



La diversidad que adquieren las conductas humanas con relación a las pautas de movilidad y aprovisionamiento de recursos dentro de un paisaje costero ha sido un tema ampliamente debatido en arqueología (e.g., Bailey y Davidson Reference Bailey and Davidson1983; Binford Reference Binford2001; Erlandson Reference Erlandson2001). Gran parte de estos debates se han articulado a partir de diferentes modelos generales de movilidad (Bailey y Davidson Reference Bailey and Davidson1983; Binford Reference Binford1980; Kelly Reference Kelly1995) cuyas predicciones se basan fundamentalmente sobre la estructura del ambiente y la distribución espacial y temporal (estacional) de los recursos más significativos para la subsistencia humana. En este sentido, el análisis de los radios o rangos de aprovisionamiento de recursos se plantea como un tema central en la comprensión de las pautas de movilidad (Bailey y Davidson Reference Bailey and Davidson1983; Binford Reference Binford1980; Fitzhugh Reference Fitzhugh, Fitzhugh and Habu2002; Kelly Reference Kelly1995:135). Estos son analizados mediante diversas premisas, pero fundamentalmente considerando los costos implicados en recorrer un espacio determinado (Bailey y Davidson Reference Bailey and Davidson1983) y en el transporte que involucra, entre otros factores, el volumen y peso de la carga, la distancia y la tecnología empleada (e.g., Lupo Reference Lupo2006; Metcalfe y Barlow Reference Metcalfe and Barlow1992). Debido al carácter fragmentario del registro arqueológico, muchas de las predicciones generadas por estos modelos etnoarqueológicos son difíciles de evaluar (Lupo Reference Lupo2006). No obstante, los estudios zooarqueológicos se han afianzado en la discusión de estos problemas a través de modelos cada vez más exhaustivos (e.g., Alunni Reference Alunni2018; Burger et al. Reference Burger, Hamilton and Walker2005; Cannon Reference Cannon2003; Lupo Reference Lupo2006).
Los conjuntos arqueológicos costeros de Tierra del Fuego poseen un importante potencial para contribuir al conocimiento general sobre las pautas de movilidad y uso de los ambientes costeros por cazadores-recolectores-pescadores. Esto se debe a que los casos etnográficos observados en esa región no se corresponden con las expectativas derivadas de los parámetros generales con los que usualmente se identifican sociedades costeras de climas fríos y templados (Binford Reference Binford2001; Kelly Reference Kelly1995). Estas excepciones se vinculan mayormente con reducidas prácticas de almacenamiento (Binford Reference Binford2001:257), lo cual implica movimientos logísticos amplios (Kelly Reference Kelly1995) y una elevada movilidad residencial de los grupos fueguinos (Binford Reference Binford2001:422). Además, se ha señalado que las pautas de procesamiento y descarte identificadas en el registro arqueológico costero de Tierra del Fuego (Borrero Reference Borrero1986; Muñoz Reference Muñoz2005) no dan cuenta de las expectativas derivadas de los modelos etnoarqueológicos propuestos por Binford (Reference Binford1980). Los antecedentes disponibles para el norte de Tierra del Fuego indican que los cazadores-recolectores pedestres explotaron guanacos a partir de rangos logísticos amplios, combinados con una elevada movilidad residencial, la cual estuvo orientada a promediar la disponibilidad del recurso en el espacio (Borrero Reference Borrero1986; Muñoz Reference Muñoz2005). No obstante, los patrones implicados para la explotación de recursos pudieron resultar diferentes al sur de esa isla.
La costa meridional de Tierra del Fuego ofrece un panorama interesante para abordar esta problemática debido a la existencia de factores culturales y ambientales que influyeron sobre las posibilidades de movilidad y transporte de los grupos cazadores-recolectores-pescadores (Orquera y Piana Reference Orquera and Piana1999). La explotación de animales en esa región cubrió un espectro amplio, registrándose desde conjuntos totalmente integrados por recursos marinos hasta contextos con una casi total dominancia de animales terrestres, en particular el guanaco (Lama guanicoe; Orquera y Piana Reference Orquera and Piana1999; Zangrando Reference Zangrando2009). A partir de este registro podrían plantearse diferentes situaciones organizativas o logísticas en el aprovisionamiento de las presas. El guanaco fue el único animal terrestre cuyo aprovechamiento planteó una planificación en términos logísticos (sensu Binford Reference Binford1980) y tuvo un rol significativo como fuente de alimento y de materias primas para la producción de tecnología. De esta manera, el modo en que fue aprovechado este recurso dentro de un contexto de ocupación costera puede aportar información relevante para comprender los patrones conductuales que configuraron el uso del espacio, sus recursos y la movilidad entre la costa y el interior.
Recientemente nos propusimos examinar las estrategias del uso del espacio y aprovisionamiento de recursos por sociedades cazadoras-recolectoras-pescadoras en la Bahía Moat (Figura 1; Zangrando Reference Zangrando2010). En el marco de este proyecto, uno de nuestros propósitos fue el de evaluar con qué amplitud los espacios hacia el interior fueron integrados con los ámbitos litorales. A diferencia de los sectores más occidentales del canal Beagle, en la Bahía Moat el espaldar montañoso es menos elevado y los valles de ríos son más amplios y transversales a la línea de costa, lo cual ofrece mejores condiciones de accesibilidad hacia el interior. Además, la cuenca del río homónimo pudo haber facilitado la circulación entre espacios terrestres y la costa sur de Tierra del Fuego. La Bahía Moat y el área que se proyecta valle adentro es un ámbito para el cual no se habían efectuado investigaciones arqueológicas previas a nuestros estudios y los únicos antecedentes disponibles provienen de la etnografía. Esta fuente de información señala que a fines del siglo diecinueve y principios del siglo veinte, el uso de este espacio pudo haberse desarrollado a través de diferentes estrategias de subsistencia y formas de movilidad que habrían estado articuladas a partir de capacidades tecnológicas distintas para la captura de los recursos (Bridges Reference Bridges1878).

Figura 1. Ubicación del área de estudio y los sitios arqueológicos analizados.
Estos antecedentes brindan una perspectiva interesante para repensar y discutir algunos aspectos de la movilidad humana que han sido escasamente tratados en la arqueología de la franja meridional de Tierra del Fuego (Zangrando Reference Zangrando2009; Zangrando et al. Reference Zangrando, Borrazzo, Tivoli, Alunni and Martinoli2014). En el marco de estas discusiones los objetivos de este trabajo son: (a) analizar las pautas de explotación de guanacos por parte de los cazadores-recolectores-pescadores que ocuparon la Bahía Moat; (b) evaluar la importancia de estos recursos como fuente de grasas y proteínas; y (c) discutir las implicaciones para la movilidad de dichas poblaciones humanas.
