



One theory for the coexistence of ecologically similar species in diverse communities is the presence of fine-scale mechanisms to alleviate competition for shared resources (Hutchinson Reference Hutchinson1959, MacArthur & Levins Reference MacArthur and Levins1967). Dietary niche partitioning is thought to be one of the most important of these mechanisms, and increasing evidence has shown that even similar diets of animals in the same feeding guild can diverge in subtle ways (Kartzinel et al. Reference Kartzinel, Chen, Coverdale, Erickson, Kress, Kuzmina, Rubenstein, Wang and Pringle2015). This could be due to active competition, whereby the fundamental niche of each species is reduced to a smaller realized niche in areas where the species co-occur. Alternatively, dietary partitioning could be due to the evolution of specialization, whereby each species evolves adaptations that allow it to better exploit a subset of the available dietary resources. If abundance and competition are high, resource partitioning may occur at multiple scales, including among genera (Giannini & Kalko Reference Giannini and Kalko2004, Lopez & Vaughan Reference Lopez and Vaughan2007), among species (Roughgarden Reference Roughgarden1976) and within species (Clark Reference Clark1980, Emmons Reference Emmons1980, Lewis et al. Reference Lewis, O’Connell, Lewis, Campagna and Hoelzel2006, Mott et al. Reference Mott, Herrod and Clarke2017, Pacala & Roughgarden Reference Pacala and Roughgarden1982).
This study focuses on mechanisms of inter- and intraspecific resource partitioning among three species of frugivorous phyllostomid bat in the genus Carollia (C. castanea, C. perspicillata and C. sowelli). These species are ideal for examining ecological and evolutionary mechanisms for niche partitioning because they co-occur throughout much of their range, yet have similar patterns of habitat and resource use (Fleming Reference Fleming1991, Lopez & Vaughan Reference Lopez and Vaughan2007). All three species are highly abundant and forage primarily on fruits of Piper – a hyper-diverse plant genus that is dominant in forest understoreys across much of the Neotropics (Jaramillo & Manos Reference Jaramillo and Manos2001). If competition among species of Carollia has resulted in the evolution of fine-scale specialization, such that certain bats are adapted to certain species of Piper, then differences in diet among or within bat species would likely be reflected in specific physiological or behavioural traits that impact the preferences or foraging efficiency of different bats on specific species of Piper. This could occur at the interspecific scale, with dietary partitioning among species, or at the intraspecific scale, with dietary partitioning based on sex, age class or reproductive status. Fewer studies have examined niche partitioning at this finer scale; however, dietary variation among individuals can reduce competition and promote species coexistence by reducing the number of individuals in a population that overlap in niche space (Cloyed & Eason Reference Cloyed and Eason2017).
The goal of this study was to examine evidence for fine-scale niche partitioning occurring at the inter- or intraspecific scale in Carollia–Piper interactions. We hypothesized that competition among bats feeding on the shared resource of fruits of Piper has led to the evolution of differential specialization, whereby certain species of Carollia or intraspecific groups (e.g. sexes, age classes, reproductive statuses) consume more and prefer certain species of Piper. We tested this hypothesis through dietary analyses and behavioural experiments examining fruit preference. First, we used dietary analysis based on faecal samples to test whether bat species, sex, age class, or reproductive status predicts variation in (1) consumption of fruits in the genus Piper versus fruits from other genera, (2) consumption of species of Piper in mid- to late-successional habitats versus species of Piper in early-successional habitats, and (3) the overall composition of different species of Piper in the diet. Second, we used data from behavioural experiments examining fruit preference to test whether bat preference for different species of Piper varies depending on bat species, sex, age class or reproductive status.
