



Introduction
For hundreds of years, only four species of Graphis (in the sense of Lücking et al. (Reference Lücking, Archer and Aptroot2009)) were known from Europe. Seven species have since been added (Lepista & Aptroot Reference Lepista and Aptroot2016), all recently collected in Portugal. Since 2016, some species of the genus Graphis have been relocated to the phylogenetically somewhat distant genus Allographa (Lücking & Kalb Reference Lücking and Kalb2018). This includes some species treated below and those names are cross-referenced here.
Over the last few years, the first author has continued to study Graphis in Portugal where this genus is abundant and extremely diverse, compared with other regions in Europe (Lepista & Aptroot Reference Lepista and Aptroot2016). In recent years we have studied over 100 specimens of this genus from an area in Portugal called Mata Nacional do Buçaco (MNB) (Fig. 1). All 11 European species of Graphis were already known from Portugal, either from the Serra de Sintra and/or the Planalto das Cezaredas. In MNB these 11 species were also found and, in addition, several Graphis specimens were collected that clearly did not belong to any reported species.

Fig. 1. Images of Mata Nacional do Buçaco (MNB). A, ‘Porta de Sula’, one of the entrances to MNB. B, a large specimen of Dicksonia antarctica, located in the ‘Vale dos Fetos’ (Valley of Ferns). C & D, two views of the climax forest of Buçaco, dominated by Quercus trees. E, detail of the leaves of Ilex aquifolium, a common species at MNB. F, a very old exotic tree species, located in the gardens inside MNB. In colour online.
Mata Nacional do Buçaco is a region in Portugal located c. 40 km from the Atlantic coast, 20 km from Coimbra and 200 km from Lisbon. It occupies a part of the Sierra of the same name, Serra do Buçaco, also known as ‘Serra do Luso’, ‘Serra de Carvalho’, ‘Serra de Santo António’ or ‘Serra do Cântaro’. Administratively it lies in the village of Luso in the Mealhada municipality in the district of Aveiro. The MNB is located between 40°22ʹ15ʺ and 40°23ʹ01ʺN and 8°21ʹ26ʺ and 8°22ʹ30ʺW.
The climate in this area as measured in detail in 2007 (Veiga 2014; Fundação Mata do Buçaco Reference Mata do Buçaco2015) is super-humid mesothermal, with moderate water deficit in the summer and small thermal efficiency at the same time. The average annual rainfall is 1525 mm, with 137 days/year with precipitation equal to or greater than 0.1 mm and 54 days/year with precipitation equal to or greater than 10 mm. The maximum monthly rainfall is 225 mm in December, with the month of January the second wettest with 221 mm. The driest month is August with 20.4 mm of rainfall, followed by July with 25 mm. The annual average temperature is 14.0 °C, with a maximum of 21.4 °C in August and a minimum of 7.6 °C in January, which translates into an annual temperature range of 13.8 °C. The annual average maximum temperature is 18.2 °C, with a maximum of 27.4 °C in August and a minimum of 10.3 °C in January. The average annual minimum temperature is 9.7 °C, with a maximum of 15.5 °C in August and a minimum of 4.8 °C in January. The absolute maximum temperature was recorded in August 2007 at 40.4 °C. The registered absolute minimum temperature was −3.1 °C in February 2007. The annual frequency of occurrence of the minimum number of days with temperatures below 0 °C is 3.7, with 2.0 days in January. The frequency of annual occurrence of the number of days with maximum temperatures above 25 °C is 67, with a maximum of 20.5 days in August and zero days in the months November through to March. The annual average insolation is 1718 hours of sun, with a maximum of 276 in July and a minimum of 38 in January. The annual average relative air humidity is 80.2%, with a maximum of 88.4% in November and a minimum of 71.0% in July.
