0·95 (BS <70%). The gap in the root branch spans 65% of the root branch length. Clades discussed in the text are indicated with the letter a–g. Species with cylindrical-celled tomentum are indicated with a black circle.
Our tree topology is largely consistent with previous studies by Otálora et al. (Reference Otálora, Aragón, Martinez and Wedin2013, Reference Otálora, Jørgensen and Wedin2014), except for the deviating position of Rostania Trevis., here falling out closer to the base of the tree (Fig. 1). Included specimens of Collema nest within Leptogium but the supporting branch receives PP<0·95. Species having a tomentum formed of hyphal hairs with cylindrical cells are found in three places in our tree topology: 1) L. juressianum Tav. being basally attached to the species with spherical-celled hyphal hairs, 2) L. furfuraceum (Harm.) Sierk, L. papillosum (B. de Lesd.) C. W. Dodge, and L. pseudofurfuraceum P. M. Jørg. & A. K. Wallace (clade f) and 3) L. burnetiae Dodge, L. hildenbrandii (Garov.) Nyl., L. pedicellatum P. M. Jørg., and L. saturninum (clade g).
The Species
Leptogium krogiae Bjelland, Frisch & Bendiksby sp. nov.
MycoBank No.: MB 817371
Leptogium species similar to L. hibernicum, but differing in the thinner thallus (190–210 µm vs. 390–480 µm), shallowly striate instead of deeply furrowed lobes, and only rarely distinctly flattened isidia. Apothecia stipitate with wrinkled pedicel, subhymenial euparaplectenchymatous layer, and excipular cortex of one cell layer. Ascospores submuriform, 32–38×11–14 µm, 8 per ascus.
Type: Tanzania, Iringa region, Iringa District, Mufundi escarpment, 8°40'S, 35°20'E, 1750–1800 m, August 1989, H. Krog no 4T 15/027 (O—holotype).

Fig. 2 A, Leptogium krogiae Bjelland, Frisch & Bendiksby, sp. nov. (O, holotype; photograph Lena Ohnheiser), apothecium indicated by arrow. B, Leptogium hibernicum s. lat clade (typical specimen) (see Fig. 1; photograph Einar Timdal). C, Leptogium delavayi, apothecium section (similar to L. krogiae). D, Leptogium pedicellatum, apothecium section (similar to L. burgessii and L. ethiopicum), showing different location of the euparaplectenchyma. Illustrations C and D from Asahina (Reference Asahina1935) with tissue descriptions and approximate scale bars superimposed. Abbreviations: ept=euparaplectenchyma, hym=hymenium, subhym=subhymenium, hyp=hypothecium. Scales: A & B=1 cm; C & D=c. 1 mm. In colour online.
Thallus brownish or bluish grey, dry lobes to 1 cm broad, wet lobes swollen to 190–210 µm thick; quadratic cells of cortex 5–6 µm thick; thallus plane to undulate, transversely wrinkled-striate only on the thickest portions, usually in the middle of the thallus; columnar hyphae present, straight to sinuose, 2·5–3·0 µm wide; isidia mainly laminal, scattered to agglomerated, grey to brown, granular to cylindrical to coralloid, occasionally flattened, and sometimes pubescent-tomentose close to the thallus surface. Lower surface paler, uniformly pubescent-tomentose; hyphal hairs unbranched to irregular branched; 80–110 µm long, with (5–)9–15 globose cells. Photobiont Nostoc; filaments often long (20–30 cells), c. 2·5 µm diam.
Apothecia very rare, up to 3 mm diam., stipitate; pedicel with longitudinal wrinkles in older apothecia; mature disc concave, red-brown; thalline exciple bearing lobules and sometimes numerous apical upright isidia (one specimen). Hymenium 180–220 µm high; subhymenium and hypothecium not clearly separated, 60 µm thick, yellowish; subhymenial euparaplectenchymatous layer 160–180 µm thick, of 10 cell layers (Fig. 2C); excipular cortex of one cell layer. Ascospores 8 per ascus, colourless, ellipsoid with acute ends, submuriform, 32–38×11–14 µm.
Etymology. Named after the late Norwegian lichenologist Hildur Krog to commemorate her major contributions to the exploration of East African lichens.