Methods
Study site and system
All field experiments were conducted at La Selva Biological Station (LSBS), Heredia Province, Costa Rica during April–July 2012 and June–July 2013. The station is managed by the Organization for Tropical Studies (OTS) and comprises ~1600 ha of tropical wet forest. The site has a high diversity of Piper with over 60 species present and three species of Carollia co-occurring. The three species of Carollia (C. perspicillata, C. sowelli and C. castanea) occur in similar abundances at LSBS, and all three are among the most abundant bat species at the site (Fleming Reference Fleming1991). Species in the genus Carollia have relatively specialized diets compared with other South American frugivorous bat species (Banack Reference Banack1998), with infructescences (hereafter referred to as fruits) of Piper contributing at least 45–47% of the diet (Fleming Reference Fleming1991, Lopez & Vaughan Reference Lopez and Vaughan2007). However, the three species of Carollia at our site may differ in their relative specialization on Piper; past studies have found C. castanea is the most specialized, C. sowelli is intermediate, and C. perspicillata has the most varied diet (Fleming Reference Fleming1991, Thies & Kalko Reference Thies and Kalko2004, York & Billings Reference York and Billings2009, York & Papes Reference York and Papes2007). Furthermore, past work suggests that species of Carollia may differ in their relative reliance on different species of Piper (Lopez & Vaughan Reference Lopez and Vaughan2007). Each species of Piper differs in fruit presentation, size and chemistry (Hieu et al. Reference Hieu, Thang, Hoi and Ogunwande2014, Kato & Furlan Reference Kato and Furlan2007, Whitehead et al. Reference Whitehead, Jeffrey, Leonard, Dodson, Dyer and Bowers2013, Whitehead unpubl. data). Thus, there are several mechanisms through which different species, sexes, age classes or reproductive statuses of Carollia could discriminate among different species of Piper and evolve behavioural preferences that allow them to differentially specialize on particular species of Piper.
Diet composition
To test whether the overall dietary composition differed among and within three species of Carollia, we used data from faecal samples collected from 210 wild-captured individuals from April–July 2012 (55 C. castanea, 54 C. perspicillata, 101 C. sowelli). Bats were captured at LSBS using mist-nets (Kunz et al. Reference Kunz, Hodgkison, Weise, Kunz and Parsons2009), which were set up in various secondary-forest locations and moved to new areas on a weekly rotation. Upon capture, basic measurements were recorded for each bat, including sex, age class, weight, reproductive status and forearm and tibia lengths. Bats were then placed in cloth holding bags for 10–45 min (prior to their use in other experiments; Whitehead et al. Reference Whitehead, Obando-Quesada and Bowers2016), and faecal samples were collected opportunistically from the cloth holding bags when bats defecated during this time period. Faecal samples were collected in 2-mL microcentrifuge tubes and stored in ethanol prior to seed identification.
Seed identification to species was accomplished for ~96% of the seeds of Piper in faecal samples through seed comparisons. A seed reference collection was assembled from ripe fruits taken directly from plants and identified by Susan Whitehead, Marisol Luna and Orlando Vargas. All faecal samples were examined under a dissecting scope, and counts of the total numbers of seeds from each species of Piper were recorded. In cases where faecal samples contained greater than 1000 seeds, we sorted all seeds by species and estimated the total counts of each by counting subsamples in a grid and extrapolating to the estimated total. Seeds of other genera and rare species of Piper (4% unidentified) for which we did not have matches in our reference collection were identified to morphospecies.
Preference trials
To test whether different species of Carollia have different relative preferences for different species of Piper, preference trials were conducted in flight cages with 123 individuals of Carollia in April–May 2012 and June–July 2013 (Table 1). Bats were captured in mist-nets, as above, and all males and non-reproductive females of C. castanea, C. sowelli and C. perspicillata were retained for trials. Reproductive females, noted by either pregnancy or lactation, were released immediately and not used in preference trials (although we did collect faecal samples from reproductive females and included them in the dietary analysis, described above). The participating bats were kept in cloth bags and transported to flight cages (2 × 2 × 1 m) located within the station. Four flight cages were used, and bats were housed individually in all experiments. Following the experiments, all animals were marked temporarily by clipping fur to avoid re-use in the same experiment and were returned to the site of capture on the same night.
