INTRODUCTION
Light is one of the most important resources for understorey saplings in forest ecosystems. The relative light intensity on a forest floor is only a few per cent in tropical rain forests (Chazdon & Fetcher Reference CHAZDON and FETCHER1984, Pearcy Reference PEARCY1983, Yoda Reference YODA1974). Understorey saplings must survive in the light-limited conditions until a canopy gap is created, and must grow rapidly after gap formation to outcompete other saplings (Canham Reference CANHAM1988, Poulson & Platt Reference POULSON and PLATT1996, Takahashi Reference TAKAHASHI1996). Most saplings die before growing to the reproductive stage, according to survivorship curves of tree species (Kohyama & Fujita Reference KOHYAMA and FUJITA1981, Nakashizuka Reference NAKASHIZUKA1991). Thus, the sapling stage is important in the life history of tree species (Messier et al. Reference MESSIER, DOUCET, RUEL, CLAVEAU, KELLY and LECHOWICZ1999).
For growth and survival, saplings must have photosynthetic production, which is determined by total leaf area per sapling and assimilation rates per unit leaf area (Lambers & Poorter Reference LAMBERS and POORTER1992, Poorter & Remkes Reference POORTER and REMKES1990, Takahashi et al. Reference TAKAHASHI, SEINO and KOHYAMA2001). Crown architecture is one of the factors affecting sapling leaf area. The crown architecture of saplings can be divided into two extremes: unbranched and highly branched types (Kohyama Reference KOHYAMA1987, Kohyama & Hotta Reference KOHYAMA and HOTTA1990). Highly branched species develop their crowns by allocating carbon more to leaves and branches than to the trunk compared with unbranched species. The developed crown is advantageous for survival in the understorey because of a large assimilative area (Sterck et al. Reference STERCK, MARTINÉZ-RAMOS, DYER-LEAL, RODRÍGUEZ-VELAZQUEZ and POORTER2003). By contrast, the height growth rate of unbranched species is greater than that of highly branched species because a lower biomass increment is required per unit height growth for the unbranched species (Coomes & Grubb Reference COOMES and GRUBB1998, Kohyama Reference KOHYAMA1987). Unbranched species tend to have a small number of large leaves (Kohyama Reference KOHYAMA1987, Takahashi & Rustandi Reference TAKAHASHI and RUSTANDI2006, Takahashi et al. Reference TAKAHASHI, SEINO and KOHYAMA2001). Corner's rules suggested that (1) the stouter the stem, the bigger the leaves and the more complicated their form; and (2) the greater the ramification, the smaller become the branches and their appendages (Corner Reference CORNER1949, Hallé et al. Reference HALLÉ, OLDEMAN and TOMLINSON1978, White Reference WHITE1983). Thus, the combination of the degree of branching and leaf size agrees with Corner's second rule. Large individual leaves compensate for a small number of shoots, and a low leaf support cost (leaf-support tissue mass per unit leaf area) contributes to efficient height growth of unbranched species. Thus, interspecific variation in the degree of branching, as a crown architectural trait, is important for growth and survival of saplings.
Leaf traits are important for carbon gain at the level of individual leaves. Interspecific variation in leaf life span is large spanning a few months to more than 10 y, and relates to the leaf morphology and physiology (Chabot & Hicks Reference CHABOT and HICKS1982, Koike Reference KOIKE1988, Reich et al. Reference REICH, WALTERS and ELLSWORTH1992). Generally, shorter-lived leaves are thinner, and have greater leaf nitrogen per mass (N mass) and assimilative capacity (Reich et al. Reference REICH, UHL, WALTERS and ELLSWORTH1991, Reference REICH, WALTERS and ELLSWORTH1992). N mass is positively correlated with the mass-based assimilation capacity (Ellsworth & Reich Reference ELLSWORTH and REICH1992, Reference ELLSWORTH and REICH1993), because photosynthetic enzymes, such as RuBP carboxylase, contain large quantities of nitrogen (Evans Reference EVANS1989, Seemann et al. Reference SEEMANN, SHARKEY, WANG and OSMOND1987). The long photosynthetically active period of long-lived leaves compensates for their low assimilative capacity (Kudo Reference KUDO1999). Given the high assimilation capacity of shorter-lived leaves, the height growth of saplings with shorter-lived leaves is expected to be greater in the light. However, saplings with short-lived leaves must produce many leaves to maintain the current sapling leaf area. If the carbon gain of a sapling is lower than the minimum biomass required to maintain the current sapling leaf area, the growth of the sapling gradually decreases. Such high minimum growth is disadvantageous for the survival in understorey conditions where carbon gain is limited (King Reference KING1994). Therefore, the crown architectural and leaf traits strongly influence the growth and survival of saplings.
