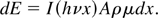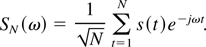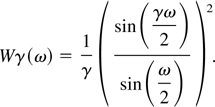1. INTRODUCTION
The study of the interaction of biological cells with ionizing
radiation is very important in biophysics and medical physics in order
to investigate cell behavior and cell response to external factors and
to understand how cells are working even in normal conditions. In
respect to this, the recent advent of high-brightness X-ray sources,
like synchrotrons and laser-plasma sources, has implied important
changes in experimental techniques. Indeed, unlike conventional
sources, which require long exposure times to give high irradiation
doses, such sources allow a very fast deposition of high doses. Hence
the damage can be considered to take place almost instantaneously, and
the cell reaction begins immediately after the damage. This allows a
clear separation of the two steps of dose deposition and repair, which
may not be the case with low brightness sources, and hence allows the
realization of conceptually cleaner experiments.
Also laser-plasma sources are very efficient in the low energy part
of the X-ray spectrum, allowing new wavelengths to be used in
radiobiology experiments. As already pointed out in literature, these
wavelengths are interesting not only because of an increase in the
range of investigated parameters, but also because the low photon
energy implies low energy secondary electrons and hence damage that is
strictly localized around the deposition region (Con
et al., 1977; Goodhead, 1977;
Goodhead & Thacker, 1977). In the case of
the experiments that will be presented in this article, another
important characteristic of low energy X-ray photons has been
exploited, namely, that they may have a very short penetration range in
biological matter. Thus, by carefully choosing the irradiation
wavelength, it is, in principle, conceivable to perform differential
experiments in radiobiology, realizing a preferential dose deposition
to the external cell compartments (cell membrane and cytoplasm) with
little interference with DNA genetic activity taking place in the cell
nucleus. This is important because such external compartments are
strictly related to the cell metabolic activity, which is the subject
of the techniques presented in this article.
In our experiments, a laser-plasma source has been used to irradiate
Saccharomyces cerevisiae yeast cells with soft X-rays. The
experiment has been realized at Rutherford Appleton Laboratory with the
goal of affecting and studying cell metabolism (Turcu et al.,
1994a, 1994b). Yeast cells have largely been used
as a model system for investigating cellular metabolic activity, in
particular because they are eukaryotic cells, hence strictly related to
higher organisms, while at the same time being relatively simple and
unicellular (Tuite, 1964; Fremter, 1983). Moreover they are easy to handle,
nonpathogenic, and can be dehydrated and stay viable.
Teflon (CF2) strips were used as the laser target (Batani
et al., 1995, 1996), emitting a K-shell X-ray spectrum
from F ions centered on 0.9 keV, with a very large absorption
in biological matter. The structure of yeast, characterized by a large
and massive cell wall (Klis, 1994; Batani et al., 1998) contributed to
stopping the X-rays before the nucleus, supporting the idea of inducing
differential damages.
To verify the occurrence of such preferential energy deposition,
X-ray doses to the different cell regions were calculated following the
exponential Lambert–Bouguet–Beer law through a spherical
model of the cell with concentric and differentiated compartments.
The use of high-brightness sources and low-energy X rays might,
however, be of limited use if it is not coupled to new investigation
diagnostic means. Indeed the classical techniques used in radiobiology
experiments to monitor cell response, in particular the colony's
formation ability, necessarily imply a measurement of cellular response
at very late times. Moreover, only those damages capable of an
influence on daughter cells, that is, on following generations, are
registered, which again restricts the investigation to damages of a
genetic nature. When metabolic activity is investigated, other
diagnostics must then be developed.
In our case the experimental technique is based on monitoring the
production of CO2 from the cells with pressure sensors. This
is an old technique (Warburg, 1930), which
may now be rediscovered, taking advantage of recent technological
developments. The analysis can start immediately after irradiation and
can be continued up to several hours. Other advantages are that the
technique is on-line and noninvasive, preserves the cell population,
and can give quantitative information in a continuous way. Also it
allows us to monitor the response of cells to external perturbations.
In the present study such perturbations have been induced by changes in
the feeding conditions (type of sugar) and/or by soft X-ray
irradiation.
The study of CO2 production is interesting because it is
related to energy production (through the glycolytic cycle and the
ATP–ADP balance), being one of the final products of metabolic
activity. There is a complicated series of enzymatic reactions for the
degradation of sugar, which, in eukaryotic cells, includes respiration,
in the presence of oxygen, and fermentation, for instance in yeast
cells, in anaerobic conditions.
