



En América el estudio de los perros (Canis familiaris = Canis lupus familiaris) hallados en sitios arqueológicos ha permitido discutir, con variable grado de profundidad, su época de arribo al continente y las potenciales rutas de dispersión seguidas hasta la Patagonia, su variabilidad geno/fenotípica, sus roles en la economía, la organización social y el sistema de creencias amerindias, entre otros (e.g., Berón et al. Reference Berón, Prates and Prevosti2015; Fiedel Reference Fiedel2005; Mitchell Reference Mitchell2017; Prates, Berón y Prevosti Reference Prates, Berón, Prevosti, Berón, Luna, Bonomo, Montalvo, Aranda and Aizpitarte2010; Valadez et al. Reference Valadez, Mendoza and Blanco2004; Witt et al. Reference Witt, Judd, Kitchen, Grier, Kohler, Ortman, Kemp and Malhi2015). En Norteamérica los perros se registran desde el Holoceno temprano (ca. 10.000-9000 años cal aP; Perri et al. Reference Perri, Widga, Lawler, Martin, Loebel, Farnsworth, Kohn and Buenger2019), mientras que en Sudamérica son más recientes. El registro más confiable es de finales del Holoceno medio y proviene del sitio arqueológico Real Alto en Ecuador, con un contexto datado por seriación cerámica anclada con fechados de aproximadamente 4350-4050 años cal aP (Solomon y Stahl Reference Solomon and Stahl2011:621). Ya para el Holoceno tardío los registros son más frecuentes y tienen una amplia distribución que abarca a Perú, Bolivia, Chile, Argentina, Brasil y Uruguay (Acosta et al. Reference Acosta, Loponte and Esponda2011; Allison et al. Reference Allison, Focacci and Santoro1982; Belotti López de Medina Reference Belotti López de Medina2017; López Mazz et al. Reference López Mazz, Moreno, Bracco and González2018; Mendoza España Reference Mendoza España2013; Milheira et al. Reference Milheira, Loponte, Esponda, Acosta and Ulguim2017; Prates, Berón y Prevosti Reference Prates, Prevosti and Berón2010; Vásquez et al. Reference Vásquez, Rosales, Gálvez and Dorado2016; entre otros). La fecha taxón más antigua de Sudamérica corresponde al sitio arqueológico Ancón en Perú, con una antigüedad de aproximadamente 3080-2740 años cal aPFootnote 1 (Brothwell et al. Reference Brothwell, Malaga and Burleigh1979:157).
Desde su ingreso al continente americano y durante milenios, los perros fueron el único animal domesticado entre las poblaciones indígenas. Esta situación se mantuvo en muchos sectores de América del Sur, fuera del área andina, hasta la conquista europea en los siglos quince y dieciséis. Los perros indígenas fueron documentados por primera vez por Cristóbal Colón en su diario de viaje, sólo cinco días después de pisar tierra americana en 1492 (Colón [1492] en Fernández de Navarrete Reference Fernández de Navarrete1858:184). Desde el área Circum-Caribe hasta la Patagonia Austral, los registros arqueológico, etnohistórico y etnográfico muestran los distintos roles que tuvieron los perros para innumerables poblaciones amerindias, que además poseían palabras específicas para llamarlos en sus lenguas nativas. Entre estos diversos roles se destacan los siguientes: fueron una ayuda fundamental para la caza, la pesca y el pastoreo; se consideraron bienes de intercambio; se emplearon para el transporte de toldos y objetos; actuaron en batallas; se usaron como fuente de calor corporal; su carne sirvió como alimento en ocasiones especiales; sus pieles y pelos tejidos fueron usados para manufacturar vestimentas y mantas; sus dientes se usaron para collares y adornos y su cráneo para instrumentos sonoros; participaron de mitos, danzas y rituales mortuorios y de curación de enfermedades, y también en ceremonias religiosas en las que se los sacrificaba y comía; fueron representados en la cerámica, en objetos de metal y en el arte rupestre, entre otros (e.g., Allen Reference Allen1920; Cabrera Reference Cabrera1934; Coiazzi Reference Coiazzi1914; Gallardo Reference Gallardo1964–1965; Latcham Reference Latcham1922; Mendoza España Reference Mendoza España2013; Palermo Reference Palermo1988; Schwartz Reference Schwartz1997; Vásquez et al. Reference Vásquez, Rosales, Gálvez and Dorado2016).
Una revisión de la obra antropológica reunida en los seis volúmenes del Handbook of South American Indians (Steward Reference Steward1946–1950), complementada con otra bibliografía etnográfica, muestra los repetidos usos que tenían los perros, desde momentos inmediatamente posteriores a la llegada de los conquistadores europeos en el siglo dieciséis hasta tiempos más recientes en los siglos diecinueve y veinte. En las tierras bajas del Caribe, el delta del río Orinoco, el Chaco y la Patagonia, los perros eran considerados bienes personales a los que se mataba y enterraba junto a sus dueños (Belaieff Reference Belaieff, Steward and Steward1946:379; Cooper Reference Cooper and Steward1946a:156, 166; Fitz-Roy Reference Fitz-Roy1839:167; Kirchhoff Reference Kirchhoff and Steward1948a:877, Reference Kirchhoff and Steward1948b:226). También se registran de manera frecuente en contextos funerarios, acompañando entierros humanos en la costa norte de Chile, el desierto de Atacama, el noroeste de Argentina y Perú (Bennett Reference Bennett, Steward and Steward1946:607; Bird Reference Bird, Steward and Steward1946a:592; Cabrera Reference Cabrera1934:82–83; Larco Hoyle Reference Larco Hoyle and Steward1946:150). En la Amazonía fueron mascotas apreciadas y tratadas como personas, donde podían ser amamantados desde cachorros por las mujeres y también ser enterrados como los seres humanos (Horton Reference Horton and Steward1948:273). En el Chaco sus huesos intervenían en rituales mortuorios y su carne cruda era el alimento de los curanderos toba y wichí (Métraux Reference Métraux and Steward1946:329, 360). En Paraguay los chalupíes sacrificaban perros, previamente alimentados con la carne de una persona muerta, y luego los enterraban junto al difunto (Susnik Reference Susnik1983:64).