Table 1. Summary of bats used in preference trials, captured April–May 2012 and June–July 2013 at La Selva Biological Station in Costa Rica

Individual bats were released in flight cages where fruit of two species of Piper were presented near the end of the cage. The peduncle of each fruit was placed in a hole drilled at the end of a wooden dowel to mimic the upright presentation of ripe fruit of Piper used in this experiment. Each pairwise trial compared the preference of a single bat for P. sancti-felicis versus one of six other species: P. peracuminatum, P. peltatum, P. reticulatum, P. silvivagum, P. umbricola or P. colonense. All other species were compared with P. sancti-felicis as a baseline because it is a highly abundant species at the site, it fruits year-round and, according to results of the dietary analysis (Figure 1), represents ~50% of the diet of all three species of Carollia. Thus, a choice between a fruit of P. sancti-felicis or another species of Piper is a typical choice faced nightly by a foraging bat at LSBS. Most bats were used in only one choice trial, but 28 individuals (~15%) were used in multiple trials (up to five trials) comparing P. sancti-felicis to several different species of Piper, and our statistical analysis accounted for the non-independence of multiple trials conducted with the same bat. During the experiment, flight cages were monitored using Sony Nightshot video cameras equipped with infrared lights to avoid disturbing the bats’ behaviour. Videos were later reviewed, and the first fruit removed by each bat was recorded as its feeding preference. Individuals that did not feed after 1 h were recorded as ‘no-choice’. The feeding preferences of 6–14 individual bats were tested for each combination of species of Carollia and Piper in this manner. This excludes C. castanea and P. umbricola, for which only one trial was conducted due to limited availability of ripe fruits and limited C. castanea captures.

Figure 1. Proportion of unique plant species occurrences from faecal samples collected from three species of Carollia at La Selva Biological Station in Costa Rica. Seeds were categorized as one of the five most common species of Piper, one of eight uncommon species of Piper, or other genera. In total, we recorded 263 unique occurrences (i.e. multiple seeds from the same species in the same faecal sample were only counted once) in 210 total faecal samples. Uncommon species of Piper included P. peracuminatum (7 occurrences), P. peltatum (2), P. holdridgeanum (1), P. auritum (1), P. evasum (1), P. cyanophyllum (1), P. nudifolium (1) and P. urostachyum (1).
Fruits used in the experiments were harvested each day from plants growing around the research station. Ripe fruits were gathered by shaking the desired plant, which causes the ripened fruits to fall, and were distinguished from unripe fruits by their soft texture and smell. Only fruits that had ripened on the day of the experiment were used due to the fruits’ rapid decomposition. If not consumed on the first night they are ripe, fruits begin to rot within 24 h (Thies & Kalko Reference Thies and Kalko2004, Whitehead & Bowers Reference Whitehead and Bowers2014).
Statistical methods
To assess the completeness of our sampling of bat diets, we extrapolated species richness for Piper using rarefaction based on the function specpool in R package ‘vegan’ (Samarah Reference Samarah2017). To explore how the relative reliance on fruits of Piper may vary among and within bat species, we first created a binary variable from the faecal-sample data depending on whether the sample contained exclusively fruits of Piper (1) or other genera (either alone or mixed with Piper) (0). We fitted the data to a generalized linear mixed-effects model (GLMM) with a binomial distribution using the glmer function in the ‘lme4’ package in R (Bates et al. Reference Bates, Mächler, Bolker and Walker2015). Fixed effects included bat species, age class, sex and reproductive status. Date was included as a random effect. To test the significance of each fixed effect as a predictor, we used a model simplification approach (Crawley Reference Crawley and Crawley2007), comparing complex models to simplified versions using likelihood ratio tests. No interactions were included in the models due to limited sample sizes. After finding a significant effect of species, a post hoc Tukey’s HSD test was used to examine differences in diet among the three species.