Interspecific variation in the degree of branching may correlate with that in the leaf traits. If unbranched species have short-lived large leaves, their saplings can grow in height rapidly in canopy gaps by the combination of large carbon allocation to the trunk and large assimilative capacity of leaves. By contrast, if highly branched species have long-lived small leaves, combination of a large assimilative area with long-lived leaves is advantageous for survival in light-limited understorey conditions. Takahashi et al. (Reference TAKAHASHI, SEINO and KOHYAMA2005) examined eight deciduous broad-leaved species and observed that mid-successional unbranched species tend to have larger leaves with higher N mass and thinner leaves as compared with highly branched late-successional species. However, the relationship between the degree of branching and leaf traits other than leaf size was rather weak (Takahashi et al. Reference TAKAHASHI, SEINO and KOHYAMA2005), probably because the small interspecific variation in leaf life span due to the deciduous habit reduces the interspecific variation of leaf traits such as leaf mass per area (cf. Klooster et al. Reference KLOOSTER, THOMAS and STERCK2007). Thus, examination of evergreen species with large variation in leaf life span is important to understand the relationship between degree of branching and leaf traits associated with the leaf life span.
This study clarified (1) if leaf traits correlate with degree of branching and (2) if both degree of branching and leaf life span correlate with height growth rates by investigating interspecific variation in leaf traits of leaf life span, leaf nitrogen, leaf mass per area and leaf size in relation to degree of branching in eight evergreen species in a tropical rain forest, Indonesia.
STUDY SITE
This study was done in a tropical montane rain forest in Gunung Halimun National Park (6°44′S, 106°32′E, 1100 m asl), West Java, Indonesia. The annual mean rainfall was 3869 mm, recorded at the office of a tea plantation near the study site during 1998 to 2002. The rainy season is from October to April, and the monthly rainfall exceeds 300 mm during this period. The monthly mean temperatures were between 24 °C and 31 °C in 1998 and 1999 (Takahashi & Mikami Reference TAKAHASHI and MIKAMI2006).
The forest of the Gunung Halimun National Park was a reserve, and, therefore, anthropogenic effects on the vegetation were absent. Two permanent plots, 100 × 100 m in area, were established on Mount Kendung (Suzuki et al. Reference SUZUKI, YONEDA, SIMBOLON, MUHIDIN, WAKIYAMA, Yoneda, Sugardjito and Simbolon1997, Reference SUZUKI, YONEDA, SIMBOLON, FANANI, NISHIMURA, KIMURA, Simbolon, Yoneda and Sugardjito1998). The number of tree species greater than 4.8 cm dbh in the two plots was 116 and 103 species, the total basal area was 36.3 m2 ha−1 and 35.4 m2 ha−1, respectively, and the maximum tree heights were 54 m and 37 m, respectively. The dominant species in the two plots were Altingia excelsa Noronha, Castanopsis acuminatissima A. DC. ex Hance, Schima wallichii Choisy and Quercus lineata Blume (Suzuki et al. Reference SUZUKI, YONEDA, SIMBOLON, MUHIDIN, WAKIYAMA, Yoneda, Sugardjito and Simbolon1997, Reference SUZUKI, YONEDA, SIMBOLON, FANANI, NISHIMURA, KIMURA, Simbolon, Yoneda and Sugardjito1998). Several ginger species dominated the forest floor (Nishimura & Suzuki Reference NISHIMURA and SUZUKI2000, Takahashi Reference TAKAHASHI2004).
MATERIALS AND METHODS
Field methods
Eight species were chosen for analysis (Table 1), because these species were abundant compared with other species. The crown architecture differed between the eight species, i.e. their branching structure varied between unbranched and highly developed types (Takahashi & Rustandi Reference TAKAHASHI and RUSTANDI2006). Castanopsis acuminatissima was the most-branched species, and Macaranga semiglobosa J. J. S. was the most unbranched species (Table 1). Adult size differed among the eight species (Table 1). Lasianthus tomentosus Bl. and Symplocos cochinchinesis (Lour.) Moore. ssp. laurina (Retz.) Noot. are shrub species. Macaranga semiglobosa is a subcanopy tree species. Clidemia hirta (L.) D. Don is an invasive subshrub species, native to Central and South America (Wester & Wood Reference WESTER and WOOD1977). The other species were canopy tree species.
Table 1. Characteristics of saplings of eight species in a tropical montane forest, Indonesia. Species are arranged in descending order by degree of branching. U = understorey conditions, G = canopy gap conditions. C = Canopy species, SC = subcanopy species, S = shrub species, SS = subshrub species.