Los perros también se usaban para el acecho y la caza de diversas aves y mamíferos terrestres, como jaguares (Panthera onca) por los guaraníes (Métraux Reference Métraux and Steward1948a:81), o pecaríes (Tayassuidae) y tapires (Tapirus terrestris) por los kaingang (Basile Becker Reference Basile Becker1976:180; Métraux Reference Métraux and Steward1946:451–452). Otros registros etnográficos muestran que además se utilizaban para la obtención de mamíferos que viven en ambientes acuáticos, como por ejemplo coipos (Myocastor coypus) en el caso de los chaqueños (Métraux Reference Métraux and Steward1946:260), o lobitos de río patagónicos (Lontra provocax) entre los yámanas (Fitz-Roy Reference Fitz-Roy1839:187). En las montañas de Ecuador y Perú y la Amazonía se emplearon como señuelo o cebo para cazar jaguares (Métraux Reference Métraux and Steward1948b:704; Steward y Métraux Reference Steward, Métraux and Steward1948:609). En los llanos de Mojos, durante las inundaciones, se empleaban en una estrategia de caza en la que se rodeaban las islas donde se refugiaban los cérvidos y a las que se entraba con manadas de perros, así las presas atemorizadas corrían al agua donde las esperaban los cazadores en sus canoas (Métraux Reference Métraux and Steward1948c:412). Incluso eran especialmente entrenados para nadar y bucear por poblaciones que vivían en ambientes acuáticos, como por ejemplo los chonos (Cooper Reference Cooper, Steward and Steward1946b:51) y los alakaluf (Bird Reference Bird, Steward and Steward1946b:60, 62), cuyos perros en el agua conducían a los peces hacia las redes. Por último, es interesante mencionar que, para algunas poblaciones sudamericanas, los perros eran símbolos de riqueza y prestigio, y podían intercambiarse (Fitz-Roy Reference Fitz-Roy1839:151, 172; Latcham Reference Latcham1922:42, 47; Steward y Métraux Reference Steward, Métraux and Steward1948:543). Por ejemplo, los cara de las sierras del norte de Ecuador intercambiaban sus perros, sal y mantas por achiote, loros, monos y niños traídos de las tierras bajas del este (Murra Reference Murra and Steward1946:794).
Desde el punto de vista arqueológico, antes de conocerse la nueva información presentada en este artículo, la evidencia disponible para Sudamérica sugería que la mayor densidad de C. familiaris estuvo restringida a los Andes centro-sur y al altiplano (Ecuador, Perú, Bolivia, Chile y Argentina), y vinculada a sociedades agro-pastoriles con alto grado de sedentarismo y complejidad social institucionalizada (Allison et al. Reference Allison, Focacci and Santoro1982; Mendoza España Reference Mendoza España2013; Prates, Prevosti y Berón Reference Berón, Gutiérrez, Nigris, Fernández, Giardina, Gil, Izeta, Neme and Yacobaccio2010). En contraposición, hay algunas regiones que no presentan hasta el momento hallazgos arqueológicos prehispánicos confirmados de esta especie, como por ejemplo Amazonía, Gran Chaco, Sierras Centrales, centro-oeste de Argentina y sur de Chile.
En Argentina, el hallazgo de C. familiaris en contextos prehispánicos resulta cada vez más frecuente. Este taxón ha sido registrado sin ambigüedades en las regiones Noroeste, Noreste, Pampeana y Norpatagonia (Belotti López de Medina Reference Belotti López de Medina2017; Cabrera Reference Cabrera1934; Day Pilaría Reference Day Pilaría2018; Loponte y Acosta Reference Loponte and Acosta2016; Prates, Berón y Prevosti Reference Prates, Berón, Prevosti, Berón, Luna, Bonomo, Montalvo, Aranda and Aizpitarte2010). En este marco, este artículo tiene como objetivo discutir el rol de los perros entre las sociedades indígenas que vivieron en los ambientes fluviales de los ríos Paraná y Uruguay durante el Holoceno tardío. Para ello se presentan ocho especímenes óseos y dentarios correspondientes a C. familiaris, procedentes de cinco sitios arqueológicos de las provincias de Santa Fe y Entre Ríos, ubicadas en la región Noreste de Argentina (NEA). De los especímenes estudiados sólo uno había sido publicado y asignado con reservas a un perro (Salemme et al. Reference Salemme, Tonni, Ceruti, Iriondo and Cione1987); los siete restantes, determinados por primera vez en este trabajo, fueron hallados en excavaciones sistemáticas y en revisiones de las colecciones del Museo Arqueológico Manuel Almeida (MAMA) de Gualeguaychú, y del Museo de Ciencias Naturales y Antropológicas Profesor Antonio Serrano (MAS) de Paraná, ambos de la provincia de Entre Ríos.
Materiales y métodos
Los especímenes estudiados proceden de los siguientes sitios arqueológicos: La Lechuza (LZA, ca. 1751-672 años cal aP; Cornero Reference Cornero2016, Reference Cornero, Politis and Bonomo2018; NISP = 1), Arroyo Las Mulas 1 (ALM1, ca. 1060-623 años cal aP; Ceruti Reference Ceruti2003; Cornero Reference Cornero2019; NISP = 1), La Palmera V (LPV, ca. 667-513 años cal aP; Salemme et al. Reference Salemme, Tonni, Ceruti, Iriondo and Cione1987; NISP = 1), Sambaquí de Puerto Landa (SPL, ca. 1071-524 años cal aP; Almeida Reference Almeida1983; Castro y Del Papa Reference Castro and Papa2015; Verzi et al. Reference Verzi, Olivares, Hadler, Castro and Tonni2018; NISP = 3) y Cerros de los Pampas (CP; Almeida Reference Almeida1983; NISP = 1). Se suma un espécimen procedente del río Uruguay inferior (Entre Ríos; NISP = 1), registrado en el MAMA y cuya procedencia no está determinada con exactitud dentro del departamento Gualeguaychú (Figura 1 y Tabla 1).

Figura 1. Procedencia de los especímenes de Canis familiaris mencionados en el artículo (triángulos: nuevos registros presentados en este estudio; círculos: registros previos). 1: La Lechuza. 2: Arroyo Las Mulas 1. 3: La Palmera V. 4: Sambaquí de Puerto Landa. 5: Cerros de los Pampas. 6: MAMA-s/p-A. 7: Cerro Mayor. 8: Cerro Lutz. 9: La Argentina. 10: Cerro Farall. 11: Anahí. 12: La Bellaca 2. 13: Las Marías. 14: Chenque 1. 15: Angostura 1. 16: PSG-07. 17: La Yeguada. 18: Puntas de San Luis. 19: CH2D01. 20: Potrerillo de Santa Teresa. 21: Cráneo Marcado. (Color en la versión electrónica)
Tabla 1. Especímenes estudiados de Canis familiaris y dataciones radiocarbónicas obtenidas.