Expectativas relativas a la explotación de guanacos e implicaciones para la movilidad humana entre la costa y el interior
La costa sur de Tierra del Fuego presenta un clima templado-frío oceánico, con una temperatura media anual de 5,3°C (Iturraspe y Schroeder Reference Iturraspe, Schroeder, Orquera and Piana1999). Es un ámbito ventoso y húmedo, con valores de precipitaciones que oscilan entre 500 y 1.000 mm anuales. La Cordillera Fueguina se extiende en estrecha cercanía a lo largo de la costa; sus cumbres agudas alcanzan una altitud de 2.500 m snm y se van haciendo más bajas y menos escarpadas hacia el este. El constante exceso hídrico, la ausencia de glaciares y la baja acumulación nívea favorecen la formación de grandes extensiones de turberas y numerosas lagunas pequeñas (Roig y Collado Reference Roig, Collado, Blanco and de la Balze2004). La zona está caracterizada por sectores de bosques de Nothofagus spp. más o menos densos entremezclados con áreas de turberas y pastizales. En el sector litoral habitan una diversidad importante de animales marinos, entre los que se incluyen pinnípedos, cetáceos, aves, peces y moluscos (Bastida y Rodríguez Reference Bastida and Rodríguez2003; Forcelli Reference Forcelli2000; Lloris y Rucabado Reference Lloris and Rucabado1991; Schiavini y Yorio Reference Schiavini and Yorio1995).
La Bahía Moat se ubica hacia la margen sur-oriental de Tierra del Fuego, entre el Río Cambaceres y la Bahía Sloggett. Este sector se localiza en las últimas estribaciones del canal Beagle (Figura 1), quedando entonces más expuesto a los vientos y tormentas del sureste. Las precipitaciones (media anual de ca. 700 mm) son algo mayores que las que ocurren en las cercanías de la Bahía de Ushuaia y lindantes con la cordillera andina (media anual de ca. 500 mm). El área que se proyecta hacia el interior exhibe valles amplios y transversales a la línea de costa, con sierras que alcanzan alturas entre 100 y 900 m snm. Esta zona también se caracteriza por presentar un campo de drumlins de 80 km2 de extensión (Borromei et al. Reference Borromei, Ponce, Coronato, Candel, Olivera and Okuda2014). Existen varios cursos fluviales que desaguan al mar, pero el Río Moat (Figura 1) es el afluente más extenso que recorre cerca de 50 km del valle en sentido sureste-noroeste, pudiendo haber conformado una vía de comunicación entre el interior y la costa tanto para los animales como para los grupos humanos (Zangrando Reference Zangrando2010).
Los guanacos en Tierra del Fuego se distribuyen en toda la provincia pero con mayor abundancia en el ecotono bosque-estepa (1,73 individuos/km2; Montes et al. Reference Montes, Lamo and Zavatti2000:Tabla 2), en la franja central de la isla. Aquí las condiciones topográficas y la calidad del forraje favorecen la selectividad de estos espacios para el pastoreo e incluso permiten el afianzamiento territorial de las tropillas familiares. En el entorno boscoso y cordillerano de nuestra área de estudio (localizada aproximadamente 50 km hacia el sur del ecotono), los guanacos son hallados en menores proporciones (0,68 individuos/km2; Montes et al. Reference Montes, Lamo and Zavatti2000: Tabla 2) y de forma más discontinua. El patrón de dispersión muestra que en esencia existe una población de guanacos que circula entre 40 y 80 km desde la costa y hasta aproximadamente 400 m de altura, mientras otros grupos están más vinculados al litoral y se desplazan hasta 20 km desde la playa y hasta unos 200 m de altura (Schiavini et al. Reference Schiavini, Escobar and Deferrari2010). No obstante, estos radios de acción fluctúan estacionalmente. Durante el otoño e invierno, la carga nívea y la severidad de las condiciones habitables en las zonas altas desencadenan desplazamientos migratorios hacia sectores de menor altitud. En consecuencia, durante dicho momento del año, la población de guanacos relacionada con la costa circula en un espacio de hasta 10 km desde el litoral (Schiavini et al. Reference Schiavini, Escobar and Deferrari2010). Por el contrario, la frecuencia de camélidos en la costa disminuye hacia el verano, dado que estos aprovechan la mejor calidad del forraje que ofrece el rebrote vegetal en el interior y en las áreas de mayor elevación (Raedeke Reference Raedeke1976). Entonces, en la época estival se nuclean principalmente en un radio de acción entre 10 y 20 km con respecto a la playa (Schiavini et al. Reference Schiavini, Escobar and Deferrari2010).
El patrón de dispersión arriba descripto tiene implicancias directas para los rangos de acción vinculados con las pautas de subsistencia de los cazadores-recolectores. De acuerdo con la estructura espacial del guanaco y sus movimientos estacionales, es probable que los grupos humanos hayan desarrollado estrategias alternativas según el momento del año. Durante el invierno, pudieron haber practicado desplazamientos cercanos a los entornos costeros que no superaran los 10 km de distancia hacia el interior. En cambio, en verano la amplitud del espacio recorrido pudo haber sido mayor, lo cual habría implicado trayectos más amplios hacia el valle y una planificación diferente para el aprovisionamiento y transporte de alimento. Al respecto, Bridges (Reference Bridges1878:11-12) describió, para la margen sur de Tierra del Fuego, que los yaganes, pueblo originario que ocupaba el canal Beagle y sus inmediaciones al momento de la llegada de los europeos, a menudo incursionaban hacia el interior de las montañas, lejos de la costa, para obtener guanacos e instalaban lugares de cacería desde los cuales salían a recorrer grandes extensiones y se dispersaban al hacerlo. También relata que catorce indígenas partieron durante cinco días llegando a recorrer 25 km de distancia desde el litoral, capturando cinco guanacos (Bridges Reference Bridges1878:12).