Takahashi & Rustandi (Reference TAKAHASHI and RUSTANDI2006) examined the crown allometry and height growth of understorey and gap individuals shorter than 2.5 m of the eight species used in this study. They examined about 40 individuals of each of the eight species. In this study, we examined the leaf traits of these understorey and gap individuals in relation to the degree of branching found by Takahashi & Rustandi (Reference TAKAHASHI and RUSTANDI2006).
All leaves were counted for each individual in March 2003, and were marked with a paint marker. All leaves of each individual were counted again and newly emerged leaves were marked with a paint marker in October 2003 and March 2004. The leaf life span (LLS, y) of each individual was calculated by the following equation (King Reference KING1994).
where NM is the mean number of leaves per plant, RP is the leaf production rate per plant and RL is the leaf loss rate per plant. The number of leaves in March 2003 and March 2004 was used for the calculation.
We collected several leaves from the leader shoot of each individual in March 2004. The leaves were oven-dried for at least 24 hour at 80 °C and were weighed. The leaf area was measured by using the computer graphic software NIH Image and a scanner. Petioles were not included in estimated leaf areas. The mean leaf area of individual leaves was calculated for each species. After the leaf area was measured, the leaves were ground into a powder and the nitrogen concentration per leaf dry mass (N mass) was determined by using a C-N analyser (MT-1600, Yanaco Co., Tokyo). The nitrogen concentration per leaf area (N area) was calculated by the product of N mass and leaf mass per area (LMA).
Statistical analysis
Takahashi & Rustandi (Reference TAKAHASHI and RUSTANDI2006) showed three allometric relationships relating to crown development: total leaf area per individual plant (A L), crown projection area (A C), and total length of trunk and branches (L TB), each against trunk height. Trunk height was defined as vertical distance from the ground to the highest apex, i.e. trunk height is equivalent to plant height. L TB was used to express the degree of branching at a given trunk height. Takahashi & Rustandi (Reference TAKAHASHI and RUSTANDI2006) compared the interspecific differences of the crown allometry at trunk heights 45, 90 and 180 cm. The interspecific differences were pronounced at 180 cm because more species differences were found on slopes of the log-log allometry than on the intercepts. Thus, in this study we used L TB at the trunk height of 180 cm to represent the degree of branching of each species. Takahashi & Rustandi (Reference TAKAHASHI and RUSTANDI2006) also showed that A L and A C were positively correlated with L TB at trunk height 180 cm, indicating that highly branched species have well-developed crowns.