Three of the natural habitats of the Habitats Directive (European Commission 1992) are present in the study area, all located in the climax forest at Cruz Alta: 1) laurel forest, located on the northern slopes and north-eastern climax forests, a subtype of the priority habitat arborescent matorral with Laurus nobilis (5230); 2) Carvalhais Galician-Portuguese Quercus robur and Q. pyrenaica forest, located on the northern slope of the climax forests and a subtype of the priority habitat Mediterranean deciduous forest (9230); 3) thermo-Mediterranean and pre-desert scrub (5330), located on the west and north-west slope, forming a large part of the climax forests where there is a clear dominance of Phillyrea latifolia.
The natural forest of that area, of which there are still some patches (e.g. near the Cruz Alta) is primarily comprised of Arbutus unedo, Erica arborea, Ilex aquifolium, Laurus nobilis, Olea europaea var. sylvestris, Phillyrea latifolia, Pyrus communis, Quercus robur and Viburnum tinus.
The trees in the MNB support a moderately rich lichen flora, mainly of the Lobarion and Graphidion communities with, for example, four species of Lobaria and Sticta limbata.
Here, we report five Graphis species found recently in MNB as new to Portugal and even to the whole of Europe. All newly reported species are described and keyed out below. The specimens cited here have been assigned to pantropical species that were previously described but not recorded from Europe (Lücking et al. Reference Lücking, Archer and Aptroot2009). It brings the total number of Graphis species known from Europe to 16 (Lepista & Aptroot Reference Lepista and Aptroot2016). This number includes the three species now treated in Allographa. All of these occur in Portugal and nine are, within Europe, known exclusively from Portugal (see below).
Material and Methods