Ecology and distribution. Corticolous in mist-affected woodland and montane forest at altitudes of 1800–3300 m, scattered, not uncommon (Jørgensen 1973 Reference Jørgensena ; Swinscow & Krog Reference Swinscow and Krog1988; Krog Reference Krog1991, Reference Krog2000; Alstrup & Christensen Reference Alstrup and Christensen2006; all as L. hibernicum). Confirmed specimens have been seen from central Ethiopia through the Kenyan highlands to the Iringa region in south-western Tanzania. Leptogium krogiae seems to be common in the Taita Hills in south-eastern Kenya (J. Rikkinen, pers. comm.).
Notes. Leptogium krogiae (Fig. 2A) is morphologically close to L. hibernicum (Fig. 2B), with which it has been identified in the past (Jørgensen 1973 Reference Jørgensena ; Swinscow & Krog Reference Swinscow and Krog1988; Krog Reference Krog1991, Reference Krog2000; Alstrup & Christensen Reference Alstrup and Christensen2006). Both species share the brownish to bluish grey lobes with predominantly laminal isidia, short tomentum of spherical-celled hyphal hairs mainly on the lower surface, and lobe striation. While apothecia are present in some of the specimens of L. krogiae investigated, fertile specimens are unknown for L. hibernicum (Kitaura & Marcelli Reference Kitaura and Marcelli2013; M. Bendiksby, T. Bjelland, A. Frisch, M. H. Jørgensen & M. Görts, unpublished data). Morphological differences are mainly observed in the degree of striation of the upper lobe surface, lobe thickness, the length of the Nostoc filaments, and isidium shape.
In Leptogium krogiae, the striation is typically rather shallow and scattered compared to the deeply furrowed lobes of L. hibernicum. According to P. M. Jørgensen (pers.com.), the degree of striation of the lobes may reflect the thallus thickness which, when wet, is 390–480 µm in L. hibernicum (Aragón et al. Reference Aragón, Otálora and Martínez2005) and 190–210 µm in L. krogiae. We have also observed marked differences in the length of the Nostoc filaments, which on average consist of 12–16 cells in L. hibernicum (Aragón et al. Reference Aragón, Otálora and Martínez2005) and 20–30 cells in L. krogiae. The shape of the isidia in L. krogiae varies from granular to cylindrical to coralloid. The isidia in L. krogiae are more rarely flattened compared to those in L. hibernicum.
No DNA sequence data are yet available for L. burgessii, L. ethiopicum, L. mantiqueirense and L. nylanderi, but these species are morphologically clearly differentiated from both L. hibernicum and L. krogiae. Neither L. burgessii, L. ethiopicum, L. mantiqueirense nor L. velutinum have isidia. Furthermore, L. burgessii differs from L. krogiae in the absence of a subhymenial euparaplectenchymatous layer (see Fig. 2C vs. 2D), the thick euparaplectenchymatous amphithecium, thinner lobes (40–50 µm) and a smooth to slightly rugulose surface. Leptogium ethiopicum was considered a possible synonym of L. burgessii (Jørgensen 1973 Reference Jørgensena ) but was recently accepted as an independent taxon (Kitaura & Marcelli Reference Kitaura and Marcelli2013). The type and only specimen of the species from the Chokke Mountains in Ethiopia (FH-Dodge!) is difficult to interpret, but clearly differs from L. krogiae in the absence of a subhymenial euparaplectenchymatous layer (see Fig. 2C vs. 2D). Further differences include the thick euparaplectenchymatous amphithecium, thinner lobes (80–90 µm), absence of striation on the upper lobe surface, shorter tomentum (10–20 µm) and presence of degraded lobules on the rim of the apothecial margin rather than isidia.
Leptogium velutinum differs from L. krogiae in the absence of isidia and a subhymenial euparaplectenchymatous layer, thinner lobes (50–55 µm), a smooth surface (rugulose under ×20 magnification) and a velvety lower side, and the tomentum hairs have isodiametric cells at their base and cylindrical cells at their apex (Kitaura et al. Reference Kitaura, Käffer, Marcelli and Martins2014). Leptogium nylanderi differs from L. krogiae by having a subhymenial colloplectenchymatous layer, a euparaplectenchymatous amphithecium and an apothecium with granular isidia and thinner lobes (70–90 µm) (Kitaura & Marcelli Reference Kitaura and Marcelli2013). Leptogium laceroides differs from L. krogiae in the absence of a subhymenial euparaplectenchymatous layer, thinner lobes (70–100 µm) and by having a smooth surface (Kitaura & Marcelli Reference Kitaura and Marcelli2013).