Metabolic oscillations in yeast have been evidenced and appear to be
a relevant tool for the investigation of control and feedback
mechanisms at work in biological systems (Nicolis
& Prigogine, 1977; Winfree, 1983;
Cortassa & Aon, 1994). Here, to evidence
their presence, the obtained experimental records of pressure versus
time were analyzed with a spectral technique based on the discrete
Fourier transform (DFT) algorithm (Brillinger,
1981; Ljung, 1987). Oscillations were
indeed observed and it was shown that soft X-ray irradiation changed
their period. Unlike other approaches already used in literature, our
technique not only is, again, noninvasive, but allows metabolic
oscillations to be evidenced in a whole cell population.
In this article, after describing in detail the different parts of
the experimental setup, some preliminary experimental results are
shown. Although some aspects of the several techniques described in
this article have already been, partially, presented in literature, we
think that the combined approach presented here fully exploits their
potentials, and may finally lead to the development of new experimental
techniques in radiobiology.
2. PREPARATION OF THE BIOLOGICAL SAMPLES
The biological specimen was commercial dry Saccharomyces
cerevisiae Hansen yeast (produced by Aboca) that needed to be
hydrated about 1 h before use. The analyzed samples were fixed volumes
(4 ml) of a suspension of cells in deionized water (or phosphate
buffer) with a typical concentration of 2 mg/ml corresponding to
about 2 × 107 cells/ml. The cells were deposited
on a paper filter in a monolayer by filtering in a Venturi tube. The
used filters were Whatman standard cellulose filter papers of grade
1.
After a short period for drying, the filters were put onto a
Hostaphan film so that yeast cells were hosted by a
“sandwich” made of Hostaphan on one side and paper filter
on the other. The thickness of the Hostaphan filter was 1 μm.
For the samples undergoing irradiation, such sandwiches were then
deposited into an automatic handling robot for X-ray exposure (the
Hostaphan film facing the X-ray source) and the dose could be
delivered. Finally the filters were put again in the same quantity of
water (4 ml) and the bottles were sealed and submerged in a thermal
bath at 32°C (temperature variations were lower than 0.1°C).
Different kinds of sugar were added to the suspension as nourishment in
order to study the different metabolic responses. The standard sugar
concentration was 20 mg/ml.
Some intermediate investigations were performed, including the
optimization of the support of the biological target (trying different
paper filters and different preparation procedures). We also verified
that CO2 production from simple cell suspensions was
equivalent to that of sample deposited on the paper filters. To study
specifically fermentation, during some experimental runs an Argon
atmosphere was used instead of air normally present in the bottle, in
order to realize more strict anaerobic conditions and cause a complete
inhibition of respiration.
3. GAS PRODUCTION MEASUREMENT
The overall process is based on the measurement of the pressure
increase in sealed bottles where CO2 production can be
monitored. The bottles were connected to the pressure sensors whose
output is linked to a PC through an Amplicon 16-bit 20 K sample/s
data acquisition board, which has a resolution less than 1.5 μV.
The pressure sensors were connected to a stabilized 10 V power
supply. We used differential silicon pressure sensors (Low Pressure
Sensor type AM5310 DV from Miteco) of high sensitivity (from 0 to 100
mbar, giving 25-mV full-scale output, with an estimated accuracy of 0.1
mV and a sensitivity of about 2.5 mbar/mV). Figure 1 shows the scheme of the data acquisition
system.
Acquisition system of the Miteco Low Pressure Sensor type AM5310
DV.
The sensors were calibrated, showing a very good linearity in the
region of interest (0–100 mbar) and further on. Also no
hysteresis was observed (Batani et al.,
1995; Costato et al.,
1996).
Before analyzing the data, we verified that for control samples (pure
water) the pressure measurements precisely reflected the atmospheric
pressure behavior. Also, to be sure that the pressure increase was
really due to CO2 production, mass spectrometry was used.
Figure 2 shows that the gas produced during
fermentation is CO2 (mass number 44) that was not present in
the initial blank run.
Mass spectrometry of produced gas as a function of mass number. The
initial atmosphere was argon only, corresponding to anaerobic
conditions. The gas produced during fermentation is CO2
(mass number 44) that was not present in the initial blank
run.
The acquisition system allowed the simultaneous recording of data
from 16 different sample bottles. To check data reproducibility we
always had different runs for the same parameters.