Los modelos de optimización constituyen una valiosa herramienta analítica para explorar las estrategias de movilidad, transporte y procesamiento bajo esas posibles situaciones y para inferir estos comportamientos a partir del análisis de las arqueofaunas (Burger et al. Reference Burger, Hamilton and Walker2005; Cannon Reference Cannon2003; Lupo Reference Lupo2006; Madrigal y Holt Reference Madrigal and Holt2002; Metcalfe y Barlow Reference Metcalfe and Barlow1992). En trabajos previos hemos desarrollado una serie de expectativas arqueológicas para analizar la explotación de guanacos en la Bahía Moat sobre la base de estos modelos (Alunni Reference Alunni2018; Alunni y Zangrando Reference Alunni and Zangrando2012). Estas pueden ser abordadas considerando los costos de búsqueda, el rendimiento por unidad de peso (kcal/g) y el tiempo que lleva procesarlos para adquirir los distintos productos que sus carcasas ofrecen (kcal/h; e.g., Burger et al. Reference Burger, Hamilton and Walker2005; Cannon Reference Cannon2003; Lupo Reference Lupo2006; Madrigal y Holt Reference Madrigal and Holt2002; Metcalfe y Barlow Reference Metcalfe and Barlow1992). Esto implica tener en cuenta, por un lado, anatomía económica y el valor nutritivo que contienen las carcasas de los guanacos en función de los costos involucrados en el transporte y procesamiento y, por otro lado, la abundancia y distribución de guanacos en las condiciones arriba señaladas para la Bahía Moat. Las premisas tras estas variables suponen que los cazadores tienden a desplazarse para buscar el alimento desde sus bases residenciales (home range) y acarrear las presas hacia el lugar de consumo final (Cannon Reference Cannon2003; Metcalfe y Barlow Reference Metcalfe and Barlow1992). Un aspecto crítico a tener en cuenta es la distancia (tiempo de viaje o de búsqueda) a los recursos principales. Se presume que, a medida que aumenta la distancia entre la localidad de matanza y el lugar central, el cazador dedicará más tiempo en procesar una carcasa en el lugar de adquisición y más radical será el descarte de elementos de menor utilidad (Metcalfe y Barlow Reference Metcalfe and Barlow1992). De este modo, los individuos logran reducir el peso de la carga y el paquete transportable estará compuesto sólo por aquellas porciones que ofrezcan mayores calorías por unidad de peso (kcal/g).
En el marco de estas discusiones, también hemos propuesto que resulta necesario considerar no sólo el volumen de calorías obtenidas, sino también los productos específicos que los individuos buscan hallar en función de las necesidades dietarias y los requerimientos metabólicos (Alunni Reference Alunni2018). El consumo de grasas es de crucial importancia para las sociedades móviles que habitan climas fríos porque estas constituyen importantes reservas de energía (calorías), pero también por ser fuentes de vitaminas, minerales y ácidos grasos esenciales que no son elaborados por el cuerpo y deben obtenerse de los alimentos. Sobre este aspecto, si bien un guanaco adulto (100-140 kg; Raedeke Reference Raedeke1976:104) tiene un rendimiento calórico elevado de aproximadamente 78.315 kcal/individuo (Schiavini Reference Schiavini1990), posee carne magra, ya que la cantidad de grasa muscular que proporciona es menor al 1% (García Reference García1976). Las fuentes de lípidos más ricas están contenidas en el cráneo y en los canales medulares, aunque también posee dentro de su estructura ósea grasa trabecular alojada en el tejido esponjoso de los huesos (grasa ósea). En general, la médula se adquiere fácilmente a través de uno o varios golpes sobre la diáfisis del hueso; en cambio, la extracción de grasa ósea constituye una técnica de mayor costo, dado que sólo puede ser obtenida por medio del hervido. Esto implica no sólo mayor tiempo de procesamiento, sino también el uso de combustible y otros materiales (como la confección de contenedores) que complejizan la técnica de extracción (Janzen et al. Reference Janzen, Reid, Vasquez and Gifford-Gonzalez2014).
Conjuntos zooarqueológicos
Las prospecciones efectuadas a lo largo del litoral de la Bahía Moat y espacios aledaños (entre Baliza Davinson y Punta Final) indicaron la existencia de 56 sitios distribuidos en nueve localidades arqueológicas. La mayor parte de ellos corresponden a concheros en forma de montículos (Zangrando et al. Reference Zangrando, Borrazzo, Tivoli, Alunni and Martinoli2014). Hasta el momento, se han efectuado excavaciones y estudios sistemáticos en la localidad de Heshkaia. En este trabajo se analizan los conjuntos correspondientes a los sitios 28, 34 y 35. Todos ellos cuentan con emplazamientos costeros diferentes y con dataciones radiocarbónicas correspondientes al Holoceno tardío. La composición y proporciones de especímenes identificados para cada conjunto se presentan en la Figura 2.

Figura 2. Frecuencia de especímenes correspondientes a las categorías taxonómicas generales de vertebrados identificadas en diferentes conjuntos zooarqueológicos de la localidad de Heshkaia.
Heshkaia 28 (678 ± 38 años 14C aP; Alunni y Zangrando Reference Alunni and Zangrando2012) es un conchero aislado de escasa extensión (20 m2 aproximadamente) y potencia. Se excavaron 9 m2 y se registró una profundidad máxima de 30 cm en el depósito. Se observaron varios niveles termoalterados. El total de restos óseos recuperados en este sitio es de 11.476 especímenes, de los cuales el 82% corresponde a peces. El resto de los especímenes identificados se reparten entre mamíferos: 4% de guanaco (Lama guanicoe), 1% de cetáceos, 1% de pinnípedos, 10% de mamíferos indeterminados y escasos restos de roedores. También se recuperaron restos de aves (2%).
Heshkaia 34 (981 ± 36 años 14C aP; Alunni y Zangrando Reference Alunni and Zangrando2012) es un conchero compuesto por dos estructuras anulares y cinco montículos de gran extensión que en total cubren una superficie de 1.500 m2 aproximadamente. En este sitio se recuperaron en total 4.143 restos óseos, de los cuales el 48% corresponde a guanacos, el 12% a aves, el 2% a peces, el 1% a pinnípedos y un 36% de mamíferos indeterminados. En proporciones menores al 1% también se recuperaron restos de cetáceos, cánidos, un espécimen de roedor y uno de oveja (Ovis aries), este último en niveles que corresponden al suelo actual que cubre el depósito de conchero.
Heshkaia 35 (656 ± 25 años 14C aP; Alunni y Zangrando Reference Alunni and Zangrando2012) es un conchero aislado de considerable potencia que cubre una superficie de aproximadamente 40 m2. Se excavaron 20 m2 con una profundidad máxima de 60 cm. En la excavación de este sitio se obtuvo un total de 8.214 especímenes óseos, entre los que se encuentran representados un 48% de guanacos, un 26% de peces, un 10% de aves, 14% de mamíferos indeterminados y 1% de roedores. Con valores menores al 1% también están representados pinnípedos, cetáceos y cánidos.
Métodos
Se utilizaron los métodos comúnmente empleados para calcular las diversas medidas de abundancia en los análisis arqueofaunísticos: número de especímenes identificados (NISP), número mínimo de elementos (MNE), número mínimo de individuos (MNI), número mínimo de unidades anatómicas (MAU) y abundancia anatómica relativa (MAU%; Binford Reference Binford1978). La determinación de las clases de edad para guanacos se realizó a partir de dos criterios: (a) el cronograma de erupción, desarrollo y desgaste dentario (Kaufmann Reference Kaufmann2009; Raedeke Reference Raedeke1976); y (b) el cronograma de fusiones óseas (Kaufmann Reference Kaufmann2009).