Only two additional Leptogium species with both spherical-celled hairs and a thick subhymenial euparaplectenchymatous layer (Fig. 2C) are described at a global level. 1) Leptogium digitatum is easily distinguished by the deeply ridged lobes bearing a dense tomentum on both surfaces, while the tomentum is largely confined to the lower lobe surfaces in L. krogiae; isidia and lobules are absent and the ascospores are smaller (25–30×10–13 µm). 2) Leptogium mantiqueirense differs from L. krogiae in the non-isidiate and non-lobulate thallus with smooth margins, larger apothecia (to 6 mm diam.) bearing a basally fused collar of lobules at the edge of the otherwise smooth margin, smaller ascospores (22–30×10–13 µm) and anatomical details such as a thinner thallus (100–190 µm), shorter tomentum (5–20(–35) µm), lower subhymenium (12–25 µm), lower hymenium (110–150 µm) and a subhymenial euparaplectenchymatous layer (85–125 µm). A comparison of diagnostic characters of Leptogium species with spherical-celled hyphal hairs is presented in Table 2.
Table 2 Taxonomic characteristics of Leptogium section Mallotium with spherical-celled or velvet-like hairs (authors’ data; Kitaura & Marcelli Reference Kitaura and Marcelli2013; Kitaura et al. Reference Kitaura, Käffer, Marcelli and Martins2014)

Selected specimens examined. Ethiopia: Sidamo (Shewa) Province: Wondo Gennet, 20 km S of Shashemenne, elev. 1800–2000 m, on trees in mixed forest, i 1972, Krog s. n. (O); between Shashemenne and Kofele, elev. 2400 m, on trees in mixed forest, i 1972, Krog s. n. (O); ibid., Krog E27/36 (O). Arussi Province: W slope of Mt. Chilalo, E of Asella, elev. 3000–3100 m, on trees in Hagenia–Hypericum forest, i 1972, Krog E30/41, E30/43, E30/44 (O).—Kenya: S slope of Mt. Kenya, Arundinaria alpina bamboo thicket ab. Kamaweti Forest Station, elev. 2400–2700 m, on Nuxia trees, 29 xi 1971, Pócs 6601/K & Fadew (BG). Rift Valley Province: Elgeyo Marakwet District, Chebiemit forest 1 km W of Cheptongei, 0°57'N, 35°29'E, elev. 2300 m, on trees, 1973, Krog 2K11/121 (O); ibid., forest 2 km NE of Kapcherop, 1°04'N, 35°20'E, elev. 2200 m, on fallen forest tree, 1973, Krog 2K14/113a (O); Kajiado District, Ngong Hills, 1°27'S, 36°37'E, elev. 2300 m, on shrubs and rocks, 1972, Krog K45/141 (O). Central Province: Mt. Elgon, S of Suam River valley to Kapchalwa Gate, 1°15'N, 34°50'E, elev. 2500–3100 m, 1973, Ryvarden 9360 (O); Nyeri District, Mt. Kenya Safari Club, 0°02'S, 37°07'E, elev. 2100 m, 1976, Krog 4K19/143 (O); Nyeri District, Mt. Kenya, S slope, Regati Forest Station, 0°20'S, 37°15'E, elev. 2200–2300 m, 1973, Ryvarden 9827 (O); Kirinyaga District, Mt. Kenya, near Castle Forest Station, 0°23'S, 38°18'E, elev. 1900 m, on trees, 1972, Krog K49/128 (O); Kirinyaga District, Aberdare Mts., 10 km W of Tusha, ericaceous zone, 0°31'S, 36°44'E, elev. 2900 m, on trees, 1974, Krog 3K31/184 (O); Kirinyaga District, Mt. Kenya, 2 km NW of Irangi Forest Station, moist deciduous forest near River Ena, 0°20'S, 37°28'E, 2000 m, 1972, Krog K48/152, K48/153, K48/154 (O); Kirinyaga District, Mt. Kenya, by Thiba Fishing Camp, 0°24'S, 37°18'E, elev. 1850 m, on tree, 1972, Krog K52/116 (O). Eastern Province: Meru District, Mt. Kenya, open forest on E side of Themwe, 0°09'S, 37°34'E, elev. 2100 m, on rocks and trees, 1974, Krog 3K16/288 (O).—Tanzania: Tanga Province: Lushoto District, West Usambara Mts., along boundary of Mazumbai University Forest Reserve, in montane rainforest between Sagara Ridge and motor road, 4°49'S, 38°30'E, elev. 1500–1900 m, 1988, Krog 2T13/6 (O); ibid., Usambara Mts., Magamba c. 4 km N of Lushoto, 4°50'S, 38°15'E, elev. 1600–2000 m, 1973, Ryvarden 10922 (O); ibid., West Usambara Mts., Mazumbai University Forest Reserve, Sagara Ridge, 4°49'S, 38°30'E, elev. 1850–1980 m, in mossy elfin forest and Philippia heath with rock outcrops, 1988, Krog 2T09/18 (O); ibid., along motor road from Mazumbai University Forest Station to the edge of the forest reserve, 4°48'S, 38°30'E, elev. 1500–1560 m, 1988, Krog 2T08/023 (O). Southern Highlands Province: Iringa District, Mufindi escarpment, 8°40'S, 35°20'E, elev. 1750–1800 m, 1989, Krog 4T15/027, 4T15/28 (O); Iringa District, Buffalo Hill at edge of Mufindi escarpment, on trees in montane rainforest and at forest edge, 8°40'S, 35°20'E, elev. 1850–1900 m, 1989, Krog 4T10/168, 4T10/170, 4T10/173 (O).

