INTRODUCTION
Most protected areas in African savannas are heterogeneous with many vegetation types. The occurrence of many vegetation types offers varied foraging opportunities to large herbivores allowing them a wide choice of habitats resulting in their diverse distribution. When large herbivores utilize a wide range of vegetation types they are considered to be less selective (O'Kane et al. Reference O'KANE, DUFFY, PAGE and MACDONALD2013). A study of the use of heterogeneous vegetation by large herbivores, particularly mixed feeders with their unique ability to feed on both grass and browse, offers an opportunity to determine the extent of their selectivity.
The impala Aepyceros melampus Lichtenstein is a mixed feeder or grazer-browser that is widely distributed in most protected areas in sub-Saharan Africa (Grobler et al. Reference GROBLER, HAYTER, LABUSCHAGNE, NEL and COETZER2017). The ability of the impala to switch from grazing to browsing when grass quality declines (Klein & Fairall Reference KLEIN and FAIRALL1986, Kleynhans et al. Reference KLEYNHANS, JOLLES, BOS and OLFF2011, Smithers Reference SMITHERS1983, Wronski Reference WRONSKI2003), means it is able to exploit forage resources in heterogeneous environments (Shannon et al. Reference SHANNON, MACKEY and SLOTOW2013). Thus, the impala is expected to forage in vegetation types with both good grazing and browsing. The impala occupies grasslands in the wet season and then switches to vegetation types with browse in the dry season (Jarman Reference JARMAN1972), and browses more often in the dry than the wet season (O'Kane et al. Reference O'KANE, DUFFY, PAGE and MACDONALD2011).
In African savanna with distinct seasonality the impala diet switch from grass to browse occurs during the interface of wet and dry season. The quality of grass and browse at the time of diet switching has not been extensively studied. However, browse contribution to the impala diet increases during the dry season (Hansen et al. Reference HANSEN, MUGAMBI and BAUNI1985). The impala, as a small ruminant, selects the most digestible components of forage such as newly sprouting shoots and leaves and avoids coarse plant material such as stems and sheaths (Gordon & Illius Reference GORDON and ILLIUS1996), to meet its nutritional requirements (McNaughton Reference MCNAUGHTON1990). Newly sprouting shoots and leaves have high nutrient content and low fibre content (Macandza et al. Reference MACANDZA, OWEN-SMITH and CROSS2004, Sensenig et al. Reference SENSENIG, DEMMENT and LACA2010). During the dry season soil moisture declines to levels that can no longer support grass sprouting (Mandinyenya pers. obs.), triggering the switch from grazing to browsing by the impala.
In heterogeneous environments the impala is expected to use vegetation types with more nutritious foraging patches and avoid those with unpalatable grasses and browse (Fryxell et al. Reference FRYXELL, WILMSHURST and SINCLAIR2004). Nitrogen is a good proxy for grass and browse quality (Zengeya et al. Reference ZENGEYA, MUTANGA and MURWIRA2013), together with phosphorus and calcium which are important nutrients for herbivore growth, reproduction and lactation (Grasman & Hellgren Reference GRASMAN and HELLGREN1993, Murray Reference MURRAY, Sinclair and Arcese1995). In vitro gas production and fibre (acid detergent fibre) content are important indicators of rumen fermentation of both grass and browse (Sebata et al. Reference SEBATA, NDLOVU and DUBE2011). The decline in grass quality from wet to dry season results in low availability of digestible nutrients and hence low herbivore energy intake (Shannon et al. Reference SHANNON, MACKEY and SLOTOW2013).
Although the impala prefers leafy short grasses (O'Kane et al. Reference O'KANE, PAGE and MACDONALD2014), it is able to selectively pluck individual leaves from tall grasses, allowing it to feed on grasses of different heights (Arsenault & Owen-Smith Reference ARSENAULT and OWEN-SMITH2008). However, tall grass tends to obscure its grazing sight lines and can conceal predators (Hopcraft et al. Reference HOPCRAFT, MORALES, BEYER, BORNER, MWANGOMO, SINCLAIR, OLFF and HAYDON2014). Thus, an understanding of how grass height influences the impala use of heterogeneous vegetation is important in getting an insight into its anti-predator behaviour.