Los especímenes fueron recuperados en posición estratigráfica asociados con otros restos faunísticos, junto a restos botánicos, cerámicos y líticos. En la mayoría de los sitios arqueológicos de procedencia de las muestras analizadas también se recuperaron restos óseos humanos (Texto Suplementario 1), aunque no hay evidencias de que los perros hayan formado parte de contextos funerarios humanos. En estos sitios arqueológicos se desarrollaron actividades domésticas que incluyeron la elaboración de vasijas, la manufactura de instrumentos de hueso o piedra, la preparación y consumo de alimentos, y a veces también inhumaciones humanas. Excepto La Palmera V,Footnote 2 los demás se vinculan principalmente con la entidad arqueológica Goya-Malabrigo (sensu Politis y Bonomo Reference Politis and Bonomo2012, Reference Politis and Bonomo2018), asociada con poblaciones canoeras y constructoras de montículos, que vivieron en el curso medio e inferior del río Paraná e inferior del río Uruguay, desde hace aproximadamente 2000 años aP hasta el siglo diecisiete. Estas poblaciones, identificadas en las crónicas con distintos etnónimos (chaná, timbú, mbeguá, corondás, quiloazas, mocoretás, entre otros), basaron su economía en la caza, la pesca, la recolección y la horticultura a pequeña escala de maíz, poroto y zapallo. También representaron parte de los animales del entorno en la alfarería con los característicos apéndices zoomorfos de aves, mamíferos, reptiles y moluscos.
La determinación anatómica y taxonómica de los especímenes se realizó mediante la comparación morfológica con la colección de Mastozoología del Centro Regional de Investigaciones Científicas y Transferencia Tecnológica (La Rioja) y con bibliografía especializada (Morey y Wiant Reference Morey and Wiant1992; Prates, Berón y Prevosti Reference Prates, Berón, Prevosti, Berón, Luna, Bonomo, Montalvo, Aranda and Aizpitarte2010; Prevosti et al. Reference Prevosti, Santiago, Prates and Salemme2011; Tedford et al. Reference Tedford, Taylor and Wang1995; entre otros). En adición, se hicieron análisis morfométricos en dientes (Prevosti y Lamas Reference Prevosti and Lamas2006), hemimandíbula y poscráneo (von den Driesch Reference von den Driesch1976). Cada una de las medidas consideradas es un promedio de tres mediciones hechas con un calibre de 0,01 mm de precisión marca Mitutoyo. Los valores fueron comparados con los de distintos perros prehispánicos publicados para el Cono Sur. Además, se incorporaron valores de una muestra comparativa de cánidos silvestres sudamericanos, actuales y paleontológicos del Holoceno (Chrysocyon brachyurus —aguará guazú, Dusicyon avus —zorro extinto, Cerdocyon thous —zorro de monte, Lycalopex gymnocercus —zorro gris), procedentes de Argentina (Prevosti Reference Prevosti2006; Prevosti et al. Reference Prevosti, Ramírez, Schiaffini, Martin, Sauthier, Carrera, Sillero-Zubiri and Pardiñas2015).
Para la estimación de la edad de muerte de los individuos se utilizaron las categorías etarias para perros propuestas en Morris (Reference Morris2011). Para la mandíbula se consideró la erupción dentaria (Silver Reference Silver, Brothwell and Higgs1969) y el grado de desgaste del m1 (Horard-Herbin Reference Horard-Herbin and Crockford2000) e incisivos y caninos (Piérard Reference Piérard1967), y para los huesos largos, la fusión epifisiaria (von Pfeil y DeCamp Reference von Pfeil and DeCamp2009). La estimación sexual se realizó a partir de rasgos cualitativos de la fosa masetérica (Shigehara et al. Reference Shigehara, Onodera, Eto and Crockford1997). La estimación de la masa corporal de los individuos se realizó por medio de fórmulas de regresión del m1 (Legendre y Roth Reference Legendre and Roth1988), hemimandíbula (Losey et al. Reference Losey, Osipov, Sivakumaran, Nomokonova, Kovychev and Diatchina2015) y poscráneo (Losey et al. Reference Losey, McLachlin, Nomokonova, Latham and Harrington2017). Las categorías de tamaño (pequeño, mediano y grande) se usan de acuerdo con los estándares de razas actuales.
Paralelamente, con el objetivo de acceder a la historia de vida de los animales (Losey et al. Reference Losey, Bazaliinskii, Garvie-Lok, Germonpré, Leonard, Allen, Katzenberg and Sablin2011; Morris Reference Morris2011), y también a la historia tafonómica de los especímenes, se relevaron paleopatologías, evidencias de meteorización, depositación química, marcas de raíces, de roedores y de carnívoros, huellas de corte, de impacto y de machacado y tipos de fracturas (e.g., Behrensmeyer Reference Behrensmeyer1978; Binford Reference Binford1981; Chrószcz et al. Reference Chrószcz, Janeczek, Bielichová, Gralak and Onar2015; Fernández-Jalvo y Andrews Reference Fernández-Jalvo and Andrews2016). Por último, se obtuvieron tres nuevas fechas taxón por medio de dataciones radiocarbónicas (AMS).
Resultados
La muestra estudiada se compone de especímenes craneales (NISP = 7) y poscraneales (NISP = 1) y en conjunto representan un MNI = 7 (Tabla 1 y Figura 2). Los análisis basados en criterios morfológicos y morfométricos permitieron discriminar los especímenes estudiados de las especies de cánidos silvestres sudamericanos y asignarlos con precisión a C. familiaris (Texto Suplementario 1, Tablas Suplementarias 1, 2, 3 y Figura Suplementaria 1). Asimismo, el relevamiento tafonómico permitió identificar la presencia de huellas de procesamiento antrópico en seis especímenes (huellas de corte, machacado, impacto, desbastes y negativos de lascado) y marcas de carnívoros en dos de los especímenes (pits, punctures y scoring) (Figura 3 y Texto Suplementario 1).