Se emplearon, asimismo, distintas variables para evaluar la integridad de los conjuntos con el propósito de identificar los procesos tafonómicos que los pudieron haber afectado. Se correlacionó la densidad mineral ósea de guanacos (Stahl Reference Stahl1999) con el MAU% para evaluar los efectos de esta propiedad intrínseca de los huesos sobre la abundancia anatómica. Para ello se tuvo en cuenta la separación entre el conjunto óseo total de guanacos y aquellos huesos que correspondían a ejemplares adultos, según el grado de fusión ósea. Esto responde al supuesto que los elementos de individuos más jóvenes son menos densos y deberían ser menos resistentes a los procesos atricionales (Gutiérrez et al. Reference Gutiérrez, Kaufmann, González, Massigoge and Álvarez2010). Otra de las vías de análisis utilizadas para explorar el estado de las muestras fueron los estadios de meteorización según tres grandes categorías definidas sobre la base de los estadios de Behrensmeyer (Reference Behrensmeyer1978): baja (estadios 0-1), media (estadios 2-3) y alta (estadios 4-5). Al igual que lo que ocurre con la densidad del hueso, la edad de los individuos favorece distintos procesos de deterioro, por lo que se evaluó esta variable distinguiendo el conjunto en general y los especímenes correspondientes a ejemplares fusionados y no fusionados.
Las pautas de transporte fueron evaluadas calculando, en primer lugar, la razón entre el MNE observado y el esperado, siguiendo la propuesta de dividir la carcasa por regiones anatómicas según el volumen de carne y médula que proporcionan (De Nigris Reference De Nigris2004). En segundo lugar, la abundancia anatómica (MAU%) fue correlacionada con los índices de utilidad y el rendimiento energético presentados y discutidos por Alunni (Reference Alunni2018). En este caso se utilizaron los valores de anatomía económica ya conocidos para guanacos (Borrero Reference Borrero, Davis and Reeves1990; De Nigris y Mengoni Goñalons Reference De Nigris, Mengoni Goñalons, Civalero, Fernández and Guráieb2004; Mengoni Goñalons Reference Mengoni Goñalons, Elkin, Madero, Goñalons, Olivera, del Carmen Reigadas and Yacobaccio1996) pero contemplando, además, el retorno energético obtenido por el peso de cada parte y los costos involucrados en el procesamiento de las carcasas. Se empleó el coeficiente de correlación por rango de rho de Spearman con un nivel de significancia de 0,05.
Luego, se analizaron las marcas de procesamiento y patrones de fractura para interpretar las distintas etapas de desorganización del esqueleto que acontecen durante el trozamiento primario (separación de grandes porciones por regiones anatómicas para el transporte al lugar central), el secundario (posterior desarticulación de los elementos) y el final o consumo (descarne, extracción de médula y grasa ósea; Binford Reference Binford1981; Soulier y Costamagno Reference Soulier and Costamagno2017). Para ello, la primera tarea consistió en identificar el tipo de marca cultural (corte, raspado, percusión y machacado) y su ubicación dentro de cada elemento en el que se encuentran.
Posteriormente, se evaluó la fragmentación a partir de diversas variables. Para examinar particularmente las actividades de consumo de médula ósea, se registró la proporción de huesos enteros, la frecuencia de fracturas frescas (green; Morlan Reference Morlan1994) y el número de negativos de impacto para todos aquellos elementos que poseen esta sustancia. Si las actividades de consumo en el sitio estuvieron orientadas al desarrollo de esta actividad, en general se espera que las porciones articulares permanezcan completas o poco fragmentadas. Pero el panorama cambia si estas partes fueron posteriormente procesadas para remover la grasa ósea contenida en el tejido trabecular, dado que para ello es necesario fragmentar las articulaciones para someterlas al hervido. Si bien aún no existen criterios claros que permitan identificar esta práctica, en un intento de explorar su existencia, aquí se cuantificó la frecuencia de porciones completas que poseen tejido esponjoso y que pueden ser sometidas a esta técnica de cocción: cuerpos vertebrales, pelvis, articulaciones y porciones proximales y distales de los huesos largos, carpianos y tarsianos. Luego, se siguió la propuesta de Morin y Soulier (Reference Morin and Soulier2017) y se consignó el porcentaje de marcas de hundimiento (crushing marks) o negativos de impacto y huellas de extracción (tear marks). Las primeras resultan de la fuerza de compresión operando sobre la porción del hueso que permanece en el yunque o que está en contacto con el percutor. En el centro del punto de impacto se visualiza una depresión del hueso (Morin y Soulier Reference Morin and Soulier2017:100, Figuras 1 y 2). Las huellas de extracción (Morin y Soulier Reference Morin and Soulier2017: Figuras 1 y 8) se producen cuando los huesos frescos son fragmentados siguiendo un plano subparalelo relativo a la superficie cortical del elemento, producto de una tensión radial extensiva. Esta última genera dos tipos de marcas de extracción: una “positiva”, que corresponde al fragmento removido, y otra “negativa”, que es la cicatriz que queda en el cuerpo del elemento luego del desprendimiento.
Se emplearon una serie de criterios morfológicos que presentan los especímenes sometidos al procesamiento de la grasa, determinados sobre la base de los estudios experimentales realizados por Morin y Soulier (Reference Morin and Soulier2017) a partir de las extremidades de huesos largos de ciervo colorado (Cervus elaphus). Estos fueron caracterizados únicamente para el fémur, el húmero, la tibia, el radio-cúbito y los metapodios distales, y enfatizan la forma que adquieren los fragmentos epifisitarios o cercanos a las epífisis luego del procesamiento (Morin y Soulier Reference Morin and Soulier2017:Tabla Suplementaria 1). De acuerdo con los autores, un criterio diagnóstico es aquel que haya sido identificado en por lo menos un tercio de los números de elementos conocidos. Estos criterios fueron descriptos y cuantificados y se registró si presentaban huellas de hundimiento o extracción.
Resultados
En los tres conjuntos se reconocieron todas las clases de edad, aunque se destaca la preponderancia de adultos y subadultos en Heshkaia 34 y 35 (Figura 3). En Heshkaia 34 y 35 los cálculos de correlaciones entre la densidad mineral ósea y el MAU% fueron débiles y estadísticamente no significativos tanto para el conjunto total como para los elementos fusionados (Tabla 1). Heshkaia 28, por otra parte, presenta resultados débiles pero estadísticamente significativos, lo que plantea la posibilidad que la composición o estructura física de los distintos huesos haya condicionado parcialmente su preservación ante agentes y procesos naturales posdepositacionales.
Tabla 1. Correlaciones entre MAU%, Densidad Mineral Ósea (DMO) e índices de rendimiento energético (Prueba de Spearman).


Figura 3. Clases de edad identificadas para los restos de guanacos provenientes de Heshkaia 35, 34 y 28.
Con respecto a la meteorización de la muestra general, los especímenes fusionados y los no fusionados se encuentran en su mayoría dentro de los estadios 0-1, en tanto unos pocos fragmentos alcanzan los niveles más avanzados (Figura 4). Esto sugiere condiciones relativamente similares en cuanto al entierro de los conjuntos.