We assessed habitat use, in terms of vegetation type use of the impala in a heterogeneous protected area, ZNP. We hypothesized that: (1) the impala density varies with vegetation type, (2) the impala switch from grazing to browsing coincides with a decline in grass nutritive value to below that of browse, and (3) grass height varies with vegetation type.
METHODS
Study site
We conducted the study from January 2013 to December 2015 in the Zambezi National Park (ZNP) in Victoria Falls, Zimbabwe (17°53'S, 25°41'E). Zambezi National Park is a 560 km2 protected area characterized by gently undulating terrain between 600 and 1200 m asl. Rainfall is highly seasonal, with an annual mean of 650 mm, which falls almost exclusively between October and April. The year can be divided into three seasons: wet (December–April), cool dry (May–August) and hot dry (September–November) (Childes & Walker Reference CHILDES and WALKER1987). The soil types are mainly lithosols and regosols developed on Karoo volcanic and Kalahari hydro-geological formations respectively (Chenje et al. Reference CHENJE, SOLA and PALECNZY1998).
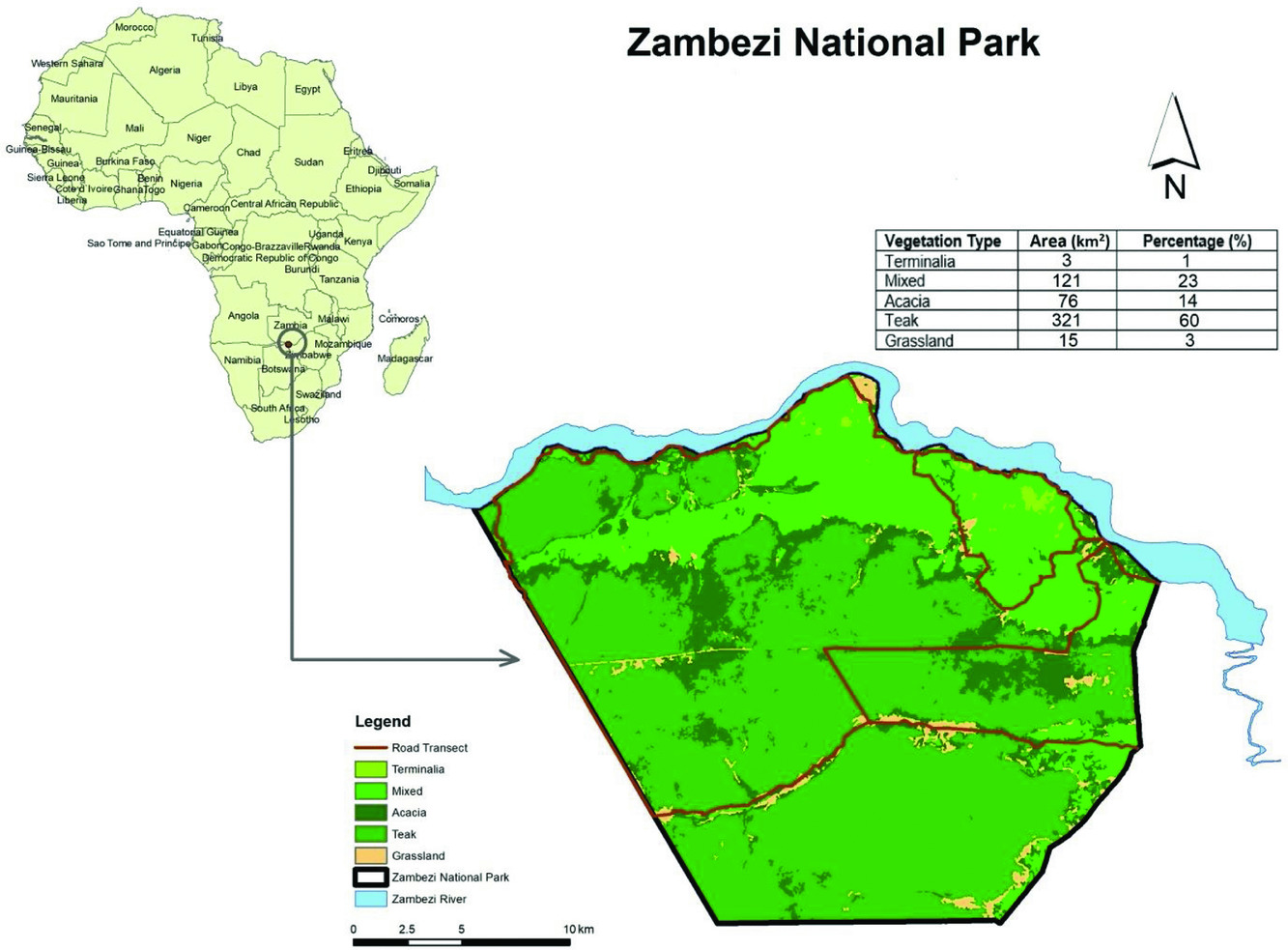
Vegetation in the ZNP consists of a mixture of forests, bushlands, woodlands and open grasslands and can be divided into five types viz. mixed, acacia, Zambezi teak (Baikiaea plurijuga), terminalia and grasslands (Table 1, Figure 1). The five vegetation types were determined using an unsupervised classification on a Landsat satellite image which was classified into five land cover classes using the K means classification algorithm. The image pixels were grouped into clusters based on their reflectance properties and these clusters were manually identified as different vegetation classes using the software ArcMap 10.4.1 (Esri 2011). The images were downloaded from USGS GloVIS website (http://glovis.usgs.gov/) based on scene ID 171r075 and for 16 March 2015, which was towards the end of the growing season. The image quality had 0% cloud cover and was orthorectified.