Figura 2. Especímenes estudiados de Canis familiaris. A: La Lechuza (MRA-LZA-D7-130), hemimandíbula derecha: vista labial, oclusal y lingual. B: Sambaquí de Puerto Landa (MAMA-SPL-A), hemimandíbula izquierda: vista labial, oclusal y lingual. C: Cerros de los Pampas (MAMA-CP-244), hemimandíbula derecha: vista labial, oclusal y lingual. D: Arroyo Las Mulas 1 (MAS-LM1-SO1-D11), primer molar inferior derecho: vista labial, lingual y oclusal. E: La Palmera V (MAS-LPV-C1-5), primer molar superior izquierdo: vista labial, lingual, oclusal y mesial. F: Sambaquí de Puerto Landa (MAMA-SPL-B), fémur izquierdo: vista craneal y caudal. G: Sambaquí de Puerto Landa (MAMA-SPL-85), canino superior izquierdo: vista labial y lingual. H: Margen derecha del río Uruguay sin procedencia exacta (MAMA-s/p-A), canino inferior derecho: vista labial y lingual. (Color en la versión electrónica)
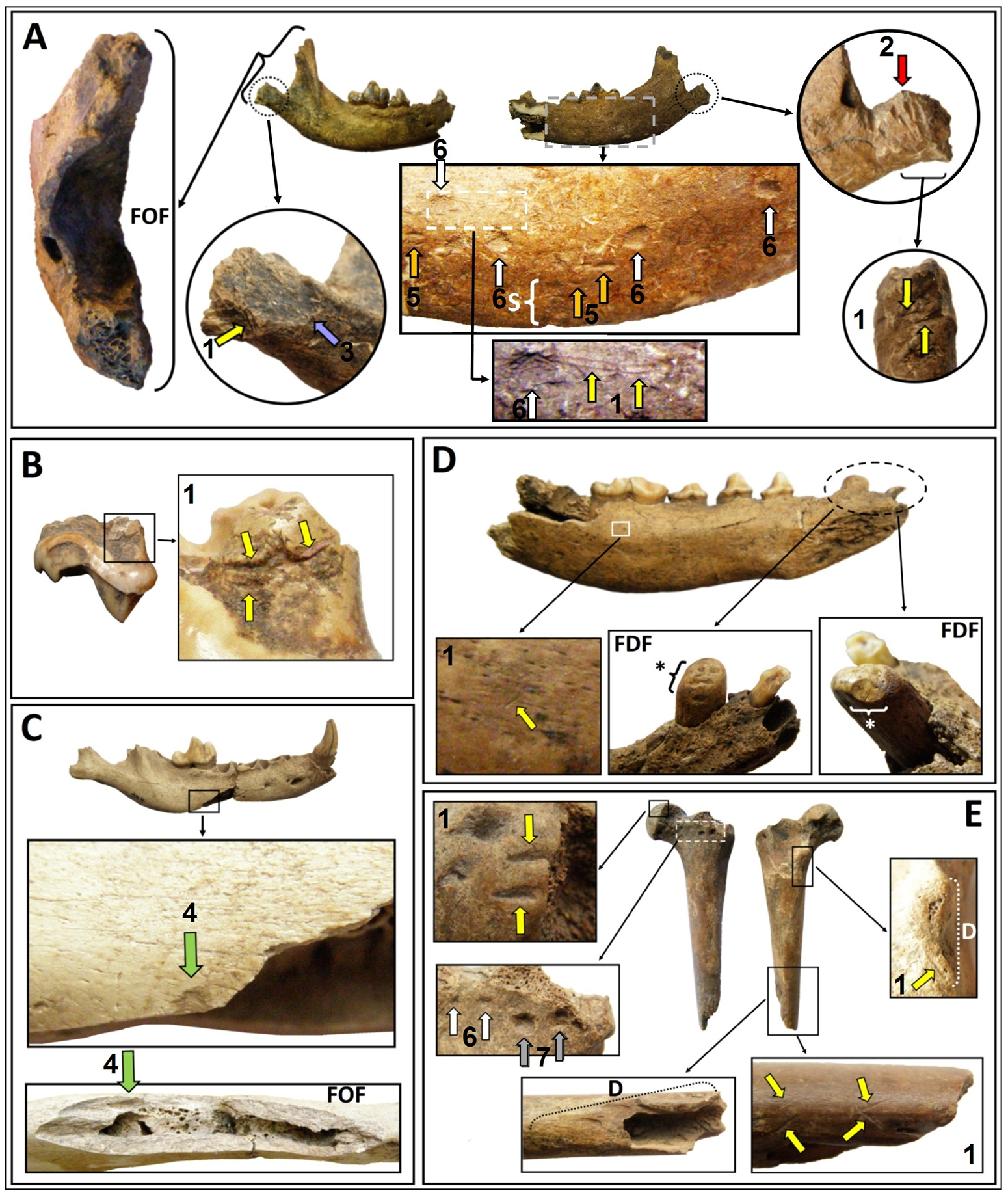
Figura 3. Especímenes de Canis familiaris con huellas antrópicas y marcas de carnívoros. A: La Lechuza (MRA-LZA-D7-130); B: La Palmera V (MAS-LPV-C1-5). C: Cerros de los Pampas (MAMA-CP-244). D: Sambaquí de Puerto Landa (MAMA-SPL-A). E: Sambaquí de Puerto Landa (MAMA-SPL-B). Número y color de las flechas. 1-Amarillo: huellas de corte; 2-Rojo: machacado; 3-Azul: negativo de lascado; 4-Verde: negativo de impacto; 5-Anaranjado: posibles crescent-shape pits; 6-Blanco: pits; 7-Gris: punctures. S: scoring. D: Desbaste. FOF: fractura ósea fresca. FDF: fractura dental fresca sobre el canino. *Fractura secundaria sobre la corona del canino. (Color en la versión electrónica)
Discusión y contextualización de los resultados
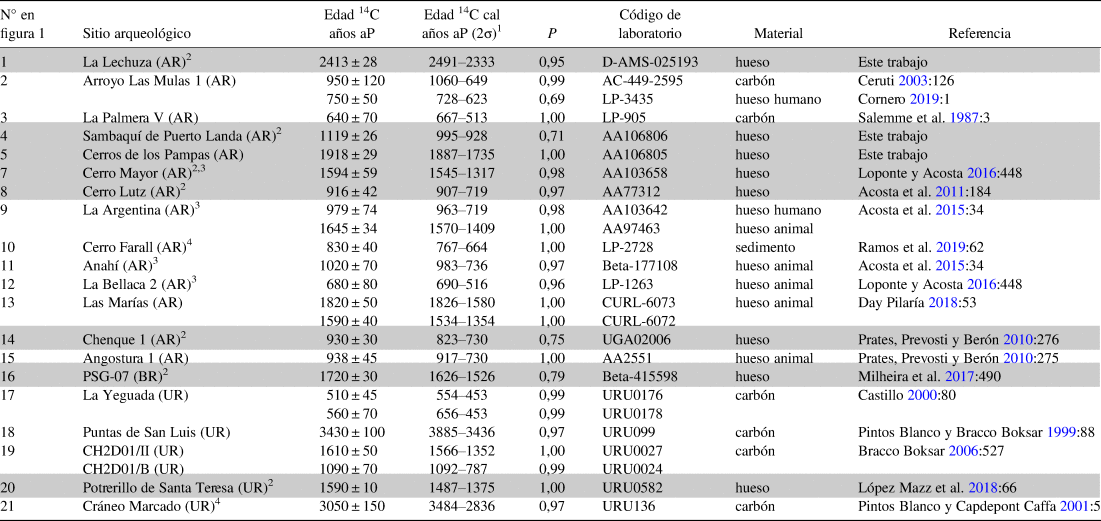
Los perros estudiados en este trabajo permiten ampliar la dispersión geográfica y temporal conocida hasta el momento para este taxón. Si bien ya había perros registrados para sitios arqueológicos de la región, estos se limitaban a la cuenca inferior del Plata (Day Pilaría Reference Day Pilaría2018; Loponte y Acosta Reference Loponte and Acosta2016). Con los especímenes de LZA, ALM1 y LPV se confirma la presencia de este animal en el curso medio del río Paraná. Asimismo, las tres nuevas fechas taxón aportadas en este artículo se suman a las cinco existentes para Argentina, Brasil y Uruguay (Tabla 2 y Texto Suplementario 2). Si bien son necesarias más fechas taxón para los perros que no fueron directamente fechados, el contexto arqueológico general de los sitios arqueológicos de procedencia y las dataciones anteriores a la conquista europea indicarían que corresponden a perros prehispánicos. En ALM1 y LPV los molares se ubican estratigráficamente por encima de las dataciones, a 15 y 10 cm respectivamente. En SPL la vinculación cronológica más clara está dada por la idéntica procedencia del fémur y la hemimandíbula datada (Tabla 1).
Tabla 2. Edades radiocarbónicas de sitios arqueológicos con Canis familiaris en Argentina, Brasil y Uruguay.
Nota: Las dataciones sombreadas indican las fechas taxón y las restantes corresponden a dataciones de los contextos arqueológicos.
1 Las dataciones fueron calibradas con Calib Rev 7.0.4 (Stuiver et al. Reference Stuiver, Reimer and Reimer2017) utilizando la curva de calibración para el Hemisferio Sur (SHCal13; Hogg et al. Reference Hogg, Hua, Blackwell, Niu, Buck, Guilderson, Heaton, Palmer, Reimer, Reimer, Turney and Zimmerman2013).
2 Sitios arqueológicos que presentan más fechados contextuales además de las fechas taxón.