To test whether the three bat species vary in diet composition of species of Piper from different successional stages, we first grouped the 13 species of Piper into two categories: (1) early-successional species, and (2) mid- and late-successional species (Table 2). Due to the highly variable number of seeds found in the fruits of different species of Piper, seed count data from faecal samples were transformed into presence/absence data. Counts of unique occurrences of species of Piper found in early-successional habitats versus mid- to late-successional in each faecal sample were then used as a binomial response variable. Data were fitted to a binomial GLMM with bat species, sex, age class and reproductive status as fixed effects and date as a random effect. No interactions were included in these models due to limited sample sizes for assessing higher-level interactions among these variables. Variable significance was tested using a model simplification approach as above.
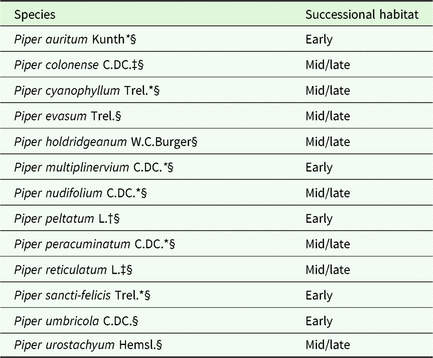
Table 2. Species of Piper detected in Carollia diet and grouped by successional stage. Successional stage classification is based on *Fleming (Reference Fleming, Dyer and Palmer2004); †Jaramillo & Callejas (Reference Jaramillo, Callejas, Dyer and Palmer2004); ‡Thies & Kalko (Reference Thies and Kalko2004); §Personal observations
To test for overall differences in the dietary abundances of different species of Piper among bat diets, we performed a non-metric multidimensional scaling (NMDS) analysis (Minchin Reference Minchin1987) based on the relative abundances of each species of Piper in each faecal sample for individual bat. We then examined whether the data were partitioned based on species, sex, age class and reproductive status. Prior to the analysis, we created a simplified dataset: unidentified species were grouped and uncommon species of Piper (species found in less than 10 faecal samples) were grouped. Abundance for the five most common species of Piper were included individually. The ordination was based on the Bray–Curtis dissimilarity index and used 200 replicates with random starting coordinates. A three-dimensional solution was chosen based on a scree plot, which plots the decrease in stress based on increases in dimension (Clarke Reference Clarke1993). This analysis was conducted using the ‘vegan’ package in R. To estimate food niche overlap among the three bat species and within species based on age class, sex and reproductive status, we calculated Pianka’s symmetric equation (Pianka Reference Pianka1973) using the pianka function in the ‘EcoSimR’ package in R. This niche overlap index varies between 0 (no overlap) and 1 (complete overlap).
Lastly, to test whether there is variation among and within bat species in their preference for different species of Piper, we used data collected from the preference trials. The data were fitted to a binomial GLMM using binary fruit choice (P. sancti-felicis = 0, other species of Piper = 1) as the response variable. We began with a global model that included fixed effects of the species of Carollia, the species of Piper and their interaction, as well as bat sex, age class and reproductive status. The individual bat identifier and date were included as random effects. A model simplification approach was used as above to test the significance of each fixed-effect term. After finding a significant interaction between the species of Piper and species of Carollia, the data were analysed separately for each species of Piper, using only bat species as a predictor variable. Differences in preference among bat species were tested using model simplification, as above. Where significant differences were found among bat species, a Tukey post hoc test was conducted to determine where the differences occurred among the three species of Carollia. In addition to examining differences among bat species, for each type of Piper trial we also examined whether there were overall preferences for P. sancti-felicis or the other species of Piper. This was examined across all bat species using an intercept-only binomial generalized linear model (GLM) predicting fruit choice. In these models, the parameter estimate for the intercept can be interpreted to determine whether the log-odds ratio is different than 1:1 (i.e. random choice) for that type of trial.