The effects of leaf life span, canopy conditions (gap vs understorey) and the interaction on leaf traits LMA, N mass, N area, leaf size and height growth rate were examined by general linear models. The mean leaf area of individual leaves was used as the leaf size for each species in each canopy condition. The canopy condition was treated as a categorical variable. The effects of L TB at trunk height 180 cm, canopy conditions (gap vs understorey) and the interaction on leaf traits LMA, N mass, N area, leaf size, leaf life span and height growth rate were also examined by general linear models.
RESULTS
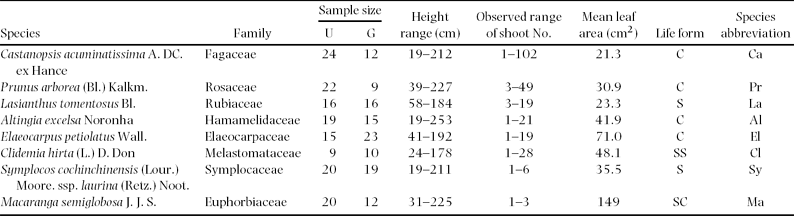
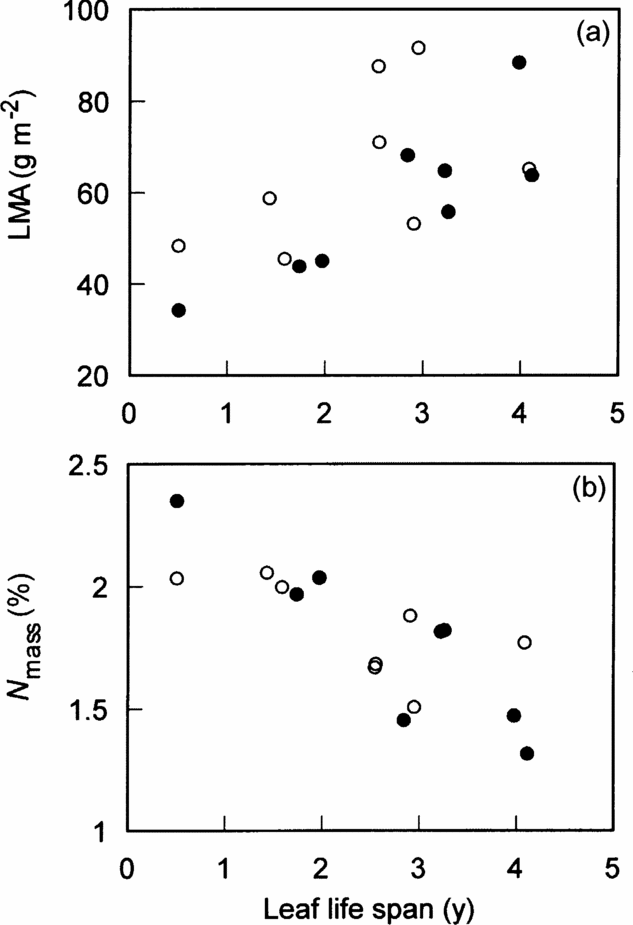
Interspecific variation in two leaf traits of the four was significantly correlated with leaf life span. LMA was positively (P < 0.01) and N mass was negatively (P < 0.01) correlated with leaf life span (Figure 1a, b, Table 2). However, LMA and N mass were hardly affected by canopy conditions (gap vs understorey) and the interaction between canopy conditions and leaf life span (Figure 1a, b, Table 2). Canopy conditions (gap vs understorey), leaf life span and the interaction did not affect on leaf size and N area (Table 2). Thus, shorter-lived leaves had higher N mass and lower LMA, irrespective of canopy conditions.
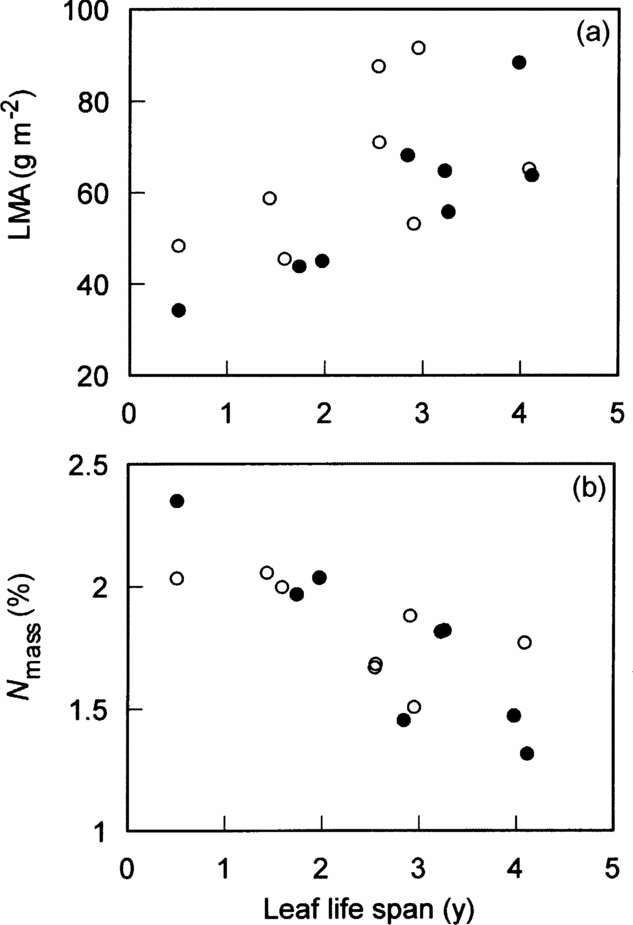
Figure 1. Relationships of LMA (a) and N mass (b) with leaf life span among eight species in a tropical montane forest, Indonesia. Solid and open circles represent understorey and gap individuals, respectively. The results of statistical tests are shown in Table 2.
Table 2. The results of general linear model for the effects of canopy conditions (gap vs understorey), leaf life span (LLS) and the interaction on leaf size of individual leaves, leaf mass per area (LMA), leaf nitrogen concentration per leaf mass (N mass), per leaf area (N area) and height growth rate (HG). Leaf area was used as leaf size. Leaf size and N mass were ln-transformed and square-root arcsine-transformed before the analysis, respectively. F 1,12-values are shown with the significance level. Whole-model R 2 is also shown on the bottom of each column. ** = P < 0.01, *** = P < 0.001. n = 16. Two significant relationships are shown in Figures 1a (LMA), 1b (N mass) and 3b (HG).