3 Los datos sobre estos fechados presentan problemas, ya que en distintas publicaciones aparecen con diferentes valores radiocarbónicos, rangos, códigos de laboratorio, material datado o incluso referidos a distintos sitios arqueológicos (compárese e.g., Acosta et al. Reference Acosta, Buc, Ramírez, Prevosti and Loponte2015:34; Loponte y Acosta Reference Loponte and Acosta2016:434 y Loponte y Corriale Reference Loponte and Corriale2013:1385).
4 No se ha publicado evidencia clara sobre la determinación taxonómica del perro de este sitio arqueológico. AR: Argentina. BR: Brasil. UR: Uruguay.
De acuerdo con la muestra analizada (NISP = 8; MNI = 7), SPL tiene un MNI = 2 ya que, si bien los especímenes son uno inferior (hemimandíbula, MAMA-SPL-A) y otro superior (C1, MAMA-SPL-85), presentan edades notablemente diferentes; los dientes de la hemimandíbula muestran un intenso desgaste que no se registra en el canino aislado y, por lo tanto, no corresponderían al mismo individuo. Paralelamente, el fémur procede del mismo sector y nivel estratigráfico que la hemimandíbula, indicando posiblemente que correspondan al mismo individuo. Por el momento, SPL junto a CH2D01 (y posiblemente Cerro Mayor) son los únicos sitios arqueológicos de la región con más de un perro.
Edad y sexo
Las edades estimadas para los perros estudiados van desde juveniles, ligeramente mayores de cuatro a cinco meses, hasta adultos mayores, con más de ocho años (Tabla 1). Los individuos que llegaron a la etapa adulta o la superaron, indican su cuidado durante su etapa más vulnerable, que es su primer año. Específicamente, el estado de la dentición del individuo MAMA-SPL-A, sea un individuo adulto mayor (o joven/adulto con una dieta muy abrasiva), muestra el cuidado humano de ese animal, ya que de cualquier manera un cánido con esos dientes vería fuertemente reducidas sus posibilidades de supervivencia en estado silvestre. Un cuidado especial también es propuesto por Berón (Reference Berón, Gutiérrez, Nigris, Fernández, Giardina, Gil, Izeta, Neme and Yacobaccio2010:547) para el perro adulto mayor de Chenque 1, el cual además presenta una herida con recuperación ósea que no fue la causa de su muerte. En relación con el sexo, se estimaron dos machos correspondientes a LZA y CP (Tabla 1), siendo el primero subadulto y el otro juvenil; los que se suman a la hembra adulta de Cerro Lutz (Acosta et al. Reference Acosta, Loponte and Esponda2011), constituyendo los tres únicos casos de la región donde se ha estimado el sexo.
Reconstrucción morfológica
En cuanto a la estimación de la masa corporal (Tabla 1), los perros de LZA, LPV y SPL presentaban valores entre 13 y 17 kg y son semejantes a las máximas masas corporales registradas para Argentina, donde los promedios estimados abarcan un rango entre 6 y 16 kg (véase Belotti López de Medina Reference Belotti López de Medina2017); y también para Brasil y Uruguay, cuyo rango conjunto varía entre 11 y 17 kg (Texto Suplementario 2 y Tabla Suplementaria 4). Por su parte, los perros de ALM1 y CP son los individuos de mayor masa corporal publicados hasta el momento para Sudamérica. Si bien el ejemplar de ALM1 sobresale de la media, los restantes perros aquí estudiados, junto a los de Brasil y Uruguay, indican homogeneidad en cuanto al tamaño corporal mediano (masa corporal y alzada) de los individuos, posiblemente como resultado de la acción humana o una reproducción libre (panmixia; Bartosiewicz Reference Bartosiewicz2002).