Identification and descriptive work were carried out in Amadora using an AmScope T490A compound microscope, and in Soest using an Olympus SZX7 stereomicroscope and an Olympus BX50 compound microscope with differential interference contrast, connected to a Nikon Coolpix digital camera. Sections were mounted in tap water, in which all measurements were taken. Some of the specimens were tested in the field for the reaction with KOH (10–15%), but for all specimens the reaction to KOH was analyzed under the compound microscope, using a mount of a thin section of the thallus in water to which a drop of KOH was added. Lirella morph terminology mostly follows Lücking et al. (Reference Lücking, Archer and Aptroot2009). Descriptions are based on cited material from Portugal only. All specimens mentioned were collected in 2015 or 2016, and are preserved in LISU.
The Species
Graphis dendrogramma Nyl.
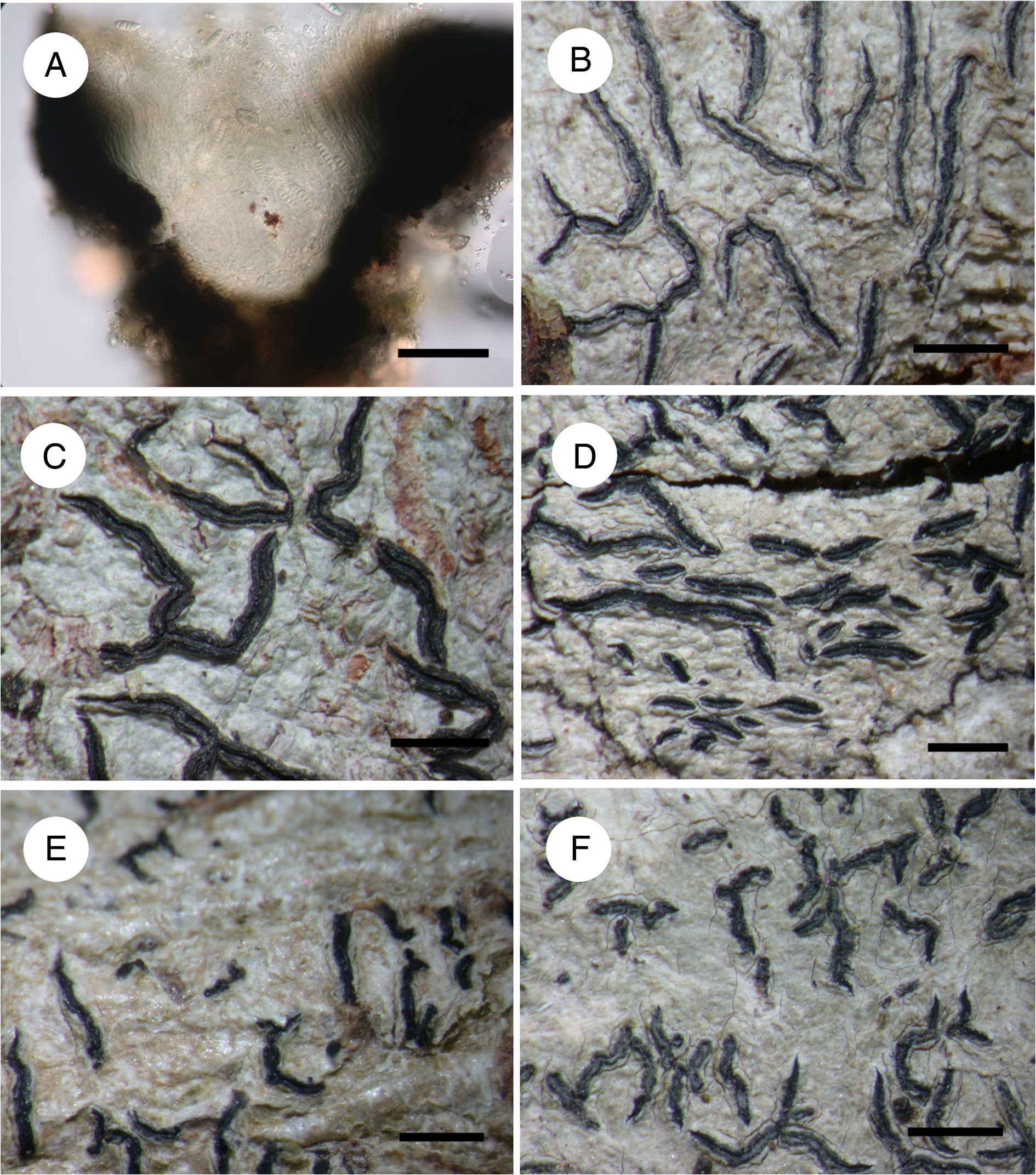
Fig. 2. Graphis species from Mata Nacional do Buçaco. A & B, Graphis dendrogramma (Zaca 9245 p.p.). A, transverse section through apothecium. B, habitus. C, G. duplicata (Zaca 9269), habitus. D, G. gonimica (Zaca 9245 p.p.), habitus. E, G. librata (Zaca 9574), habitus. F, G. pyrrhocheiloides (Zaca 9395), habitus. Scales: A = 50 μm; B–F = 1 mm. In colour online.
Thallus KOH+ yellow (stictic acid). Lirellae dendrogramma-morph, no striation, excipulum laterally carbonized, disc closed, labia pruinose, hymenium not inspersed. Ascospores 6–9-septate, 29–45 × 6.5–8.5 μm, (3.5–)4–5(–6) times as long as wide (n = 13).
Specimen examined
Portugal: Mata do Buçaco, 2015, Zaca 9431, 9245 p.p.
Graphis duplicata Ach.
(Fig. 2C)
Thallus KOH−. Lirellae duplicata-morph, with striation, excipulum laterally carbonized, disc closed, labia not pruinose, hymenium not inspersed. Ascospores 7–11-septate, (30.2–)31.2–42.7(–49.3) × (6.9–)7.3–8.8(–9.9) μm, (3.8–)4–4.9(–5.6) times as long as wide (n = 18).
Specimens examined
Portugal: Mata do Buçaco, 2015, Zaca 9269; 2016, Zaca 757.
Graphis gonimica Zahlbr.
(Fig. 2D)
Thallus KOH+ yellow → red. Lirellae lineola-morph, no striation, excipulum completely carbonized, disc closed, labia not pruinose, hymenium inspersed. Ascospores 5–10-septate, (21.2–)23.4–29.2(–33.2) × (6.3–)6.6–7.8(–8.7) μm, (3.1–)3.3–4.1(–4.4) times as long as wide (n = 132).