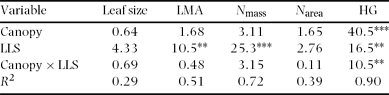
The five leaf traits were examined in relation to canopy conditions and the degree of branching expressed as the L TB at 180 cm trunk height. Leaf life span, LMA, N mass and N area were not significantly affected by canopy conditions and L TB (Table 3). However, leaf size was negatively correlated with L TB (Figure 2, Table 3, P < 0.01). Thus, the relationships of leaf traits and degree of branching were weak, except for those involving leaf size.
Table 3. The results of general linear model for the effects of canopy conditions (gap vs understorey), the total length of trunk and branches at 180 cm trunk height (L TB) and the interaction on leaf size of individual leaves, leaf life span (LLS), leaf mass per area (LMA), leaf nitrogen concentration per leaf mass (N mass), per leaf area (N area) and height growth rate (HG). Leaf area was used as leaf size. Leaf size and N mass were ln-transformed and square-root arcsine-transformed before the analysis, respectively. F 1,12-values are shown with the significance level. Whole-model R 2 is also shown on the bottom of each column. F 1,10-values in parentheses are the statistical results for the height growth rates of the seven species other than subshrub Clidemia hirta (n = 14). * = P < 0.05, ** = P < 0.01. n = 16. Two significant relationships are shown in Figures 2 (leaf size) and 3a (HG).

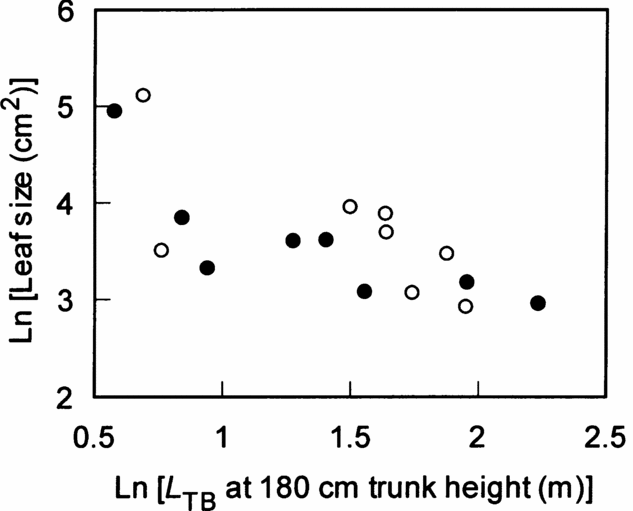
Figure 2. Relationship between leaf size and total length of trunk and branches (L TB) at 180 cm trunk height among eight species in a tropical montane forest, Indonesia. Solid and open circles represent understorey and gap individuals, respectively. The results of statistical tests are shown in Table 3.
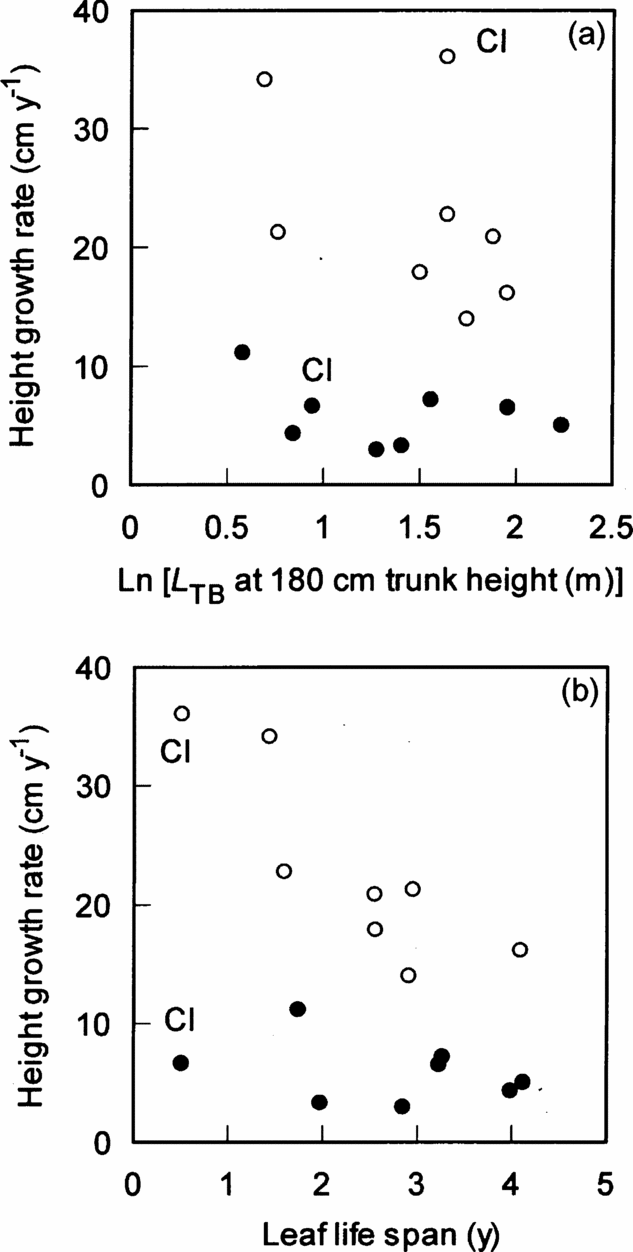
The height growth rate of understorey individuals of the eight species was low, irrespective of L TB and leaf life span (Figure 3a, b). However, the effects of canopy conditions (P < 0.001), leaf life span (P < 0.01) and the interaction (P < 0.01) significantly affected the height growth rate (Figure 3b, Table 2), indicating that species with shorter-lived leaves grew faster in gaps. The effects of the degree of branching (L TB) and the interaction between L TB and canopy conditions were not significant, indicating that the height growth rate increased in gaps, irrespective of L TB (Figure 3a, Table 3). However, when the invasive subshrub species Clidemia hirta was excluded, the effect of L TB on the height growth was significant in the other seven species (F 1,10 = 5.9, P < 0.05).