Figura 4. Estadios de meteorización para la muestra total de restos de guanacos, los especímenes fusionados y los no fusionados.
En términos de MNE, los elementos de la región apendicular son siempre más abundantes que los axiales (Tabla Suplementaria 1). Esto es consistente con la presencia predominante de unidades anatómicas que ofrecen médula y proporciones de carne moderadas (extremidades medias) y las que contienen sólo valores marginales de tuétano (calcáneos y falanges; Figura 5). A estos le siguen los metapodios y las extremidades superiores. Las cinturas escapular y pélvica aparecen en frecuencias moderadas, mientras que la representación de los elementos axiales es marcadamente menor, sobre todo las vértebras y las costillas. Por lo tanto, la razón entre MNE observado y esperado sugiere el ingreso más asiduo de unidades anatómicas que contienen sobre todo médula y médula más carne.
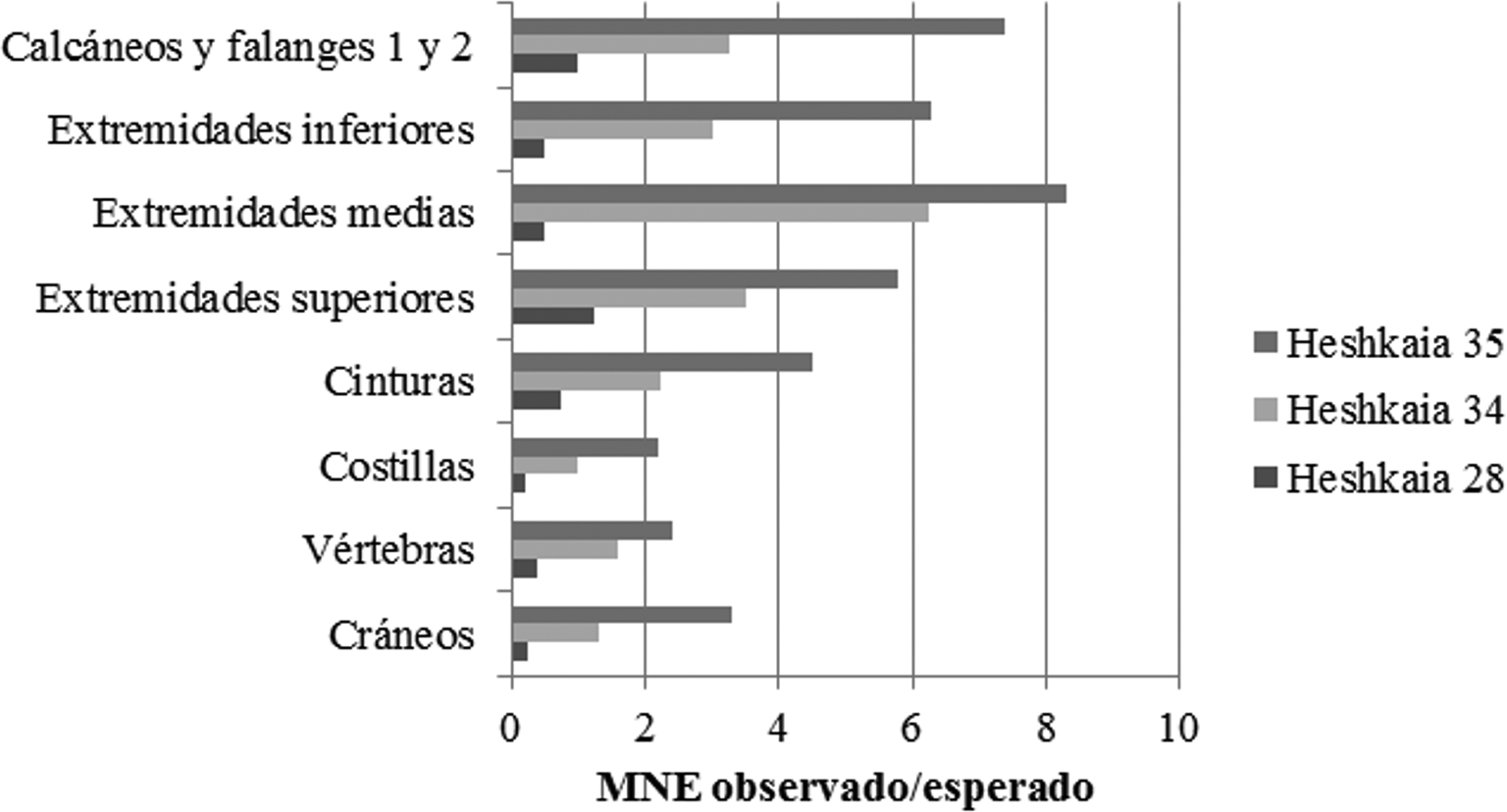
Figura 5. Razón entre MNE observado y esperado por región anatómica en los restos de guanacos pertenecientes al total de Heshkaia 28, 34 y 35.
En parte esto también se expresa en los cálculos de las correlaciones con los índices de rendimiento (Tabla 1). En Heshkaia 28 y 34 las correlaciones entre el MAU% de cada conjunto y el rendimiento energético (kcal/g) muestran resultados negativos, pero débiles y no estadísticamente significativos. Si bien la falta de significatividad podría sugerir ingresos de elementos que no dependan de los rendimientos, se destaca que los datos negativos podrían ser indicadores de cierta tendencia hacia las partes con escasa carne y aporte calórico. Esto es consistente con los cálculos de la razón entre MNE observado y esperado. En Heshkaia 35, por su parte, el panorama es más claro dado que la covarianza resultó ser fuerte, negativa y estadísticamente significativa, lo que suele interpretarse como el ingreso al sitio de porciones con bajo valor económico. Al considerar los índices de rendimiento energético y el índice de utilidad de carne (MUI), Heshkaia 28 y 35 comparten una tendencia similar, de modo que las fuerzas de las correlaciones fueron negativas, fuertes y significativas. Estos cálculos invitan a pensar en estrategias orientadas principalmente a la selección de unidades que posean poca carne y cuyo tiempo de procesamiento no reduzca de manera significativa el rendimiento energético. En cuanto a Heshkaia 34, los resultados continúan siendo negativos al considerar los índices de utilidad (Tabla 1), aunque ninguno es estadísticamente significativo. Esto también permitiría pensar en cierta tendencia hacia el acarreo de elementos con bajo rendimiento, lo que a su vez coincide con la menor frecuencia de porciones que requieren más tiempo de procesamiento y que son ricas en carne, como el costillar y las vértebras. Finalmente, con respecto a la relación entre el índice de médula y el MAU%, los resultados fueron en todos los casos débiles y sin significatividad estadística (Tabla 1).