Data availability
Data from this study are archived at the Dryad Digital Repository (doi: 10.5061/dryad.nj2b2h4).
Results
Across all 210 faecal samples, we found seeds from 13 species of Piper and 15 species from other genera (Figure 1). Insect remains were also detected in two samples from C. castanea, eight samples from C. sowelli and seven samples from C. perspicillata; however, these results were not included in further analyses due to low sample sizes. A species richness estimation suggested that our sampling captured 85% of the total Piper diversity in three bat species’ diets during our sampling period. There were five dominant species of Piper in bat diets: P. sancti-felicis, P. colonense, P. umbricola, P. multiplinervum and P. reticulatum (Figure 1). Across the three species of Carollia, 83.8% of faecal samples contained one or more of these species. Most samples (88.1%) contained only species of Piper; 7.1% of samples contained seeds from both Piper and other genera, and 4.8% contained only seeds from other genera.
In our analysis of the likelihood that samples would be exclusively species of Piper versus mixed genera, we found an overall effect of bat species on dietary composition (χ2 = 10.7, P = 0.005). All three bat species consumed primarily species of Piper (Figure 1), but a larger percentage of samples from C. castanea contained exclusively species of Piper compared with samples from C. sowelli (P = 0.04) or C. perspicillata (though not significantly so at P = 0.06) (Figure 2a). Sex also had an effect on dietary composition (χ2 = 4.72, P = 0.03), with a slightly larger percentage of faecal samples from females containing exclusively species of Piper (Figure 2b). There was a slightly larger percentage of non-reproductive bats (91.4%) consuming exclusively species of Piper compared to reproductive bats (84.0%), but the pattern was not significant (χ2 = 3.06, P = 0.08). Age class was not a significant predictor of dietary composition (χ2 = 0.006, P = 0.94).

Figure 2. Proportion of faecal samples collected from three species of Carollia at La Selva Biological Station in Costa Rica that contained exclusively species of Piper vs mixed genera (with or without Piper) varied depending on bat species (a; P = 0.005) and bat sex (b; P = 0.03).
In our analysis of the dietary composition of species of Piper from different successional stages, there were no significant differences among bat species in their relative reliance on mid- or late-successional species versus early-successional species of Piper (P = 0.82). Additionally, we found no effects of bat sex (P = 0.87) or reproductive status (P = 0.76). However, there were significant differences based on bat age class (χ2 = 4.82, P = 0.04), with adult bats being more likely than juveniles to consume early-successional species of Piper (Figure 3).

Figure 3. Proportion of unique occurrences of seeds of different species of Piper from faecal samples at La Selva Biological Station in Costa Rica. Seeds in samples were identified as early-successional or mid- to late-successional species varied across bat age class. In total, we recorded 263 unique occurrences of Piper (i.e. multiple seeds from the same species in the same faecal sample were only counted once) in 210 total faecal samples. Mid- to late-successional species of Piper were more common in samples from juvenile bats than adult bats (P = 0.04).
The NMDS analysis showed no impact of bat species, sex, age class or reproductive status on Piper dietary composition of individual bats (R 2 = 0.81, Final Stress = 0.002, Figure 4). Results from the Pianka sympatric indices calculated for each pairwise comparison among species of Carollia indicated almost complete overlap in dietary niche: C. castanea versus C. perspicillata = 0.98; C. castanea versus C. sowelli = 0.99; C. perspicillata versus C. sowelli = 0.98. The index also indicated near-complete overlap between sexes (0.99), age classes (0.98) and reproductive statuses (0.97).

Figure 4. NMDS ordination plot on the relative abundances of seeds of different species of Piper in faecal samples based on bat species (a), sex (b), age class (c), and reproductive status (d).