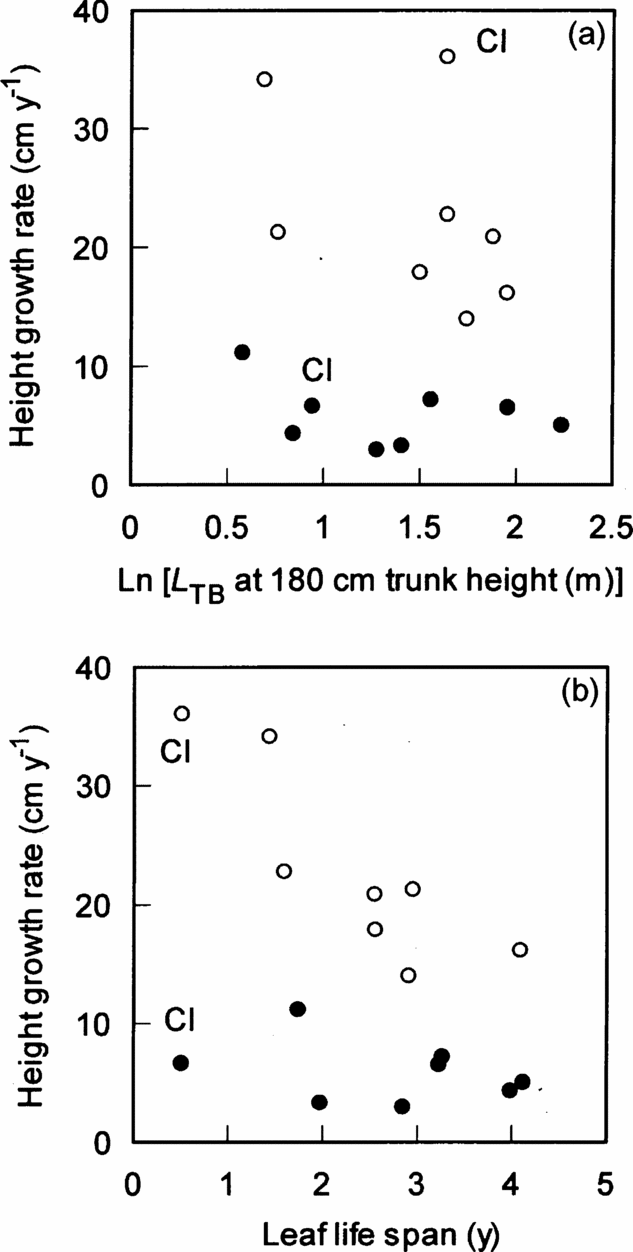
Figure 3. Relationships of height growth rate with ln-transformed L TB (a) and leaf life span (b) among eight species in a tropical montane forest, Indonesia. Solid and open circles represent understorey and gap individuals, respectively. The results of statistical tests are shown in Tables 2 and 3. Cl = Clidemia hirta.