Con respecto al procesamiento, los restos de guanacos de los tres depósitos expresan frecuencias de huellas culturales afines entre sí, ya que en cada caso estas marcas se reconocieron entre el 17% y el 19% de los especímenes. Su ubicación y orientación permitieron interpretar distintas actividades de procesamiento. En primer lugar, el cuereo pudo ser identificado en los tres sitios a partir de incisiones circulares en los laterales de los metapodios, falanges y articulares (Soulier y Costamagno Reference Soulier and Costamagno2017). El descarne y la remoción de las vísceras de la zona axial fue una práctica registrada por cortes longitudinales en las caras mediales y distales de las costillas, y por incisiones ubicadas en las apófisis y en los cuerpos de las vértebras (todas ellas observadas en Heshkaia 28, 34 y 35). En el esqueleto apendicular, Heshkaia 34 y 35 exhiben el descarne de las cinturas que dejó rastros en las espinas escapulares y sus cuellos y en las crestas ilíacas de las pelvis. En los miembros, en los tres sitios las huellas de corte se localizan a lo largo de las secciones proximales, mediales y distales de las diáfisis, y tienen orientaciones semejantes al patrón que definen distintos autores para el desarrollo de esta práctica (Binford Reference Binford1981; Soulier y Costamagno Reference Soulier and Costamagno2017).
Las evidencias de desarticulación fueron observadas en distintos elementos. En Heshkaia 34 y 35 la separación entre los cráneos y las columnas vertebrales queda evidenciada por marcas de corte localizadas en las crestas lamboideas, en los cóndilos occipitales y en los atlas. En los tres conjuntos, la segmentación de la columna vertebral dejó registros de cortes transversales, en general cortos y profundos, en los procesos articulares craneales y caudales de las vértebras. La separación entre las costillas y las vértebras torácicas está claramente representada por cortes en los extremos proximales de las primeras y por huellas de corte y negativos en los procesos transversos (Heshkaia 28, 34 y 35).
Los rastros del desmembramiento de las cinturas pélvica y escapular están presentes en Heshkaia 34 y 35. En la primera, esta práctica se identifica por cortes en las caras sacro-pélvicas de los iliones que, comúnmente, son atribuidos a la intención de separar el sacro de la pelvis. La remoción de los fémures queda evidenciada por cortes en las superficies articulares de los acetábulos de las pelvis, en sus bordes y lugares de inserción de los músculos cercanos. En los tres conjuntos, la separación de los miembros delanteros y traseros fue identificada por cortes en las articulaciones proximales y distales, así como en carpianos y tarsianos.
En las tres muestras analizadas se observa que el contenido de la cavidad medular de todos los elementos que la poseían, por escaso que haya sido, fue aprovechado para su consumo. Esto no sólo se expresa en todos los huesos que contienen esta sustancia, sino también en la frecuencia de fragmentos de huesos largos indeterminados que exhiben bordes de fractura frescos y negativos de impacto asociados (Tabla 2). Sólo permanecieron completas unas pocas falanges (1 y 2) y un calcáneo en Heshkaia 35 (Tabla 2). Las falanges 3 no fueron procesadas para el consumo, excepto en Heshkaia 34 donde un reducido número de ellas muestra negativos de impacto y bordes de fractura frescos. Los fragmentos mediales de huesos largos acompañan las modificaciones óseas que se manifiestan en los distintos elementos; muchos de estos especímenes presentan estos mismos rasgos de procesamiento asociados.
Tabla 2. Patrones de fractura identificados en huesos de guanacos que poseen grasa ósea.

Con respecto a la grasa ósea, es notable el bajo número de porciones enteras con tejido esponjoso, al tiempo que el porcentaje de huellas de extracción y marcas de hundimiento de material grasoso que exhiben los huesos es elevado (Tabla 2). Las vértebras caudales, los carpianos y los tarsianos son los únicos elementos que, en general, se hallan enteros y con escasos signos de percusión. Sin embargo, se destaca que en Heshkaia 34 se observaron articulares fragmentados con huellas de hundimiento y golpes, lo que indica que sus fracturas fueron intencionales. En el conjunto de Heshkaia 35 se registró, asimismo, un mayor porcentaje de vértebras y metapodios distales completos.
La identificación de los criterios morfológicos relacionados con el procesamiento de la grasa ósea definidos por Morin y Soulier (Reference Morin and Soulier2017), fue baja en Heshkaia 28 (n = 27). En este sitio, los tipos más frecuentes son fragmentos de tibias proximales (M1-T), húmeros proximales (C1-H) y fémures proximales (M1-F). Prácticamente la totalidad de estos especímenes presentan negativos de extracción, aunque el número de huellas de hundimiento es menor. Entonces, si bien algunos restos de guanaco de este conjunto presentan evidencias que podrían sugerir la puesta en práctica del procesamiento de la grasa ósea, es necesario manejar estos datos con cautela, dado que la frecuencia de las partes esqueletarias que puedan dar cuenta de esta actividad es baja.
En cambio, en Heshkaia 34 y 35 se observó una amplia variabilidad de criterios morfológicos (Figuras Suplementarias 1 a 9), los cuales a su vez presentan numerosos rasgos asociados con el procesamiento del tejido esponjoso (Figura 6). En Heshkaia 34 se reconoció un total de 104 criterios y, como puede observarse en la Figura 6, no existen diferencias significativas en la frecuencia de criterios identificados para cada elemento, de modo que todas las porciones con contenido trabecular pudieron haber seguido la misma trayectoria de procesamiento para el consumo. Asimismo, cada criterio presenta marcas de extracción y percusión (Figura 6), evidencias que apoyan la idea del procesamiento intencional de todos estos segmentos morfológicos asignados. Las únicas excepciones las constituyen algunos pocos especímenes distales de los fémures, los húmeros y las tibias (M1-F, M5-F, M3-T, C1-H, M2-H, M3-H, M4-H) que, si bien no muestran impactos asociados, tienen huellas de extracción.

Figura 6. Frecuencias relativas de los diferentes criterios morfológicos: (a) Heshkaia 34; (b) Heshkaia 35.