Al igual que en los otros casos del Cono Sur (López Mazz et al. Reference López Mazz, Moreno, Bracco and González2018; Loponte y Acosta Reference Loponte and Acosta2016; Prates, Prevosti y Berón Reference Berón, Gutiérrez, Nigris, Fernández, Giardina, Gil, Izeta, Neme and Yacobaccio2010), ninguno de los morfotipos de las hemimandíbulas y de los dientes aislados estudiados corresponde a las variedades americanas de perros sin pelo características de México (xoloitzcuintle), de Perú y Bolivia (perro sin pelo peruano, viringo, k'ala, ccala, ccalato, pelón) o de Argentina (pila). La ausencia de pelo es el resultado de una mutación (Displasia ectodérmica canina), que además trae aparejadas anormalidades dentarias como hipodoncia y malformaciones en las cúspides (Kupczik et al. Reference Kupczik, Cagan, Brauer and Fischer2017; Lewis et al. Reference Lewis, Reiter, Mauldin and Casal2010; Vásquez et al. Reference Vásquez, Rosales, Gálvez and Dorado2016).
Como dato comparativo, se puede agregar que la hemimandíbula izquierda de Sambaquí de Puerto Landa carece del alveolo correspondiente al p1, característica observada en numerosos perros americanos prehispánicos (e.g., Allen Reference Allen1920; Crockford Reference Crockford1997; Lupo y Janetski Reference Lupo and Janetski1994). Este mismo rasgo está presente en dos perros de Uruguay. En CH2D01/II, corresponde al alveolo del p1 derecho, y en CH2D01/B, del p1 izquierdo; además, en el último caso también falta el alveolo del M2 derecho (López Mazz et al. Reference López Mazz, Moreno, Bracco and González2018:68, 72). Por su parte, al perro de Cerro Lutz le falta el p1 izquierdo y hay un incisivo extra entre el i1 e i2 de ambas hemimandíbulas (Acosta et al. Reference Acosta, Loponte and Esponda2011). Este último caso de incisivos supernumerarios también ha sido reportado para tres ejemplares de la Quebrada del Toro (provincia de Salta, Argentina; Zetti Reference Zetti1973). Finalmente, debido a la ausencia de cráneos y poscráneos más completos, no se puede avanzar mucho más en la reconstrucción morfológica de los perros.
Tafonomía
Las modificaciones naturales relevadas en los especímenes arrojaron información semejante a la obtenida previamente para los restos arqueofaunísticos de la región (e.g., Bonomo et al. Reference Bonomo, Castro and Silva2014; Castro Reference Castro2017; Sartori y Galligani Reference Sartori and Galligani2014). En nuestro caso, una baja frecuencia de marcas de raíces y nula de roedores, así como el nulo o bajo estadio de meteorización de las muestras, indica que estos factores no han incidido significativamente en la preservación diferencial de los especímenes. La elevada representatividad del esqueleto craneal además probablemente esté influida por sesgos de muestreo hacia elementos taxonómicamente más diagnósticos, como los dientes, y por factores culturales como, por ejemplo, el tratamiento diferencial o el procesamiento dado a estos carnívoros. Por su parte, los depósitos de MnO2 son propios de ambientes con un alto grado de humedad y/o con la descomposición de materia orgánica, como el que caracteriza el entorno de los sitios arqueológicos de procedencia de las muestras.
Es importante destacar la presencia de marcas de carnívoros, que han masticado, fracturado y provocado pérdida de material óseo en algunos de los especímenes. La coexistencia de marcas de carnívoros y huellas antrópicas en los mismos huesos (e.g., LZA y SPL) podría indicar, primero un procesamiento y consumo humano de los perros, y en segunda instancia, luego del descarte, un consumo por parte de carnívoros (incluyendo posiblemente otros perros). En consonancia con lo anterior, se registraron numerosas huellas de procesamiento antrópico en seis de los ocho especímenes presentados; estas incluyen: huellas de corte, de machacado, de impacto, desbastes, un negativo de lascado y fracturas frescas asociadas a estos atributos. Las actividades inferidas a partir de cada uno de los elementos anatómicos representados son diversas.
La hemimandíbula de LZA evidencia tareas de desarticulación anatómica con el neurocráneo, descarne y extracción de la lengua, y seccionamiento de los músculos masetérico y temporal, y posiblemente cuereo. La hemimandíbula de SPL evidencia tareas de descarne y extracción de la lengua, lo que posiblemente también implicó una desarticulación mandíbula-neurocráneo previa, como en el perro de LZA. Este es el único espécimen que registra patologías como ausencia de un alveolo y dientes fracturados en vida. En cuanto a estas fracturas, en el canino y el i3, posiblemente sean producto de la dieta del animal, lo que va en consonancia con el desgaste muy avanzado de todos los dientes. Algunas causas de fracturas naturales incluyen, por ejemplo, dietas restringidas al carroñeo de carcazas y/o comidas duras, peleas entre pares y lesiones ocasionadas durante la caza por grandes presas (Losey et al. Reference Losey, Jessup, Nomokonova and Sablin2014). Cabe mencionar que las fracturas antrópicas y la remoción de dientes han sido registradas en varios sitios arqueológicos de México y en Groenlandia en tiempos históricos (Losey et al. Reference Losey, Jessup, Nomokonova and Sablin2014; Wing Reference Wing, Genoways and Dawson1984). La hemimandíbula de CP evidencia agencia humana a partir de los atributos de su fractura longitudinal y el negativo de impacto asociado. Recientemente se ha publicado una hemimandíbula de un lobo del Pleistoceno procedente del sitio arqueológico Předmostí (República Checa) del Paleolítico superior (Germonpré et al. Reference Germonpré, Lázničková-Galetová, Jimenez, Losey, Sablin, Bocherens and Martine van den2017). Esta pieza presenta una huella de impacto antrópico directamente asociada a una fractura longitudinal-oblicua semejante a la presentada en este trabajo (Figura 3C).