4. THE RADIATION SOURCE
The source was obtained with a complex laser system. A Nd laser,
converted to 2ω, was used to pump a dye oscillator followed by dye
amplifiers (pumped by an excimer laser) and by frequency converter
crystals. The laser pulse so obtained was transformed by spatial and
temporal multiplexing in a train of pulses (each 7 ps long), which
underwent a final large amplification in KrF amplifiers, producing UV
radiation at λ = 248 nm. Such laser radiation was focused onto the
target with a f = 9 cm focusing lens producing an intensity of
the order of 5 × 1015 W/cm2. The
overall system is very similar to the one already described by Turcu
et al. (1994a, 1994b).
A relevant element for the success of the experiment was the
availability of a computer driven robot for sample exposure and of a
dose control system. The exposure robot was developed at Rutherford
Appleton Laboratory to irradiate mammal cells with Cu L-shell
X-rays (Turcu et al., 1994a, 1994b).
The dose control system was based on a silicon PIN diode that
measured the X-ray energy produced by each pulse. The PIN was just
beside the cell samples and had the same filters placed before the
cells to exactly measure the same radiation that biological samples
were exposed to. Because a single laser shot was, in general, not
enough to give the required dose, successive PIN signals were
integrated and summed until the required dose was delivered. The
computer then automatically stopped the laser, also giving the number
of laser shots and the histogram of the dose distribution. In a few
minutes the irradiation procedure was completed. Figure 3 shows the scheme of the irradiation
procedure.
Scheme of the irradiation procedure.
The soft X-ray spectrum was also registered (see Section 4), which
allowed an absolute calibration of the PIN diode response. The
conversion factor used to transform the integrated PIN diode signal
(corresponding to a collected electrical charge (in nanocoulombs) to an
absorbed dose (in radians) was 2.6 rad/nC at 0.9 keV. This factor
is dependent on photon energy because of the variation of PIN
sensitivity and the different photon penetration into the biological
material.
5. SPECTRAL CHARACTERIZATION OF THE SOFT
X-RAY SOURCE
The X-ray spectral region, which can be used for the purpose of
producing preferential damage in the cell external compartments, is
quite narrow. It was known that X rays with hν ≈ 1.2
keV, already used for DNA recovery experiments, are characterized by a
large penetration depth in undifferentiated biological material (≈5
μm) and hence can deposit a large quantity of their energy into the
cell nucleus (Turcu et al., 1994a, 1994b). On the other side, the attenuation
coefficient is also low for X rays in the “water window”
region (300–500 eV) as shown in Figure
4.
Mass absorption coefficient (μ/ρ in cm2/g
× 104) of biological material (wall) and biological
material with various percentages of water (corresponding to cytoplasm,
nucleus, average cell) versus photon energy in kiloelectron
volts.
The target material should then be chosen to give emission at
energies as close as possible to the end of water window (corresponding
to the O-absorption edge). Also a K-shell spectrum,
characterized by only a few lines, would be preferable, making the dose
calculations much easier. Unfortunately there are not many elements
with these characteristics. We chose fluorine, or rather Teflon
(CF2), which was produced in thin strips (100 μm thick).
The use of strips drastically reduces the emission of debris, which
could damage the optics and/or the biological samples (Bijkerk et al., 1992). Unfortunately,
CF2 tapes also contain carbon, which emits exactly in the
water window. In the present experiment, such undesired
K-shell C emission has been removed by putting appropriate
filters (2 μm mylar plus 0.2 μm Al) before the biological
samples; these filters also removed scattered UV laser light, which
could produce important biological effects. An additional gas buffer
(He at 1 atmosphere) was present in the interaction chamber with the
main role of further reducing debris effects, but also contributing to
stop softer carbon emission.
In the observed wavelength range, fluorine emits a K-shell
X-ray spectrum (comprised between the fluorine He-α at 731 eV and
the ionization edge for the H-like ion at 1103 eV). The spectra were
recorded with flat crystal minispectrometers (described in Batani et al., 1991) using RbAP crystals
(2d = 26.121 Å) and Kodak DEF film and filtered with 2
μm mylar and 0.2 μm Al. While recording some spectra, a Cu
layer was also added on half the slit: Because Cu is characterized by
the L-absorption edge at hν ≈ 0.993 keV, this
gave a wavelength fiducial on the film, which was useful for spectra
interpretation. Densitometries of the spectra (see Fig. 5) were obtained with a Joyce Laoebl 3CS
Microdensitometer at RAL. Absolute X-ray intensities were calculated
taking into account film sensitivity (Rockett et
al., 1985), filter and buffer gas transparency (Henke, 1986), and crystal reflectivity (Alexandropoulus & Cohen, 1974).