DISCUSSION
In this study, leaf life span was positively correlated with LMA and negatively correlated with N mass among the eight species, irrespective of canopy conditions (gap and understorey). These relationships are general patterns, irrespective of growth form, plant functional type or biome (Wright et al. Reference WRIGHT, REICH, WESTBY, ACKERLY, BARUCH, BONGERS, CAVENDER-BARES, CHAPIN, CORNELISSEN, DIEMER, FLEXAS, GARNIER, GROOM, GULIAS, HIKOSAKA, LAMONT, LEE, LEE, LUSK, MIDGLEY, NAVAS, NIINEMETS, OLEKSYN, OSADA, POORTER, POOT, PRIOR, PYANKOV, ROUMET, THOMAS, TJOELKER, VENEKLAAS and VILLAR2004), because long-lived leaves must be mechanically strengthened and short-lived leaves need much nitrogen for a positive carbon balance (carbon gain by photosynthesis minus leaf construction cost) during the leaf life span. Although this study examined only eight species, the leaf traits in relation to leaf life span agreed with previous studies (Mediavilla & Escudero Reference MEDIAVILLA and ESCUDERO2003, Reich et al. Reference REICH, UHL, WALTERS and ELLSWORTH1991, Reference REICH, WALTERS and ELLSWORTH1992; Shipley et al. Reference SHIPLEY, LECHOWICZ, WRIGHT and REICH2006).
We expected a relationship between leaf traits and crown architectural traits, i.e. unbranched species have short-lived large thin leaves with high assimilative capacity (low LMA and high N mass) and highly branched species have long-lived small thick leaves with low assimilative capacity (high LMA and low N mass). However, the degree of branching was not correlated with leaf traits, except for leaf size, indicating weak relationships between crown architectural and leaf traits.
Shade tolerance of tropical trees is positively correlated with LMA and leaf life span, and is negatively correlated with N mass and maximum photosynthetic rate (A max) (Baltzer & Thomas Reference BALTZER and THOMAS2007, Poorter & Bongers Reference POORTER and BONGERS2006, Sterck et al. Reference STERCK, POORTER and SCHIEVING2006a). Shade tolerance is also positively correlated with crown architectural traits, such as wood density, crown projection area and total leaf area per plant (Henry & Aarssen Reference HENRY and AARSSEN2001, Sterck et al. Reference STERCK, VAN GELDER and POORTER2006b, Takahashi et al. Reference TAKAHASHI, SEINO and KOHYAMA2001). Leaf traits relate to light requirement for each species because high A max is associated with a high respiration rate, which is disadvantageous for survival in understorey conditions. Total leaf area per plant relates to assimilation area per plant (Takahashi et al. Reference TAKAHASHI, SEINO and KOHYAMA2001). Thus, in this study, the negative correlation between height growth and leaf life span for gap individuals of the eight species is because assimilative capacity per leaf mass is greater for leaves with a shorter life span (Koike Reference KOIKE1988, Reich et al. Reference REICH, UHL, WALTERS and ELLSWORTH1991). Also, the negative correlation between height growth rate and the degree of branching is, but only if Clidemia hirta was excluded, due to lower biomass increment required per unit height growth for unbranched species than highly branched species (Kohyama Reference KOHYAMA1987). Therefore, crown architectural and leaf traits influence the height growth of the eight species, except for Clidemia hirta in terms of the effect of the degree of branching.
Clidemia hirta is a weedy aggressive invasive species which shades out all vegetation below it, and often dominates in disturbed sites such as along trails and after wild fire (Wester & Wood Reference WESTER and WOOD1977). In addition, the rate of the increase of height growth rate in gaps is greater in Clidemia hirta than the seven other species (Takahashi & Rustandi Reference TAKAHASHI and RUSTANDI2006), nevertheless this species is not unbranched. One would expect that invasive species grow faster in introduced sites than the native sites because invasive species lack coevolved enemies in their new ranges, i.e. the enemy-release hypothesis (Elton Reference ELTON1958, Mitchell & Power Reference MITCHELL and POWER2003). However, Parker et al. (Reference PARKER, BURKEPILE and HAY2006) examined more than 100 species to test this hypothesis, and found that native herbivores suppressed exotic plants. Thus, the enemy-release hypothesis is still controversial. Genetic shift is another hypothesis to explain successful invasion of invasive species, i.e. native and introduced genotypes differ genetically in ways that lead to differences in biomass allocation, growth or photosynthesis (Blossey & Nötzold Reference BLOSSEY and NÖTZOLD1995). However, DeWalt et al. (Reference DEWALT, DENSLOW and HAMRICK2004) revealed that there were no significant differences in relative growth rates, maximal photosynthetic rates and LMA of Clidemia hirta between introduced and native sites. Thus, the high growth rate of Clidemia hirta in gaps cannot be explained by the enemy-release and genetic-shift hypotheses. The assimilative capacity of Clidemia hirta leaves is probably the greatest among the eight species because of their short leaf life span associated with the highest N mass (Figure 3b). Furthermore, Clidemia hirta does not need to construct a thick stem associated with high wood density because Clidemia hirta is a perennial subshrub (i.e. almost herbaceous species). There is no doubt that the high growth rate of Clidemia hirta in gaps is caused by its low wood density and thin stem in addition to the high assimilative capacity of the leaves. Therefore, Clidemia hirta is an exceptional species for the analysis of the relationship between height growth rate and the degree of branching.