Al respecto, cabe mencionar que en Heshkaia 35 muchas de estas porciones proximales y distales de los huesos largos que permanecieron enteras presentan rasgos comúnmente asociados al empleo de estos elementos como “machacadores” (Hajduk y Lezcano Reference Hajduk and Lezcano2005). En el conjunto arqueofaunístico del sitio se registraron, en un trabajo conjunto con la Dra. Marianne Christensen (Reference Christensen2016), al menos 31 piezas (65% del total de las articulaciones que están completas) que pueden ser catalogadas como tales, en función de las modificaciones en los bordes de fractura que fueron descriptas por trabajos experimentales (Hajduk y Lezcano Reference Hajduk and Lezcano2005). En este sentido, el empleo de las porciones proximales y distales de los huesos largos como posibles machacadores pudo también ser un factor que haya condicionado las técnicas de fractura en la búsqueda de conseguir soportes adecuados para la producción de tecnología y el desarrollo de actividades específicas. Por lo tanto, éste es un aspecto que requiere ser considerado al analizar las pautas de procesamiento para el consumo de grasa ósea, dado que el uso extendido de estas porciones pudo haber condicionado no sólo la selección de elementos a transportar, sino también las técnicas de preparación de alimentos ejecutadas en el sitio. En otras palabras, tal vez la búsqueda de materias primas óseas (en este caso, las articulaciones) adecuadas para el desempeño de determinadas tareas primó por sobre la necesidad de extraer de ellas la grasa contenida en el tejido esponjoso de las epífisis. Como demuestran las evidencias, esto no implica que dicha tarea no se haya practicado en el lugar, sino que los individuos pudieron haber optado por conservar ciertas piezas como soportes óseos y, en consecuencia, resignar el alimento que éstas ofrecen. De ser correcta esta interpretación, esta menor intensidad en el procesamiento de la grasa ósea en Heshkaia 35 podría no ser el resultado de un contexto económico distinto a lo observado en los otros sitios, sino que la esfera de subsistencia haya estado vinculada de manera integral con la producción de tecnología ósea o formas base específicas.
Discusión y conclusión
Los patrones de mortalidad identificados en Heshkaia 34 y 35 concuerdan con un modelo del tipo prime dominated (Stiner Reference Stiner1990). Bajo este esquema, la selección de presas de mayor tamaño sería una estrategia rentable si se considera que el aporte calórico de estos ejemplares es más elevado que el provisto por crías y juveniles (Stiner Reference Stiner1990). En Heshkaia 28, en cambio, solo están presentes crías, juveniles y subadultos, lo que sugiere que en algunas oportunidades el tamaño de la presa no fue un factor que influyera en el transporte. Se destaca además que la presencia de crías en todos los casos estaría relacionada con la explotación de grupos familiares. Por lo tanto, estos perfiles de mortalidad dan cuenta de cierta diversidad en el tipo de transporte: algunas capturas habrían acontecido en entornos cercanos a los emplazamientos costeros mientras otras ocurrieron bajo escenarios de mayor planificación y logística donde la movilidad entre el valle interior del Río Moat y los ámbitos costeros para el aprovisionamiento de estos recursos podría haberse desarrollado a partir de rangos amplios. Esta variabilidad en las pautas de explotación es esperable en un registro de tiempo promediado donde estarían representadas actividades desarrolladas en diferentes momentos de un ciclo anual.
Los perfiles anatómicos descriptos constituyen otra vía para evaluar el transporte. Se observa un patrón frecuente caracterizado por la preponderancia de los huesos que proveen sólo médula (extremidades inferiores, falanges y calcáneos), que brindan poca carne y una cantidad moderada de médula (extremidades medias, e.g., radio-cúbitos y tibias), o que ofrecen carne y médula (extremidades superiores, e.g., fémures). Los resultados de las correlaciones entre el MAU% y los índices de utilidad y rendimiento acompañan este perfil anatómico e indican el transporte de elementos con bajo retorno energético pero ricos en contenido graso. Esta es una tendencia general que resulta clara en Heshkaia 35, pero que en cierta medida se relativiza al considerar los conjuntos de Heshkaia 28 y Heshkaia 34. En estos dos casos las covarianzas fueron negativas, pero solo fue estadísticamente significativa la relación entre el MUI y el MAU% en Heshkaia 28. Entonces, si bien los tres sitios podrían estar indicando un tipo de transporte similar en donde predominaron partes con bajo contenido cárnico pero alto valor en grasa, es evidente que en Heshkaia 28 y 34 el ingreso de elementos no siempre habría dependido del rendimiento y del peso del hueso. Con respecto al transporte de grasas, las covarianzas débiles y no significativas entre los MAU% y los índices de médula sugieren el acarreo deliberado de esta sustancia sin contemplar las distintas proporciones de lípidos que cada hueso posee, ni su palatabilidad. En este sentido, y considerando el panorama general, la obtención de grasas pudo haber sido prioritaria en la toma de decisiones. Las porciones pudieron haberse transportado sin desperdicio, aunque la relación entre peso y beneficio no fuera redituable (por ejemplo, en el transporte de metapodios, calcáneos y falanges). Se aprecia entonces que se está ante un caso donde prevaleció la elección del tipo de nutriente por sobre la cantidad del mismo.
El registro del tipo, ubicación y frecuencia de las marcas antrópicas sobre las superficies óseas indica que se está tratando con conjuntos que han pasado por todos los estadios de procesamiento: desde el inicio de la secuencia de desarticulación (separación de la cabeza, espinazo, costillar y miembros) hasta los momentos finales relacionados con el trozamiento para el consumo. Se destaca que todos los elementos que poseen médula ósea fueron siempre fragmentados con el propósito de obtener esta sustancia, lo que demuestra que la región apendicular fue siempre aprovechada de manera integral. La importancia que adquirieron las carcasas como fuentes de lípidos se extendió hacia la extracción de la grasa ósea contenida en el tejido trabecular. Las evidencias que indican mayor intensidad de procesamiento provienen del sitio de Heshkaia 34 por exhibir un elevado número de fragmentos asociados a los criterios morfológicos definidos por Morin y Soulier (Reference Morin and Soulier2017): una baja proporción de vértebras y articulaciones completas y un alto número de huellas de procesamiento (Tabla 2; Figura 6). Asimismo, aquí se identificaron astillas con tejido trabecular que poseen negativos de extracción y que pueden ser relacionadas con la tarea de remover la grasa ósea; estos fragmentos también fueron reconocidos en Heshkaia 35, pero con un porcentaje menor (Tabla 2). Cabe destacar que además se registraron tarsianos y carpianos segmentados con negativos de percusión, lo que recuerda al tratamiento que ejercen los Nunamiut sobre estos huesos para preparar guisados (Binford Reference Binford1978).
Esta información sugiere entonces que la composición del paquete a transportar pudo depender de la distribución de los productos que ofrecen las carcasas de los guanacos, de las preferencias alimenticias de los cazadores-recolectores y de la edad de los individuos obtenidos, en los casos de Heshkaia 34 y 35. Sin bien se observa variabilidad en los modos de transportar las presas, el propósito principal de las cacerías de guanacos parece haber sido la obtención de grasas, motor que direccionó en parte las decisiones sobre qué elementos transportar, cuáles abandonar y cómo procesarlos. En este contexto, si el objetivo de los individuos fue aumentar la ingesta de lípidos, pero al mismo tiempo reducir el peso de la carga (sea por la distancia a recorrer, el número de cazadores, etc.), los costos de transporte pudieron ser minimizados por medio de un incremento en el tiempo dedicado a procesar la carcasa en el lugar de matanza. Si bien las vértebras y las costillas tienen alto contenido cárnico, no contienen médula, no son particularmente útiles para manufacturar instrumentos y tienen baja densidad estructural. Además, son elementos pesados, difíciles de procesar y, si no son aprovechados por su grasa ósea (trabecular), pueden ser descartadas inmediatamente luego del descarne en el lugar del procesamiento primario, resignando con ello el producto animal remanente (sensu Lupo Reference Lupo2006). Los huesos largos, en cambio, ofrecen carne, médula y grasa ósea, al mismo tiempo que constituyen buenos soportes para la producción de instrumentos.