Por su parte, el molar de LPV evidencia tareas de extracción dental y/o elaboración de algún tipo de artefacto. El canino sin procedencia muestra una intensa manipulación antrópica que podría explicarse como resultado de su limpieza, tal vez para la elaboración de un pendiente, como se ha observado con carnívoros de la región Pampeana y en el Noreste argentino, incluyendo C. familiaris (Acosta et al. Reference Acosta, Buc, Ramírez, Prevosti and Loponte2015; Bonomo Reference Bonomo2006; Castro Reference Castro2017). Por último, el fémur de SPL evidencia tareas de desmembramiento, descarne y extracción de médula ósea, y posiblemente también la elaboración de artefactos, como muestra el registro arqueológico y etnográfico regional, donde se han identificado instrumentos sobre huesos largos de cánidos (Castro Reference Castro2017:315, 347, 348).
Roles y usos
Las evidencias de acción antrópica en diferentes sitios arqueológicos están indicando a nivel regional el procesamiento integral (cráneo y poscráneo) de perros en el lapso temporal comprendido entre aproximadamente 2491 y 513 años cal aP (Tabla 2). Las fracturas en estado fresco, los negativos de impacto y las huellas de machacado y de corte asociadas a la desarticulación, separación de músculos como la lengua y el descarne, señalan que los perros fueron manipulados para el consumo humano. Si bien no se han registrado evidencias de termoalteración, los contextos de descarte junto a fragmentos de vasijas utilizadas para la cocción de alimentos y los huesos de otros animales consumidos apoyan esta idea. Esta manipulación se habría realizado con el objetivo de obtener su carne y médula ósea para la ingesta ocasional, sus cueros para prendas o artefactos, sus huesos para instrumentos y sus dientes para adornos. Además de sus posibles roles como compañía y ayudantes en la caza, y quizá la pesca, en las cuales su uso pudo haber ampliado el rango de presas capturadas y aumentar la eficacia de los eventos de cacería (Castro Reference Castro2017:447, 450), los perros también contribuirían a la limpieza de los desperdicios en los asentamientos, la disuasión de predadores del entorno y de otros animales que merodeaban en busca de desechos. Asimismo, aumentan las posibilidades de alerta y defensa ante posibles enemigos.
El consumo de perros es actualmente una práctica recurrente en ciertas partes de África, Asia o el Pacífico donde son apreciados por su carne y/o sus propiedades rituales y simbólicas (Le Breton Reference Le Breton2007). Esto también se ha registrado en distintos lugares de América desde el inicio mismo de la conquista europea (e.g., Allen Reference Allen1920; Herrera y Tordesillas Reference Herrera y Tordesillas1601–1615; Morey Reference Morey2010; Vásquez et al. Reference Vásquez, Rosales, Gálvez and Dorado2016; Weiss Reference Weiss1970). Es interesante además la mención entre los indígenas americanos de la cría, sacrificio y consumo de “perros mudos” (e.g., De la Garza Reference De la Garza1997:114–118; Gallardo Reference Gallardo1964–1965:36–38, 51–53; Hernández de Alba Reference Hernández de Alba and Steward1948:326). Para la región del Noreste argentino, el cronista Fernández de Oviedo y Valdés Reference Fernández de Oviedo y Valdés1852 [1546–1547]:192) señala que los agricultores denominados barrigudos “tienen unos perrillos que crian en sus casas, mudos, que no ladran, y los tienen por buen manjar, y los comen quando quieren”. Si bien está en discusión el origen natural o antrópico de la mudez de los perros americanos (véase Stahl Reference Stahl2013), Blas Jaime (el último chaná parlante) refiere que los indígenas chaná tenían perros (agó en lengua chaná) que eran mudos porque les cortaban las cuerdas vocales (cordectomía) para silenciarlos, y que además tenían otros tipos de perros que eran castrados y engordados para su consumo. Es importante destacar que, en ninguno de los perros del registro arqueológico de la región publicados previamente (Tabla 2 y Texto Suplementario 2) se habían registrado evidencias de procesamiento humano con esta magnitud. Solamente el axis de La Bellaca 2 atribuido a C. familiaris por Loponte y Acosta (Reference Loponte and Acosta2016) presentaría huellas de corte indicando su separación del atlas, y por lo tanto la cabeza del cuerpo.
Además de los usos mencionados, el perro también tuvo un rol simbólico, que se encuentra plasmado en la alfarería prehispánica de la región. Entre los apéndices zoomorfos de distintos sitios arqueológicos (Figura 4), se identificaron rasgos anatómicos diagnósticos de perros de acuerdo con la morfología de la cabeza alargada con cuello cilíndrico, nariz con cavidades nasales abiertas, ojos laterales, bocas con dientes y orejas terminadas en punta. Estas representaciones modeladas en arcilla son características de Goya-Malabrigo (Politis y Bonomo Reference Politis and Bonomo2018), donde las especies animales incluidas en los apéndices zoomorfos tuvieron una valoración simbólica destacada y, en general, no fueron una fuente regular de alimento, ya que su consumo estuvo socialmente restringido o sujeto a tabúes alimentarios permanentes o circunstanciales, completos o parciales (Bonomo et al. Reference Bonomo, Politis, Bastourre, Moreira, Bonomo and Archila2020). En estos casos particulares, estos apéndices están señalando no sólo la presencia de perros en la vida cotidiana de los indígenas, sino también creencias compartidas en relación con estos cánidos.

Figura 4. Perros modelados en cerámica del Noreste argentino (entidad arqueológica Goya-Malabrigo). A: Islas del Delta superior del Paraná (Victoria-Rosario. Colección Museo de Victoria). B: Islas de Coronda (Coronda, Santa Fe. Colección Ñañez). C: Sitio arqueológico Laguna del Plata II (Vera, Santa Fe. Colección Museo Serrano). D: Sitio arqueológico Arroyo Las Mulas 1 (La Paz, Entre Ríos. Colección Museo Serrano) (nótese la presencia de un posible collar). E: Río Paraná (San Justo, Santa Fe. Colección Museo de Gobernador Crespo). F: Sitio arqueológico Cerro Grande/Chico (Victoria, Entre Ríos. Colección Oberti). (Color en la versión electrónica)
Por último, la antigüedad del registro de perros en el NEA lleva a repensar las vías de dispersión de este taxón hacia las latitudes más meridionales del continente americano. En principio, la ausencia de registros arqueológicos en el Chaco y la Amazonía, sumada a su escasa documentación al momento del arribo de los europeos, no permiten proponer su llegada a través de estas regiones. La llegada de los perros al río Paraná pudo darse por contacto con la región andina meridional o el centro-norte de Argentina, como es el caso, por ejemplo, de las láminas de metal, las llamas, y los cóndores o las estatuillas antropomorfas representadas en cerámica (e.g., Bonomo et al. Reference Bonomo, Cabanillas and Montero2017; Castro Reference Castro2017; Politis y Pedrotta Reference Politis and Pedrotta2006). En el área andina además hay claras evidencias del consumo de perros en tiempos prehispánicos (e.g., Vásquez et al. Reference Vásquez, Rosales, Gálvez and Dorado2016). Posteriormente, la especie se habría dispersado en dos direcciones, alcanzando, por un lado, el este y noreste de Uruguay y el sudeste de Brasil, y, por otro lado, la region Pampeana y la Patagonia. Para conocer con mayor precisión el proceso de dispersión de C. familiaris, esta hipótesis deberá ser evaluada a partir de estudios de ADN y nuevas fechas taxón procedentes de distintas regiones.