Specimens examined
Portugal: Mata do Buçaco, 2015, Zaca 9245 p.p., 9250; 2016, Zaca 579, 761, 769, 785, 796, 836, 854, 892.
Graphis librata C. Knight
(Fig. 2E)
Thallus KOH+ yellow → red. Lirellae lineola-morph, no striation, excipulum laterally carbonized, disc closed, labia not pruinose, hymenium not inspersed. Ascospores 7–9-septate, (27.5–)32.9–43.4(–44) × (6.6–)7–8.5(–8.8) μm, (3.9–)4.1–5.4(–6.4) times as long as wide (n = 31).
Specimens examined
Portugal: Mata do Buçaco, 2015, Zaca 9418, 9481, 9510, 9574; 2016, Zaca 682, 747, 756.
Graphis pyrrhocheiloides Zahlbr.
(Fig. 2F)
Thallus KOH+ yellow → red. Lirellae scripta-morph, no striation, excipulum laterally carbonized, disc open, pruinose, labia not pruinose, hymenium not inspersed. Ascospores 7–10-septate, (27.5–)31.6–39.3(–41.4) × (6.6–)6.9–7.9(–8.2) μm, (4–)4.3–5.3(–5.5) times as long as wide (n = 37).
Specimens examined
Portugal: Mata do Buçaco, 2015, Zaca 9395; 2016, Zaca 598.
Key to the species of Allographa (A.) and Graphis (G.) known from Europe (all also present in Portugal)
1 Ascospores only transversely septate………2
Ascospores muriform………15
2(1) Lirellae becoming striate; hymenium never inspersed………3
Lirellae never striate (try both options for young material); hymenium often inspersed with oil droplets………6
3(2) Ascospores (small to) medium-sized (30–60 μm long), 7–13-septate………4
Ascospores medium-sized to large (27–100 × 7.5–11.5 μm), 10–15-septate, not conspicuously thick-walled (wall < 2 μm thick); thallus KOH+yellow → red………5
4(3) Ascospores conspicuously thick-walled (wall usually > 2 μm thick); thallus KOH+ yellow → red………G. elegans
Ascospores not conspicuously thick-walled (wall < 2 μm thick); thallus KOH−………G. duplicata
5(3) Excipulum completely carbonized, also below the hymenium………A. leptospora (reported as G. leptospora)
Excipulum only laterally carbonized, open below………A. verminosa (reported as G. verminosa)
6(2) Hymenium not inspersed………7
Hymenium inspersed with oil droplets………10
7(6) Thallus KOH−; disc often open and pruinose………G. scripta complex
Thallus KOH+ yellow or yellow → red………8
8(7) Thallus KOH+ yellow; disc closed; labia pruinose………G. dendrogramma
Thallus KOH+ yellow → red………9
9(8) Disc open………G. pyrrhocheiloides
Disc closed………G. librata
10(6) Thallus KOH−; disc closed; labia not pruinose; lirellae erumpent, with lateral thalline margin………G. lineola
Thallus KOH + yellow → red; other characters variable (see below)………11
11(10) Disc open; lirellae erumpent, with lateral thalline margin………12
Disc closed; lirellae variable………13
12(11) Disc white-pruinose………G. crebra
Disc not pruinose………G. handelii
13(11) Labia white-pruinose; lirellae immersed………G. plumierae
Labia not pruinose; lirellae erumpent………14
14(13) Excipulum completely carbonized, also below the hymenium………G. gonimica
Excipulum only laterally carbonized, open below………G. cincta
15(1) Apothecia emergent; disc becoming open………G. inustuloides
Apothecia superficial; disc closed………A. ruiziana (reported as G. ruiziana)
Discussion
Lepista & Aptroot (Reference Lepista and Aptroot2016) gave a lengthy discussion of the unique Graphis biota of Portugal, together with possible explanations. The present results, with this additional region included, add substantially to the degree of uniqueness of Graphis in Portugal. One possible explanation for the diverse, somewhat tropical Graphis assemblage in Portugal is climate and/or climate change, combined with the presence of park-like forest with some ecological continuity or at least old trees and high air humidity (Fig. 1). However, it should also be pointed out that in the area of investigation many exotic tree species were planted more than a century ago (Lopes Reference Lopes2012). It cannot be ruled out that some of these trees were brought from more tropical places and that some of the lichens were already present on the young trees. Since c. 1920, the region has been known to contain the only mainland European localities of several other tropical lichen species, notably the related Glyphis cicatricosa Ach.