Table 1. Description of Mixed, Acacia, Zambezi teak, Terminalia and Grassland vegetation types in the Zambezi National Park, Zimbabwe


Figure 1. Map of Zambezi National Park showing the road transect (red line) and the five vegetation types.
Zambezi National Park supports a large and diverse herbivore population which include plains zebra Equus quagga Gray, greater kudu Tragelaphus strepsiceros Pallas, African buffalo Syncherus caffer Sparrman and the impala. The principal predators are lion Panthera leo, spotted hyaena Crocuta crocuta and leopard Panthera pardus.
Impala density
We determined impala densities using road transect surveys, a modification of the line transect survey method (Buckland et al. Reference BUCKLAND, ANDERSEN, BURNHAM, LAAKE, BORCHERS and THOMAS2001), monthly between January 2013 and December 2015. A road transect of 29.7 km which passes through five vegetation types (1.2 km acacia, 3.9 km mixed, 13 km grassland, 11 km Zambezi teak and 0.6 km terminalia) (Figure 1) was driven along established roads in the ZNP in the late afternoon between 15 h and 18 h (predominant feeding time of herbivores). These roads were established for maintenance and game viewing and are considered to provide a representative sample of the area. The road drive counts involved a driver driving at a speed of between 15–20 km h−1, with two experienced observers sitting at the back of a safari vehicle, identifying and counting animals. When study animals were sighted the vehicle was stopped, and the species and number of animals recorded. A global positioning system (GPS) location was recorded using a GPS (Garmin 12XL) and the direct distance to an individual animal or centre of a group, was measured with a laser rangefinder (Yardage Pro 1000) and angle was obtained with a compass.
Grass height
Grass height was measured in six randomly selected sampling sites in each vegetation type where herbivore counts were conducted in February 2015. We directly measured grass height by placing a ruler vertically from the ground next to the grass plants to the highest apical point.