Experimental deconvoluted Teflon spectrum recorded on Kodak DEF film
and corresponding simulated spectrum. Densitometry obtained with a
Joyce Laoebl 3CS Microdensitometer.
6. DOSIMETRY
The information on cell morphology and the analysis of the X-ray
spectrum allow the development of a detailed spherical model of the
cell for the evaluation of the energy absorbed by each compartment of
the microorganism. We recall that, in a previous work (Batani et al., 1998) the following
quantities were measured for the yeast used in this experiment: cell
average radius r0 = 2.58 ± 0.54 μm,
total membrane and wall thickness 0.18 ± 0.02 μm, nucleus
radius rn = 0.95 ± 0.12 μm.
Also their distribution across the cellular population was measured.
The model allows us to separately calculate the doses in the main
cell compartments (wall, cytoplasm, nucleus, considered spherical and
concentric) and to draw a picture of where X-rays are absorbed and
radiation effects are more likely to occur. This is clear progress with
respect to the approach normally used in cellular radiobiology (see,
e.g., Frankenberg et al., 1986) in
which only the radiation penetration depth in the undifferentiated
biological material and the average dose to the whole cell are
considered (in our case, hν ≈ 0.9 keV, the penetration
depth in an undifferentiated average cell, made of 90% water, is about
1.86 μm).
The absorption coefficients and densities of the three cell
compartments are computed according to the following rationale:
- The wall is made of undifferentiated biological material (with no
water) with 7% hydrogen, 22.5% oxygen, 52% carbon, 16.5% nitrogen, 1.5%
sulphur in weight (as deduced from Frankenberg
et al., 1986; Weast & Astle,
1983) with a ρ = 1.6118 g/cm3 average
density;
- The cytoplasm is composed of 95% water and 5% biological material,
with a ρ = 1.0306 g/cm3 average density;
- The nucleus is made of 78% water and 22% biological material, with
a ρ = 1.135 g/cm3 average density.
The cell average density is ρ = 1.06118 g/cm3 (90%
water), comparable to the value (1.08 g/cm3) reported by
Frankenberg et al. (1986).
In the numerical model, cylindrical coordinates are used and the cell
is divided into a series of elemental volumes in each of which the
absorbed doses is numerically computed using the
Lambert–Bouguet–Beer law, that is, the X-ray energy
dE absorbed by a layer with thickness dx, surface
A, density ρ, and absorption coefficient μ is given
by
Here I(hν,x) is the X-ray flux of
photons with energy hν at position x. Hence
I(hν,0) is the spectrum emitted from the plasma,
and, to calculate I(hν,x) it is
necessary to consider the various filters in front of the biological
samples (0.2 μm Al and 2 μm mylar), their transparency, that of
helium, and that of the biological material itself. The doses absorbed
are then computed by summing over the elemental volumes in each
compartment. To assure the precision of the numerical calculus, the
difference between the smallest compartment volume and the sum of all
the elemental volumes inside it is taken to be smaller than a precision
parameter fixed a priori. Also, in the case of a homogenous
spherical cell, an analytical solution can be found which gives the
same result of the finite element calculations.
In this way it is possible to perform a simple study of the influence
of the various parameters (compartment sizes, cell composition,
radiation wavelength) on the absorbed dose.
Figure 6 shows the calculated doses for two
values of the X-ray energy for a cell with average dimension of the
three compartments. “Our” 0.9-keV radiation is compared to
1.5-keV soft X-rays, which would be obtained by focusing the laser onto
an Al target (Turcu et al., 1994a, 1994b), corresponding to Al
K-shell emission. The harder radiation is characterized by a
much bigger energy deposition to the cell nucleus. Moreover, the ratio
between the dose to the wall and the dose to the nucleus is higher for
the softer radiation.
Results of cell dosimetry for two values of X-ray energy: 0.9 keV,
corresponding to Teflon emission, and 1.5 keV, corresponding to Al
K-shell emission. The wall thickness is 200 nm. Doses are in
grays and they are calculated assuming an X-ray flux equal to 1
mJ/cm2 before the filters.