The climate in the mountains of Portugal is characterized as ‘Csb’ in the system of Köppen (Geiger Reference Geiger, Bartels, ten Bruggencate, Hausen, Hellwege, Schäfer and Schmidt1954), as used in the Times Atlas of the World: subtropical (temperature in hottest month between 20 °C and 22 °C), and dry (< 60 mm m−1) in summer. It is the only region in Europe with this climate type, which also occurs on the American West Coast (California), the SW coast of Chile (Santiago and surroundings), South Africa (around Cape Town), southern Australia (SE of Perth) and central Iran. From this exhaustive list it can already be seen that this climate type is rather infrequent, not to be confused with a Mediterranean climate, which is the usual climate in southern Europe (and elsewhere). Apparently, the Graphis species present here can occur under more varied climatic regimes, but not under any other occurring in Europe, where, for example, none of the humid subtropical or tropical climates occur.
All of the newly reported species, as well as the Graphis species reported in Lepista & Aptroot (Reference Lepista and Aptroot2016), are essentially pantropical. It is still a mystery whether these species arrived relatively recently or have been present for a long time. One way to investigate this would be to restudy herbarium specimens from the last century. Unfortunately, only a small number of Graphis specimens seem to have been collected in the last century in mainland Portugal. An online search through the databases of some major herbaria yielded only a handful of specimens, as compared to hundreds from countries such as Germany. The LISU herbarium is temporarily closed but also contains only a small amount of relevant material. The reason may have been that there were few lichenologists in Portugal in the last century, and that because only two Graphis species purportedly occurred in Europe, it was not deemed useful to collect many specimens of them. Very recently, two of the species reported by Lepista & Aptroot (Reference Lepista and Aptroot2016) from Portugal have been found in Great Britain. Also, it should be noted that in formerly polluted lowland regions of western Europe, all Graphis species are currently rapidly spreading onto areas and trees where they did not occur in previous centuries (see e.g. NDFF 2022).
The question remains why, as far as we know among the predominantly tropical crustose lichens, are only the graphidoid Graphidaceae (Glyphis, Graphis, Phaeographis) in Portugal unusually species-rich by European standards. At this time, we have no ready explanation for this. Perhaps further sampling in the region focusing on other groups might still reveal some surprises. It is for instance notable that the biota of another essentially tropical genus, viz. Pyrenula, in central Iran (one of the areas with a similar climate) was found to be surprisingly rich in species for a mostly dry subtropical country (Aptroot et al. Reference Aptroot, Valadbeigi and Sipman2012). Based on this, we can almost predict a similar Pyrenula diversity in Portugal.
It should also be pointed out that the current species concept in the genus Graphis is based on a combination of morphological, anatomical and chemical characters. While this is an improvement on older concepts focusing mostly on ascospores, it still provides challenges, as shown by the complex situation of the Graphis scripta species aggregate (Neuwirth & Aptroot Reference Neuwirth and Aptroot2011; Kraichak et al. Reference Kraichak, Lücking, Aptroot, Beck, Dornes, John, Lendemer, Nelsen, Neuwirth and Nutakki2015). Therefore, care must be taken in drawing substantial biogeographical conclusions when it is not precisely known that the same species is being considered. This is highlighted by the identification of the material named G. dendrogramma above, which does not fit the typical morphology of the species, although it shares the key characters. Neither is the material called G. gonimica a perfect fit. It may well be that some of the species have no available names, and this could be tested only using molecular approaches involving the addition of representative tropical material.
Author ORCID
André Aptroot, 0000-0001-7949-2594.