Chemical composition of grass and woody species
Grass samples were collected in February 2015 (wet season) and August 2015 (dry season) in each of the five vegetation types. On each sampling occasion about 100 g of grass was collected at six sampling sites per vegetation type by clipping. Woody plant samples (~100 g) were collected in August 2015 from five browse species in acacia, terminalia and mixed vegetation types. Both grass and woody plant samples were air dried and then oven dried at 60°C for a minimum of 48 h to a constant weight and ground in a Wiley mill to pass through a 1 mm sieve before chemical analysis and in vitro gas production. Chemical analysis was conducted at the Department of Animal Science, University of Zimbabwe, Zimbabwe. Grass and woody plant nitrogen (N) was determined using the standard Kjeldahl technique (AOAC 2012) and phosphorus (P) by the phosphomolybdate method after dry ashing (Watanabe & Olsen Reference WATANABE and OLSEN1965). Calcium (Ca) concentrations were analysed with an Atomic Absorption Spectrophotometer. Acid detergent fibre (ADF) was determined following standard methods (AOAC 2012). In vitro gas production (IVGP) was determined following the procedure by Menke & Steingass (Reference MENKE and STEINGASS1988). Rumen fluid used was collected before the morning feeding from a rumen-fistulated steer and kept in a flask at 37°C. Rumen-fistulated steers used for rumen fluid collection were fed on a diet of grass collected from the sampling area and diets incorporating the five woody species.
Data analysis
Estimates of impala densities were conducted using the software Distance 6.0 release 2 (Thomas et al. Reference THOMAS, BUCKLAND, REXSTAD, LAAKE, STRINDBERG, HEDLEY, BISHOP, MARQUES and BURNHAM2010) for each vegetation type per season. For each season data for four months and three years (2013–2015) were pooled.
Grass chemical composition and grass height were compared among five vegetation types, while the chemical composition of five woody species was compared, all using one-way analysis of variance (ANOVA). Tukey's honest significant difference test was used, a posteriori, to compare pairs of means. The effect of season (wet and dry) (only for grass chemical composition) was tested using the independent-samples t-tests. All data were validated for ANOVA assumptions using the Kolmogorov–Smirnov test for normality and Levene's test for homogeneity of variances. All statistical analyses were conducted using IBM SPSS v.19. The significance level was set at alpha = 0.05.
RESULTS
Impala density
The impala used mixed, acacia and terminalia while avoiding Zambezi teak and grassland vegetation types (Table 2). The impala habitat use was consistent throughout the year, although densities declined from wet to dry season.
Table 2. Mean and range (in parentheses) of impala densities (animals km−2) in five vegetation types during the wet, cool and dry season in the Zambezi National Park, Zimbabwe.

*Observations too few to estimate confidence limits.
Chemical composition of grass and woody species
Grass N, P and IVGP declined, Ca showed no clear trend, while fibre (ADF) increased between wet and dry season (Table 3). Acacia nilotica was the most nutritive with high IVGP and N, low ADF and CT among the woody species (Table 4). Nitrogen and Ca were higher in woody plants than in grasses, IVGP and ADF were higher in grasses than woody plants, while P was comparable between grasses and woody plants (Table 3, 4).
Table 3. Grass chemical composition and in vitro gas production (ml per 300 mg DM-48h) in five vegetation types in the Zambezi National Park, Zimbabwe, in the wet (February) and dry (August) season. All values are per cent means (±SE). Means with different superscripts (a, b, c, d, e) within each season differ significantly (Tukey's post hoc test). ns, not significant; * < 0.05, ** < 0.01, *** < 0.001.

Table 4. Woody plant species chemical composition and in vitro gas production (ml per 300 mg DM-48h) during the dry season (August) in the Zambezi National Park, Zimbabwe. All values are per cent means (± SE). Means with different superscripts (a, b, c, d, e) within each variable differ significantly (Tukey's post hoc test). *** < 0.001.

Grass height
Grassland had the tallest grasses and acacia the shortest (F 4,25 = 127, P < 0.001, Figure 2).

Figure 2. Grass height (mean ± SE) in five vegetation types in Zambezi National Park, Zimbabwe. Means with common lower case letters are not significantly different.