Discussion
We found three distinct phylogenetic lineages within L. hibernicum having discrete distributions (Fig. 1: clades a–c). Leptogium hibernicum in the strict sense (clade a) in our study is confined to Western Europe, some islands of the northern Atlantic (Azores, Madeira) and eastern North America. An extended study on the phylogeography of L. hibernicum in Western Europe (M. Bendiksby, T. Bjelland, A. Frisch, M. H. Jørgensen & M. Görts, unpublished data) supports this pattern. Three specimens from Chile and the south-western USA form a second lineage (clade b). We have not studied these specimens in detail, and more material is needed for drawing solid taxonomic conclusions. However, our preliminary phylogenetic results suggest that this clade represents one or more as yet unrecognized species closely related to L. hibernicum s. str. (clade a).
A third strongly supported and distinct lineage (clade c) is described here as L. krogiae. We have seen a large number of specimens of this species from East Africa but a wider distribution within Africa is suggested by reports of L. hibernicum from southern Africa (Swinscow & Krog Reference Swinscow and Krog1988) and the availability outside East Africa of habitats suitable for the species. Another example of geographically disjunct lineages within Leptogium species was given recently by Otálora et al. (Reference Otálora, Martínez, Aragón and Molina2010), who recovered the populations of L. furfuraceum from Europe and L. pseudofurfuraceum from North America as sister clades, as was the case with L. furfuraceum from Africa and L. pseudofurfuraceum from South America.
Further extra-European specimens of L. hibernicum included in our study proved unrelated to that species though they all fall within the clade covering species with a spherical-celled tomentum (Fig. 1). Leptogium sp. Krogh 4K20/141 from Kenya is morphologically close to L. laceroides (P. M. Jørgensen, pers. com.) but differs genetically from a second specimen of that species in our phylogeny collected in Colombia which is geographically closer to the type locality of L. laceroides in Mexico. Leptogium sp. Wedin 8751 from New Zealand, included as L. hibernicum in the studies of Otálora et al. (Reference Otálora, Aragón, Martinez and Wedin2013, Reference Otálora, Jørgensen and Wedin2014), was not available for study.
In the classification of Otálora et al. (Reference Otálora, Jørgensen and Wedin2014), the selected lectotype of Leptogium, L. lacerum (Ach.) Gray (=L. lichenoides (L.) Zahlbr.), falls within the reinstated genus Scytinium (Jørgensen et al. Reference Jørgensen, Otálora and Wedin2013). In order to sustain taxonomic stability and avoid the need for recombining the bulk of Leptogium species into Mallotium (Ach.) Gray, Jørgensen et al. (Reference Jørgensen, Otálora and Wedin2013) proposed conserving Leptogium with a new type, L. azureum (Sw. ex Ach.) Mont. We prefer to follow the present classification and describe the new species in Leptogium rather than Mallotium until a decision is made on this proposal.
The authors thank the curators at BG and O for the loan of specimens and for giving permission to remove small tissue samples for DNA extraction (BG and O). We also thank Prof. Per Magnus Jørgensen (MNH, UoB) for useful discussions and comments on our manuscript. This project was partly financed by a grant from the County Governor of Hordaland to TB, and partly by NTNU (equal rights funding) to MB.