Consideraciones finales
En este artículo se contribuye con nueva información científica sobre los perros indígenas prehispánicos de América del Sur. Se sumaron cinco sitios arqueológicos con este tipo de registro faunístico (LZA, ALM1, LPV, SPL y CP) a los previamente conocidos, ampliando así la escala de distribución geográfica de este taxón. Asimismo, las nuevas dataciones extienden la antigüedad de C. familiaris en la región bajo estudio a la primera mitad del Holoceno tardío. Específicamente, los fechados obtenidos en LZA y CP son los más tempranos que existen hasta el momento para el Cono Sur y muestran que la incorporación del perro en las sociedades indígenas del sudeste sudamericano fue anterior a lo que se conocía. Asimismo, la edad radiocarbónica más temprana de 2491-2333 años cal aP obtenida para el perro de LZA, constituye la segunda datación sobre el taxón más antigua de Sudamérica.
Los ejemplares de C. familiaris estudiados presentan edades y tamaños ligeramente mayores a los registrados previamente en Argentina, Brasil y Uruguay. Aun así, los perros indígenas poseían tamaños medianos a pequeños, a diferencia de los perros más grandes traídos desde Europa a partir del siglo dieciséis. Asimismo, los nuevos perros identificados corresponden al menos a un morfotipo diferente al perro sin pelo americano. Por su parte, las claras evidencias de procesamiento observadas en sus huesos y dientes, sumadas a su contexto de descarte, permiten concluir que algunos de ellos fueron matados, desarticulados y descarnados para el consumo humano.
La mayoría de los nuevos especímenes fueron recuperados en asentamientos Goya-Malabrigo, donde los perros fueron un componente más de una economía indígena diversificada, basada en la pesca, la caza y la recolección, complementada con una horticultura a pequeña escala. Dada la magnitud de las evidencias de procesamiento observadas en las muestras de distintos sitios arqueológicos, el consumo de perros parece haber sido una práctica extendida, la cual posiblemente ocurrió de manera ocasional y bajo circunstancias con un fuerte contenido simbólico ya que, además, son una de las especies que fueron representadas en la alfarería. El caso de los perros parece reflejar la situación dual de algunos animales con un estatus singular, que fueron modelados en arcilla y que también se consumieron en determinadas situaciones.
El rol de los perros en las sociedades indígenas del Cono Sur, inferido a partir de la arqueología y las crónicas tempranas, fue el de compañía en la vida y en la muerte, fuente de alimento y materias primas (pieles, huesos y dientes), referentes simbólicos representados en la cerámica y ayudantes en actividades cinegéticas. Este estrecho vínculo entre perros y humanos ha quedado plasmado en diversos contextos arqueológicos, domésticos y funerarios, que a nivel regional muestran un elevado número de ejemplares que supera los veinte individuos, evidenciando su importancia en las dimensiones socioeconómicas y simbólicas de los indígenas sudamericanos.
Canis familiaris tiene una larga historia de aproximadamente 2.500 años en el sur de Sudamérica. Por lo tanto, puede afirmarse que los seres humanos que ocuparon la región estuvieron acompañados por perros, los cuales desempeñaron diversos roles y participaron de la trayectoria histórica de las sociedades indígenas locales, desde el inicio del Holoceno tardío hasta momentos posteriores a la conquista europea del siglo dieciséis.
Agradecimientos
A Francisco Prevosti por sus valiosos aportes. A Germán Moreira, César García Esponda, Eduardo Tonni, Raúl Almeida, Carlos Ceruti, Blas Jaime, Claudio Ñañez, Diego Bracco, Diego Gobbo, Oscar Heit, Kristian González, Enoch Walls, John Fackenthal, Fabián Letieri, Gabriel Cocco, Moira Sotelo, Federica Moreno, José López Mazz, Natalia Derudi, Laura Miotti, Peter Stahl e Iban Sánchez-Pinto. Especialmente a Luciano Rey, Subdirector de Patrimonio Cultural del Ministerio de Innovación y Cultura de Santa Fe. Parte de este trabajo fue financiado por los proyectos ANPCyT PICT 1064 y UNLP 11/N885. Una primera versión de este estudio fue presentada en el congreso anual 83 de la SAA (Washington, DC) en 2018.
Declaración de disponibilidad de datos
Los especímenes reportados se encuentran en los museos Arqueológico Manuel Almeida, de Ciencias Naturales y Antropológicas Profesor Antonio Serrano y Universitario Florentino y Carlos Ameghino.
Materiales suplementarios
Para acceder a los materiales que acompañan este artículo, visitar www.journals.cambridge.org/[LAQ].
Texto Suplementario 1. Descripción taxonómica, anatómica y tafonómica de los especímenes de Canis familiaris y su comparación con otros cánidos.
Texto Suplementario 2. Nuevos datos para el registro arqueológico regional. Entierros humanos y caninos en el sudeste del Cono Sur.
Tabla Suplementaria 1. Valores morfométricos de las hemimandíbulas.
Tabla Suplementaria 2. Valores morfométricos de los dientes en las hemimandíbulas y de los dientes aislados.
Tabla Suplementaria 3. Medidas de molares superiores (m1) de Canis familiaris registrados en Argentina y Brasil.
Tabla Suplementaria 4. Atributos de Canis familiaris de Argentina, Brasil y Uruguay.
Figura Suplementaria 1. Relación entre los valores obtenidos para perros procedentes de sitios arqueológicos y cánidos silvestres de la región.