To take into account the variation of the absorbed doses due to the
different sizes of the cells, calculations were performed for various
values of the cell radius. In particular, Figure
7 shows the doses to wall, cytoplasm, and nucleus for different
cell sizes. It is clear how the dose absorbed by the nucleus is higher
in cells with a 2-μm radius, which are likely to be found in the
real cell suspension.
Dosimetry of yeast cells as a function of cell radius calculated for
0.9 keV X-ray radiation and assuming an X-ray flux equal to 1
mJ/cm2 before the filters. Wall thickness and nuclear
radius are assumed to change in a constant ratio with the cell
radius.
7. SPECTRAL ANALYSIS OF CO2
PRODUCTION VERSUS TIME
Often, after 2–3 h, oscillations became evident in
CO2 production in many samples. We then performed a spectral
analysis of these time series to extract the principal harmonic
component of the signal.
It is well known (Brillinger, 1981; Ljung, 1987) that, given a sampled signal
{s(t)}t=1,…N an
estimate of its Fourier transform is given by
It can be proved that the estimate in Eq. (2) is asymptotically (for
N → ∞) unbiased but its variance does not decrease
as N increases. So a smoothing procedure has been used, which
is the only way to improve the poor variance properties of this
estimator. In this way a modification of Eq. (2) is obtained through
convolution with a smooth function of time whose Fourier transform is
frequency window Wγ(ω), which has the property to be
sufficiently narrow to give relevance to a small group of frequencies
at a time, while estimation occurs. In particular a Bartlett window has
been used:
The parameter γ indicates how large the window is: as γ
increases the frequency window gets narrower.
The choice of the parameter γ is critical; some procedures for
optimal estimates of the window width in transfer function estimation
are given by Brillinger (1981) and Ljung
(1987), but they need knowledge of quantities
usually not available in the practical applications, such as the
spectrum of the noise affecting the measurement. So, in this case, a
practical procedure (Ljung, 1987) to solve
the bias-variance trade-off has been used: First initialize the
parameter γ at γ ≅ N/20, where N is
the number of available data; then compute and plot the Fourier
transform estimate for increasing values of γ; as γ increases,
more details of the transform appear, initially due to bias decrease
and, from a certain value of γ on, due to variance increment. The
user should choose the value of γ that he feels will separate the
two phenomena.
8. PRELIMINARY RESULTS
CO2 production was monitored by pressure silicon detectors
with both irradiated and nonirradiated samples. First, we analyzed in
detail CO2 production versus time on a very large amount of
nonirradiated samples. Figure 8 shows the
effects of the different sugars used in the sample suspensions on the
cell metabolic activity, which can be easily evidenced with our
monitoring technique. The different rate of pressure increase shows the
different cell capability of assuming different nutrients, that is,
l-glucose, glucose, d-mannose, and
αd-glucose. In particular, no CO2 pressure
increase is visible for samples fed with l-glucose, which has
a chirality that does not allow this sugar to be metabolized. This
analysis has been performed to show that an accurate CO2
pressure measurement is an efficient tool for the study of cell
metabolic activity; moreover it is affordable, precise, and
reproducible.
CO2 pressure measurements for samples fed with different
kinds of sugars, that is, l-glucose, glucose,
d-mannose, and αd-glucose.
Concerning oscillations, for the nonirradiated sample of Figure 9, a principal frequency component
corresponding to a period of 35.7 ± 2.5 s is clearly visible. A
very similar estimate of the oscillation period has been obtained from
different samples; in all cases the considered samples were 4 ml of a
suspension with 20 mg/ml of αd-glucose and 2 mg/ml
of yeast. Such oscillations were not found in the simultaneously
analyzed control water. The evidence of their metabolic origin is also
confirmed by the analysis of yeast cell suspension fed with
l-glucose, in which we never observed any oscillatory
phenomenon (see Fig. 9). The fact that
oscillations are not found in the control water, and their amplitude is
much larger than the sensor sensitivity, reasonably implies that we are
not facing ghosts or experimental artifacts.
Typical power spectra for a sample of standard cell suspension with
αd-glucose. A principal frequency of 35.7 ± 2.5 s
is visible. Comparison with a suspension fed with l-glucose is
also shown.