DISCUSSION
Our study findings show that the impala is not highly selective as it used three out of the five vegetation types. It favoured mixed, acacia and terminalia vegetation types, which constituted 37% of the protected area. The most palatable browse viz. Combretum species, D. cinerea, Acacia species and Terminalia species together with the most nutritive grasses Cynodon dactylon, Panicum maximum and Urochloa mosambicensis were found in these vegetation types. For instance, A. nigrescens is a highly nutritious browse species that is rich in P and N (Shannon et al. Reference SHANNON, MACKEY and SLOTOW2013). In addition, the three vegetation types were open improving visibility and predator detection. Grassland may have been avoided due to the tall and unpalatable grass species such as Hyparrhenia hirta, while Zambezi teak vegetation type was avoided presumably because of the absence of palatable browse and grass species as well as the dense forests with low visibility. The impala prefers open habitats (Estes Reference ESTES1991, Fritz & Bourgarel Reference FRITZ, BOURGAREL, Kingdon and Hoffmann2013), avoiding dense woodlands (Bukombe et al. Reference BUKOMBE, KITTLE, SENZOTA, MDUMA, FRYXELL and SINCLAIR2017) and tall unburnt grasslands (Rduch Reference RDUCH2016), as an anti-predator strategy (Anderson et al. Reference ANDERSON, HOPCRAFT, EBY, RITCHIE, GRACE and OLFF2010).
The lower impala density in the dry season means that individuals were less dispersed than in the wet season as they probably converged on nutrient-rich patches away from the roads. Ungulates retreat to areas of plentiful forage during the dry season (Sinclair Reference SINCLAIR and Boulière1983), while forage resources are more uniformly distributed in the wet than dry season (Bukombe et al. Reference BUKOMBE, KITTLE, SENZOTA, MDUMA, FRYXELL and SINCLAIR2017).
Grass nutritive value in terms of N, P, IVGP and ADF declined from wet to dry season. Furthermore, during the dry season woody plant leaf nutritive quality was higher than grass in terms of N, Ca and ADF content. This was in support of our second hypothesis and is presumably the major driver of the switch from grazing to browsing. The impala switch from grazing to browsing as soon as grass quality declines (Smithers Reference SMITHERS1983), for example, it predominantly grazes during the wet season and then switches to browse in the dry season (Rduch Reference RDUCH2016).
The three vegetation types used by the impala had short to medium grass height. Foraging in vegetation types with short to medium grasses allows the impala to crop high quality forage to meet its nutritional requirements (Wilmshurst et al. Reference WILMSHURST, FRYXELL and BERGMAN2000), and is in accordance with its preference for habitats with nutritive forage (Bhola et al. Reference BHOLA, OGUTU, SAID, PIEPHO and OLFF2012) and good visibility (Sinclair et al. Reference SINCLAIR, MDUMA and BRASHARES2003).
Our findings have implications for wildlife management in heterogeneous protected areas as this study clearly shows that the impala use three vegetation types which constitute only 37% of the ZNP, avoiding the Zambezi teak vegetation type which makes up 60% of the protected area.
CONCLUSIONS
Our findings show that the impala use three vegetation types with good quality grass and browse, switch from grazing to browsing when grass quality declines to below that of browse. Furthermore, the impala forage in vegetation types with short to medium grass height presumably to maximize on intake of nutritive forage and as an anti-predator strategy.
ACKNOWLEDGEMENTS
We would like to thank Mr E. Chidziya the Director-General of the Zimbabwe Parks and Wildlife Management Authority (ZPWMA) for permission to carry out this study in a Parks and Wildlife Management Estate, while Mr Tinaapi H. Madiri the ZPWMA Ecologist and Mr E. Ngosi the Area Manager for Zambezi National Park are thanked for their logistical support for this research. We would also like to thank Mr F. Nleya for his invaluable assistance with fieldwork. We are grateful to the editor in chief and two referees for their helpful comments, which greatly improved earlier drafts of the paper.