Pues bien, este panorama permite repensar algunos aspectos relativos a la movilidad y uso del espacio de los grupos que ocupan ambientes costeros en el sur de Tierra del Fuego. Como se mencionó, los cazadores-recolectores que habitan entornos litorales de climas templados y fríos suelen caracterizarse por un rango de acción limitado (sensu Kelly Reference Kelly1995) hacia espacios terrestres interiores (Binford Reference Binford2001; Kelly Reference Kelly1995), lo que puede responder a diversos factores ambientales (e.g., temperatura efectiva) y sociales (e.g., patrones de asentamiento, demografía, etc.). Sin embargo, los análisis aquí efectuados permiten plantear que la organización espacial de las prácticas de subsistencia en la Bahía Moat pudo haberse desarrollado a partir de un alto dinamismo entre interior y costa: el elevado consumo de guanacos, junto con los patrones de selección y transporte, sugieren desplazamientos hacia el interior del valle para obtener el recurso, actividad que era complementada con la explotación de organismos litorales y marinos. En otras palabras, los espacios interiores y las zonas ecológicas adyacentes al litoral pudieron haber estado integrados logísticamente dentro de un sistema de ocupación centrado geográficamente en la costa (Zangrando et al. Reference Zangrando, Borrazzo, Tivoli, Alunni and Martinoli2014). Esta interpretación marca una diferencia con lo señalado para el canal Beagle central, donde los grupos humanos habrían encontrado en el mar y en la costa la mayor parte de su alimento, mientras que el ámbito interior no suministraba alternativas de consideración, salvo en ciertos sectores circunscriptos donde se podían obtener guanacos (Orquera y Piana Reference Orquera and Piana1999:97). Por lo tanto, para sectores más occidentales del canal Beagle, estos camélidos fueron considerados por estos autores como alternativas que proveían flexibilidad al sistema de subsistencia y su aprovechamiento habría sido principalmente oportunista (Orquera y Piana Reference Orquera and Piana1999:103-105; Vázquez Reference Vázquez2015).
Similares modos de interacción entre humanos y recursos animales en una dinámica costa-interior han sido propuestos para otros espacios costeros de Patagonia insular, en donde las actividades de subsistencia también estuvieron organizadas bajo sistemas esencialmente costeros y marinos. En el entorno boscoso de la costa norte de la Península Mitre se ha planteado un alto dinamismo entre los sectores interiores y costeros, al evaluar la variación temporo-espacial de los campamentos según el uso estacional de guanacos y pinnípedos (Lanata Reference Lanata, Davis and Reeves1990). Entre el Mar de Otway y el Estrecho de Magallanes también se señaló la existencia de contextos costeros desde los cuales los grupos humanos se habrían desplazado por lo menos 70 km hacia el interior para hallar guanacos y transportarlos hacia las localidades costeras (San Román Reference San Román2011). Este autor propone que los cazadores-recolectores marinos que ocuparon los sitios de Punta Santa Ana 1 y Bahía Buena incluyeron dentro de su rango de acción zonas ecotonales de bosque y estepa, y que las carcasas de guanacos fueron transportadas diferencialmente en favor de la región apendicular (San Román Reference San Román2011). Por lo tanto, es probable que la obtención de guanacos hubiera requerido la planificación de cierta estrategia logística relacionada con la partida de un grupo de caza específico y con el empleo de campamentos operativos (sensu Binford Reference Binford1980).
Por último, cabe considerar la posibilidad que si los grupos cazadores-recolectores de la Bahía Moat hubieran organizado desplazamientos hacia el interior en rangos de movilidad variables para la exploración de guanacos —posiblemente mayor a 10 km desde la línea de costa— y que el bosque fueguino no ofrece otros recursos terrestres de consideración alternativos a estos, el procesamiento de grasa ósea y preparación de alimentos a base de este nutriente pudo haber constituido una importante reserva energética. Esto no sólo es factible en términos de aumentar la energía extraída de una carcasa frente a la contingente escasez de otros recursos, sino también porque el manejo planificado de la grasa animal facilita la movilidad (Binford Reference Binford1978; Fitzhugh Reference Fitzhugh, Fitzhugh and Habu2002; Kelly Reference Kelly1995). Esta estrategia pudo haber subsidiado el tiempo de permanencia en sectores interiores de la Bahía Moat con escasa disponibilidad de alimentos.
Agradecimientos
Estas investigaciones fueron financiadas por el proyecto PICT 2013-1011. Queremos agradecer especialmente a Snipe S.A., Alejandro Patiño y Alejandro Winograd por proporcionarnos alojamiento y ofrecernos ayuda constante durante las actividades en el campo. Celina Álvarez Soncini, Hein Bjerck, Hernán De Angelis, Sayuri Kochi, María Paz Martinoli, Jonathan Nye, Germán Pinto Vargas, Vanesa Parmigiani, Suray Pérez, Ernesto Piana, Lucas Sosa, Martín Vázquez y Milagros Poratto participaron en las tareas de excavación. Germán Pinto Vargas organizó los materiales zooarqueológicos para su estudio en laboratorio.
Declaración de disponibilidad de datos
Los materiales a partir de los cuales se derivaron los datos están disponibles en el repositorio del Centro Austral de Investigaciones Científicas (CADIC-CONICET), en la ciudad de Ushuaia, Tierra del Fuego, Argentina.
Materiales suplementarios
Para acceder a los materiales suplementarios que acompañan este artículo visitar https://doi.org/10.1017/laq.2020.27
Tabla Suplementaria 1. Medidas de Abundancia para el Conjunto de Guanacos de Heshkaia 35, 34 y 28.
Figura Suplementaria 1. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para húmeros provenientes de Heshkaia 35
Figura Suplementaria 2. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para radio-cúbitos provenientes de Heshkaia 35
Figura Suplementaria 3. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para fémures provenientes de Heshkaia 34
Figura Suplementaria 4. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para fémures provenientes de Heshkaia 34
Figura Suplementaria 5. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para tibias provenientes de Heshkaia 34
Figura Suplementaria 6. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para tibias provenientes de Heshkaia 34 (continuación de la Figura Suplementaria 5)
Figura Suplementaria 7. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para radio-cúbitos provenientes de Heshkaia 34
Figura Suplementaria 8. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para radio-cúbitos provenientes de Heshkaia 34 (continuación Figura Suplementaria 7)
Figura Suplementaria 9. Criterios morfológicos (sensu Morin y Soulier Reference Morin and Soulier2017) registrados para metapodios provenientes de Heshkaia 34