For the irradiated samples (doses between 0.5 and 1000 Gy)
oscillations were also observed. All the samples were fed with
αd-glucose, which enhanced the metabolic response. Figure 10 shows the frequency shifting and mixing
as a consequence of irradiation for cells exposed with 20 Gy. The main
frequency peak is shifted toward lower frequencies (giving now a main
component period of 52.6 ± 5.5 s) and the rising of other
frequency components is clearly visible. Such perturbations of spectra
with frequency shifts and the appearance of new frequency components
were registered in all the irradiated samples, which exhibited
oscillations. Anyway whereas a period of about 36 s has been observed
as greatly dominant in most nonirradiated samples, in the case of
irradiated cells no clear correlation between X-ray dose and the
profile of the spectrum has been found for the moment. However, in
general, because the main harmonic component moves toward lower
frequencies, the metabolism of irradiated cells appears to occur at
slightly lower rates, as confirmed also by the observed slower increase
in CO2 pressure. The appearance of many frequencies could
possibly be linked to the activation or exaltation of some less
important glycolytic reactions, as already observed by Hesse (1979).
Effect on CO2 production of irradiation. Comparison
between the power spectra of a nonirradiated cell suspension and a 20
Gy irradiated one; αd-glucose was used as nourishment in
both suspensions.
Finally, Figure 11a, b shows the variation
of pressure as a function of time for a suspension after irradiation
with soft X-rays from Cu and CF2 targets. The scales on the
two figures are the same, showing how the effect is larger for
irradiation with the softer and less penetrating CF2 X-rays.
This confirms that radiation is absorbed at different depths in the
cell and produces different metabolic effects. Again a nonmonotonic
response versus X-ray dose is evidenced in both figures.
Variation of pressure (with respect to nonirradiated cells) as a
function of time after irradiation with soft X-rays from Cu (a) and
CF2 (b) targets. The effect is larger for irradiation with
the softer and less penetrating CF2 X rays. Also, a
nonmonotonic response versus X-ray dose is evidenced in both
figures.
9. CONCLUSIONS
Oscillations related to metabolism have been previously observed in
yeast extract (Aon et al., 1992) and
in single yeast cells (Betz, 1968; Boiteux et al., 1975) with other very
invasive techniques. Because we observe the phenomenon in yeast cell
populations, a synchronization mechanism must take place, which
correlates the different cells and phases them. Such mechanisms have
already been described in literature as shock synchronization and
starvation synchronization (Cameron & Padilla,
1966; Kockova-Kratochvilova, 1990;
Tuite & Oliver, 1991; Chance et al., 1964).
Concerning CO2 production rate versus irradiation dose, we
observed (Batani et al., 1995, 1996) a nonlinear and nonmonotonic behavior, which
is quite different from what is usually found in radiobiology
experiments. The usual approach (e.g., see Tuite
& Oliver, 1991; Turcu et al., 1994a, 1994b) is based on the study of survival
curves, that is the capability of individual cells to develop colonies
after the damage induced by X rays. This only gives the response at
very long times after irradiation, thereby losing all the information
on fast cell response to damages and about cell damages that are not
likely to be transmitted to the genetic descendants.
The highlighting of such early responses is possible thanks to the
use of CO2 monitoring as a technique to follow cell
metabolism and to the use of a high-brightness laser-plasma X-ray
source that allows us to deliver large doses in a short time. This
implies that cell reaction to the damaging factors does not start
before the complete dose deposition: Hence there are not changes in the
biophysics of the studied process during the deposition phase, as
frequently happens in cell radiobiology.
These are only some examples of the sensitivity and reliability of
the diagnostics technique we have developed. Such encouraging results
may be applied to other types of cells in different experiments and may
hence be of general importance to radiobiology and biophysics
studies.
ACKNOWLEDGMENTS
The experiment has been realized at the Rutherford Appleton
Laboratory and fully supported by the E.U. Human Capital and Mobility
“Large Scale Facilities Access” Programme. We also
acknowledge a contribution from INFM, CNR (contract n.
96.00266.CT02.115.27689), and NATO (grant n. GRC961133). The authors
are indebted to D. Stevens and M. Hill (Medical Research Council,
Harwell) for stimulating discussions and technical support. A. Pozzi
thanks the Italian Physical Society (SIF) and A. Masini the INFM for
their grants. Special thanks are due to Cyril Brown, Anthony Parker,
Colin Danson, Chris Edwards, Nick Allen, Katherine Hale, Irene Gray,
Sue Tavender, William Toner of RAL, and to Franco Cotelli and Carla
Lora Lamia of Dipartimento di Scienze Biologiche (Milano). Finally we
would like to recall the memory of Prof. Michele Costato, who was the
main initiator of this work.