In the behavioural trials examining relative preferences of three species of Carollia for various species of Piper, there was an interaction between species of Carollia and species of Piper in determining fruit preference (χ2 = 21.4, P = 0.02) (Figure 5). All bat species exhibited a non-random aversion toward P. silvivagum and P. peltatum. There was a significant difference among bat species in preference for P. colonense (P = 0.04), with C. sowelli preferring P. colonense (90% of trials). Juvenile bats chose P. santi-felicis more often (74.1% of trials) than adult bats (54.5%), but the choice was not significant (χ2 = 2.75, P = 0.097). Bat sex (χ2 = 0.022, P = 0.88) and reproductive status (χ2 = 0.0024, P = 0.96) were non-significant predictors of fruit preference.

Figure 5. Results from binary choice experiments comparing preference among the three species of Carollia for P. sancti-felicis versus six other common species of Piper. Bars extending to the left from the middle line indicate a choice for P. sancti-felicis. Bars extending to the right from the middle line indicate a choice for the other species of Piper. The total lengths of the bars represent the total number of choices made by all bat species, with the proportion of those choices attributed to each species colour-coded within the bar. All bat species exhibited a non-random aversion toward P. silvivagum and P. peltatum. There was a significant difference among bat species in preference for P. colonense (P = 0.04), with a higher preference for P. colonense exhibited by C. sowelli.
Based on the interaction between species of Carollia and species of Piper, we also analysed the data separately for each combination of species of Piper. Bats exhibited non-random choice in only two sets of combinations: P. sancti-felicis with P. peltatum and P. sancti-felicis with P. silvivagum. In both cases, bats preferred P. sancti-felicis in a significant departure from random expectation. In one combination, we found limited evidence that preferences may differ among species of Carollia. For P. colonense, there was an overall difference in preference among bat species (χ2 = 6.66, P = 0.04). Carollia sowelli had the highest preference for P. colonense, but post hoc tests for all pairwise comparisons among the three bat species showed no significant differences (C. sowelli vs C. castanea, P = 0.09; C. sowelli vs C. perspicillata, P = 0.16, C. castanea vs C. perspicillata, P = 0.85; Figure 5). There were no differences in preference among bat species for any other combinations of species of Piper (Figure 5).
Discussion
Classic theory of coexistence suggests that, to avoid competitive exclusion, organisms must partition shared resources in some way (McNab Reference McNab1971, Tamsitt Reference Tamsitt1967). This could occur through ongoing competition (reducing the realized niche of organisms on ecological time scales) or through the evolution of specialization (reducing the niche breadth of species on evolutionary time scales). This study paired field observations of diet in three co-occurring species of Carollia with controlled preference experiments to explore the evidence for fine-scale dietary niche partitioning and the evolution of specialization in this genus. Our findings provide some evidence for inter- and intraspecific dietary differences in consumption of species of Piper depending on the species, sex and age class of Carollia. However, in general, there was strong overlap in both diet and fruit preference among and within the three species of Carollia, suggesting other axes of niche differentiation may be more important than diet in permitting the coexistence of these species.
Across the three bat species, our analyses of dietary records supported previous work that found C. castanea to rely more exclusively on species of Piper compared with the other two species of Carollia (Fleming Reference Fleming1991, Giannini & Kalko Reference Giannini and Kalko2004). However, our results found that all three species of Carollia consume primarily species of Piper, with <10% of dietary records attributed to items outside of the genus Piper (Figure 1). In comparison, past work at the same site has suggested that up to 50% of the diet of C. perspicillata does not include species of Piper (Fleming Reference Fleming1991). It is possible that our study design overestimated the proportion in species of Piper in bat diets due to certain limitations in our sampling, and we discuss these possible caveats in detail below. However, it is also possible that the reliance of species in the genus Carollia on fruits in the genus Piper could be increasing over time, perhaps due in part to the fact that secondary forest sites are increasingly dominant in tropical landscapes (Brown & Lugo Reference Brown and Lugo1990, Phillips et al. Reference Phillips, Newbold and Purvis2017), and include an abundance of species of Piper (Fleming Reference Fleming, Dyer and Palmer2004). Considering the strong reliance on fruits of Piper for all three bat species (Figure 1), combined with the strong overlap among species in the consumption of species of Piper (Figure 4), competition for food resources among these bats is likely high. Even where fruits of Piper are abundant, almost all (~90%) of ripe fruits are removed the first night they become ripe (Bonaccorso et al. Reference Bonaccorso, Winkelmann, Shin, Agrawal, Aslami, Bonney, Hsu, Jekielek, Knox, Kopach, Jennings, Lasky, Menesale, Richards, Rutland, Sessa, Zhaurova and Kunz2007, Fleming Reference Fleming, Dyer and Palmer2004, Thies & Kalko Reference Thies and Kalko2004, pers. field obs.), suggesting that it is not only a shared resource but a limited one.