Height growth of understorey individuals of the eight species was low, irrespective of the degree of branching and leaf life span, unlike the interspecific variation in gap individuals. A similar pattern was also reported by Coomes & Grubb (Reference COOMES and GRUBB1998) for 12 Amazonian species, i.e. height growth rates of understorey saplings were similar among the 12 species, irrespective of the degree of branching, while unbranched species tended to grow faster in gaps than branched species. Thus, the interspecific variations of height growth between gap and understorey contrast sharply with those of the leaf traits (LMA and N mass) in that the interspecific variations were explained well by the leaf life span, irrespective of canopy conditions. Similar growth rates of understorey individuals among the eight species were probably because of the shortage of light resource for assimilation, i.e. species could not assimilate at the maximal rate. Unbranched species can potentially grow in height faster than highly branched species because lower biomass is required for unit height growth. However, total leaf area of unbranched species is smaller than highly branched species in understorey conditions (Takahashi & Rustandi Reference TAKAHASHI and RUSTANDI2006), which reduces carbon gain per plant in unbranched species. Thus, the lower cost for height growth was counterbalanced by a smaller total leaf area in unbranched species, which resulted in similar height growth rates among the eight species with different degrees of branching in understorey conditions.
Of the several leaf traits examined, only individual leaf size was negatively correlated with L TB, i.e. Corner's second rule – the greater the ramification, the smaller the branches and their appendages (Corner Reference CORNER1949). Such weak relationships between the degree of branching and leaf traits (LMA and N mass) were also observed in saplings of deciduous broad-leaved species, while Corner's rule was recognized in these species (Takahashi et al. Reference TAKAHASHI, SEINO and KOHYAMA2001, Reference TAKAHASHI, SEINO and KOHYAMA2005). Ackerly & Donoghue (Reference ACKERLY and DONOGHUE1998) found that 17 Acer species in the understorey were arranged by independent morphological suits, i.e. Corner's rule and crown allometry. They argued for relatively independent evolution of two suits of traits. Although the results of this study cannot be simply compared with those of Ackerly & Donoghue (Reference ACKERLY and DONOGHUE1998) because the leaf and crown traits examined were different between the two studies, the negative relationship between leaf size and degree of branching may be an evolutionarily robust character. The number of leaves per plant is regulated by the number of shoots per plant. In unbranched species with a fewer shoots, leaf size of individual leaves should be larger to increase the total leaf area per plant. If the leaf size of highly branched species with many shoots is as large as that of unbranched species with a few shoots, total leaf area per plant increases considerably, which decreases the whole-plant carbon gain due to severe self-shading within a crown (Pearcy & Yang Reference PEARCY and YANG1998, Takenaka et al. Reference TAKENAKA, TAKAHASHI and KOHYAMA2001). King (Reference KING1998) observed that first branch height was positively correlated with leaf size in 70 species in tropical rain forests, showing that leaf size correlates with crown architecture. Thus, strong selection pressure could drive on the leaf size/number and the degree of branching. Therefore, the negative relationship between the degree of branching and leaf size (i.e. Corner's rule) is efficient for whole-plant carbon gain, irrespective of leaf life span.
This study concludes (1) that degree of branching did not correlate with the leaf traits except for leaf size in the eight species (only a negative relationship between the degree of branching and leaf size was recognized) and (2) that the height growth rate of gap individuals was negatively correlated with leaf life span for the eight species and with the degree of branching for the seven species other than the subshrub Clidemia hirta. We predicted that the shade tolerance of the species would be highest if highly branched species have long-lived small leaves with high LMA and low N mass, and the growth in gaps is highest if an unbranched form is coupled with short-lived large leaves with low LMA and high N mass. Thus, the relationships between the crown architectural and leaf traits were weaker than our prediction in the eight species examined. It is necessary to examine how such a weak correlation between leaf and crown architectural traits contributes to the regeneration of the eight species and the maintenance of species diversity in this forest.
ACKNOWLEDGEMENTS
We are grateful to Agus Rustandi, Muhammad Amir and Aden Muhidin for their assistance in the field. Thanks are also due to the staff of the Research and Development Center for Biology, Indonesian Institute of Science for their kind support. This study was made by permission of the Indonesian Institute of Science (LIPI). The field study was financially supported by a grant from the Ministry of Education, Culture, Sports, Science and Technology, Japan (No. 14255003, Leader T. Kohyama).