We hypothesized that the three species of Carollia might alleviate competition through differences in their relative specialization and preference for different species of Piper. We expected that, due to a long history of overlapping ranges, each species would have evolved specialized mechanisms that increase their foraging efficiency on certain species of Piper. However, the results from our dietary analysis indicate no clear differences among species of Carollia in the relative reliance on different species of Piper (Figure 1) – all three rely heavily on some of the most abundant species reported at the site (Fleming Reference Fleming, Dyer and Palmer2004). In contrast to previous work (Fleming Reference Fleming1991, Thies & Kalko Reference Thies and Kalko2004), our findings did not indicate species-level partitioning among three species of Carollia for early-successional versus mid- to late-successional species of Piper. Furthermore, the preference trials indicated strong overlap in behavioural patterns of resources use. We found that all bats had an aversion toward P. peltatum and P. silvivagum. These findings are in accordance with our dietary records, which included no occurrences of P. silvivagum and only two occurrences of P. peltatum. Although P. silvivagum is somewhat rare and, in our experience, restricted to clearings, P. peltatum is fairly widespread and fruits in abundance. The clear aversion toward P. peltatum by bats, based on both diet records and preference trials, suggests this plant relies primarily on other dispersers, such as ants (Whitehead, pers. field obs.). The one case in which we found statistical differences in preference among bat species was for P. colonense (Figure 5), where C. sowelli exhibited a stronger preference than C. castanea for P. colonense. Although these differences were not reflected in the dietary abundance of P. colonense (Figure 1), the specific physiological mechanisms that underlie the observed variance in preference (e.g. differences in the sensitivity of bats to different olfactory cues produced by the two species) could be explored in future studies.
The potential for resource partitioning within species of Carollia has received less attention in past studies, and our results suggest that this may be an important mechanism through which competition is reduced. Our results show that female bats rely more heavily on fruits of Piper, with 92% of faecal samples containing exclusively fruits of Piper. Our sampling was conducted during the breeding season, and past work has shown that reproductive females eat as much as non-reproductive females and males but harvest fruits in much shorter flights with less time spent exploring (Charles-Dominique Reference Charles-Dominique1991). Our results suggest that reproductive females may focus primarily on locating and removing fruits of Piper, whereas males and non-reproductive females will forage for other fruits or insects during foraging flights. Although our sample size was limited, we did find a few more insect remains in faeces from male bats compared with females (seven samples from females and 10 samples from males). Our results also revealed that juvenile bats of all three species consumed significantly more species of mid- to late-successional species of Piper compared with adult bats. This may indicate that younger bats use less-open flyways, foraging in denser areas with more cover. These findings could also align with the work of Charles-Dominique (Reference Charles-Dominique1991), which showed bachelor males (usually subadults) foraged more widely than harem males (adult males protecting a territory). Interestingly, the intraspecific differences observed in the diets of different bat sexes and age classes based on faecal samples (Figures 2–4) were not reflected in the preference experiments (Figure 5). Thus, the intraspecific differences in the relative reliance on exclusively species of Piper and consumption of species of Piper from different successional habitats are probably products of temporal and spatial specialization in habitat use as opposed to preference.
Several limitations of our dataset should be considered when interpreting our results and placing the results in the context of other research. First, all bats were captured in the first half of the night between 18h00 and 00h00 (with the majority of captures between 18h00 and 21h00). Considering our captures were in the beginning of all feeding bouts, it is possible that our faecal data indicated larger amounts of fruits of Piper in the diet than would be estimated based on sampling throughout the night. However, Thies et al. (Reference Thies, Kalko and Schnitzler2006) found that foraging time of bats in the genus Carollia will plateau after the first 3 h after sunset (21h00) because most fruits of Piper have been harvested. Bonaccorso et al. (Reference Bonaccorso, Winkelmann, Shin, Agrawal, Aslami, Bonney, Hsu, Jekielek, Knox, Kopach, Jennings, Lasky, Menesale, Richards, Rutland, Sessa, Zhaurova and Kunz2007) found that C. perspicillata avoids predation by beginning feeding later than C. castanea; thus, C. perspicillata has limited access to ripe fruits of Piper and may switch to other food sources. Second, faecal analysis, which was used in this study, is inherently biased against detecting dietary remnants that do not possess a hard exoskeleton or seed coat (Herrera et al. Reference Herrera, Gutierrez, Hobson, Altube, Diaz and Sanchez-Cordero2002). We found few samples with insect remains (two samples from C. castanea, eight samples from C. sowelli and seven samples from C. perspicillata), but these data do support past studies showing trophic stratification (York & Billings Reference York and Billings2009). Third, our study included reproductive females in the dietary analysis but excluded them from preference trials. Fourth, our analyses did not separate sexes, age classes or reproductive states by species due to limited sample sizes. Lastly, our studies were conducted during the rainy season at LSBS. Future work should incorporate both temporal seasons to test for seasonal differences in feeding preferences.
Species of Carollia have similar habitat use, foraging habits and reproductive timing and all depend on the fruits of Piper year-round as their primary food source (Fleming Reference Fleming1991). Here we show that dietary partitioning within species of Carollia based on sex and age class may be an important, but overlooked, mechanism through which competition among bats may be reduced. Furthermore, in contrast to past studies, we found little evidence of species-level dietary partitioning among the three species of Carollia. In general, our results show much stronger dietary overlap among these bat species and a stronger overall reliance on fruits of Piper than has been reported in previous studies that were conducted decades ago. Future research should continue to explore whether resource-use patterns in these highly abundant and widespread species could be changing over time – trending towards increasing competition among co-occurring bat species for resources. This has important implications for the persistence of species and interaction diversity. Continued research on the Piper–Carollia interaction provides a valuable case study for understanding specialization in fruit–frugivore interactions and the mechanisms underlying biodiversity in tropical forests.
Author ORCIDs
Lauren D. Maynard https://orcid.org/0000-0003-2059-1250, Susan R. Whitehead https://orcid.org/0000-0002-7089-4594
Acknowledgements
We thank Orlando Vargas Ramírez and Marisol Luna Martínez for their help with Piper identification and seed collections at La Selva Biological Station (Organization for Tropical Studies, OTS). We are also very grateful for the help of Bernal Matarrita Carranza and Danilo Brenes Madrigal for their positive presence and assistance throughout the project. Daniela Azofeifa assisted with field data collection. Deane Bowers provided helpful suggestions on the experimental design and use of laboratory facilities for seed identifications at the University of Colorado. York Winter kindly allowed us access to his bat cages.
Financial support
Funding for field data collection for this study was provided by a National Science Foundation Doctoral Dissertation Improvement Grant (DEB 1210884 to SRW), a National Science Foundation Research Experience for Undergraduates Grant (DBI 0851933 to the Organization for Tropical Studies), a National Geographic Waitt grant to SRW, and the Department of Ecology and Evolutionary Biology at the University of Colorado. Support for LDM during analyses and writing of the manuscript was provided through start-up funds to SRW from Virginia Polytechnic Institute and State University.







