INTRODUCTION
The Cossuridae is a small family of polychaetes that are easily recognized by a single branchial filament arising mid-dorsally from one of the anterior-most segments. The family currently contains only a single genus with 26 valid species (WoRMS Editorial Board 2016). A single cossurid species, Cossura coasta Kitamori, Reference Kitamori1960, has been recorded from West African waters (Day, Reference Day1967; Lange et al., Reference Lange, Darr and Zettler2014). This paper describes two cossurid species from the terminal lobes of the Congo deep-sea fan. This region covers an area of 2500 km2 at a depth of 4700–5100 m and an offshore distance of 750–800 km. It is a unique alluvial area in the world ocean fed by a submarine canyon and a channel-levee system that presently deposits large amounts of organic carbon originating from the Congo River by turbidity currents (Rabouille et al., Reference Rabouille, Olu, Baudin, Khripounoff, Dennielou, Arnaud, Babonneau, Bayle, Beckler, Bessette, Bombled, Bourgeois, Brandily, Caprais, Cathalot, Charlier, Corvaisier, Croguennec, Cruaud, Decker, Droz, Gayet, Godfroy, Hourdez, Le Bruchec, Le Saout, Lesaout, Lesongeur, Martinez, Mejanelle, Michalopoulos, Mouchel, Noel, Pastor, Picot, Pignet, Pozzato, Pruski, Rabiller, Raimonet, Ragueneau, Reyss, Rodier, Ruesch, Ruffine, Savignac, Senyarich, Schnyder, Sen, Stetten, Sun, Taillefert, Teixeira, Tisnerat-Laborde, Toffin, Tourolle, Toussaint, Vetion, Jouanneau and Bez2016).
MATERIALS AND METHODS
The Congo lobe project was built upon two sampling efforts during two oceanographic cruises on board the RV ‘Pourquoi Pas?’ equipped with the ROV ‘Victor 6000’: WACS (West Africa cold seeps) from 27.1.2011 to 28.2.2011 and CONGOLOBE from 12.12.2011 to 10.1.2012. For a detailed description of sampling sites and collecting methods, see the work of Rabouille et al. (Reference Rabouille, Olu, Baudin, Khripounoff, Dennielou, Arnaud, Babonneau, Bayle, Beckler, Bessette, Bombled, Bourgeois, Brandily, Caprais, Cathalot, Charlier, Corvaisier, Croguennec, Cruaud, Decker, Droz, Gayet, Godfroy, Hourdez, Le Bruchec, Le Saout, Lesaout, Lesongeur, Martinez, Mejanelle, Michalopoulos, Mouchel, Noel, Pastor, Picot, Pignet, Pozzato, Pruski, Rabiller, Raimonet, Ragueneau, Reyss, Rodier, Ruesch, Ruffine, Savignac, Senyarich, Schnyder, Sen, Stetten, Sun, Taillefert, Teixeira, Tisnerat-Laborde, Toffin, Tourolle, Toussaint, Vetion, Jouanneau and Bez2016). Sediment samples were taken in layers up to 25 cm deep and washed through sieves with 250, 300, 500 and 1000 µm mesh sizes. Animals were preserved in 4% formaldehyde solution in seawater and transferred to 80% ethanol. Specimens were examined with a stereomicroscope, and staining with methylene blue was used to enhance the contrast of small details. Some of the specimens were temporarily mounted in glycerol and studied with a compound microscope. Stacks of images were combined using Helicon Focus software to achieve completely focused images. A few specimens were critical-point dried, coated and examined using a Camscan S2 scanning electron microscope (SEM). The type specimens are deposited in the National Museum of Natural History, Paris (MNHN).
RESULTS
SYSTEMATICS
Family COSSURIDAE Day, 1963
Genus Cossura Webster & Benedict, Reference Webster and Benedict1887
Cossura platypus sp. nov.
(Figures 1 & 2)

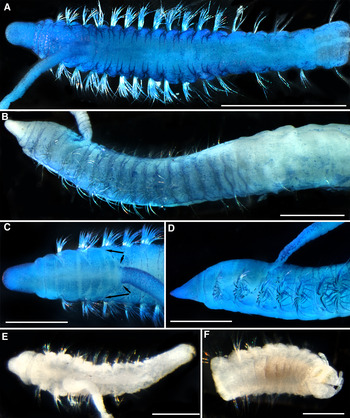
Fig. 1. Cossura platypus sp. nov., (A–D) methylene blue staining, (A) holotype, (MNHN-IA-TYPE 1779), dorsal view; (B) paratype, (MNHN-IA-TYPE 1781), lateral view; (C) paratype, (MNHN-IA-TYPE 1780), anterior end, dorsal view; arrows mark epithelial fold extending from posterior border of chaetiger 2 to base of branchial filament; (D) paratype, (MNHN-IA-TYPE 1780), anterior end, lateral view; (E) paratype, (MNHN-IA-TYPE 1782), juvenile, dorsal view; (F) paratype, (MNHN-IA-TYPE 1783), posterior end with pygidium. Scale bars: A, 1 mm; B, 500 µm, C, D, 300 µm, E, F, 200 µm.

Fig. 2. Cossura platypus sp. nov., SEM, (A) paratype, (MNHN-IA-TYPE 1780), dorsal view; (B) same specimen, anterior end, dorsal view; (C) paratype, (MNHN-IA-TYPE 1780), ventral view; (D) paratype, (MNHN-IA-TYPE 1780), anterior end, lateral view; (E) same specimen, parapodia of chaetigers 2–4; (F) paratype, (MNHN-IA-TYPE 1780), anterior end, lateral view; (G) paratype, (MNHN-IA-TYPE 1780), abdominal parapodia, anterior view. Scale bars: A, 300 µm; B–D, F, 100 µm, E, G, 30 µm.
TYPE MATERIAL
Holotype: (MNHN-IA-TYPE 1779). RV ‘Pourquoi Pas?’, WACS cruise, ROV ‘Victor 6000’, dive 425, 4.2.2011, 10:39:18, large blade cores, GCL1, layer 0–5 cm, fraction 500 µm, location REGAB, 05°47.984′S 009°42.475′E, depth 3154 m.
Paratypes: (MNHN-IA-TYPE 1780), 19 specimens, RV ‘Pourquoi Pas?’, WACS cruise, ROV ‘Victor 6000’, dive 425, 4.2.2011, 10:39:18, large blade cores, GCL1, layer 0–5 cm, fraction 500 µm, location REGAB, 05°47.984′S 009°42.475′E, depth 3154 m; (MNHN-IA-TYPE 1781), 21 specimens, RV ‘Pourquoi Pas?’, WACS cruise, ROV ‘Victor 6000’, dive 427, 8.2.2011, 11:00:06, large blade cores, GCL4, layer 0–5 cm, fraction 1000 µm, location REGAB, 05°47.866′S 009°42.685′E, depth 3156 m; (MNHN-IA-TYPE 1782), 9 specimens, RV ‘Pourquoi Pas?’, WACS cruise, ROV ‘Victor 6000’, dive 427, 9.2.2011, 02:35:00, large blade cores, GCL7, layer 1–3 cm, fraction 250 µm, location REGAB, 05°48.761′S 009°42.360′E, depth 3285 m; (MNHN-IA-TYPE 1783), 1 specimen, RV ‘Pourquoi Pas?’, WACS cruise, ROV ‘Victor 6000’, dive 427, 9.2.2011, 02:35:00, large blade cores, GCL7, layer 5–10 cm, fraction 300 µm, location REGAB, 05°48.761′S 009°42.360′E, depth 3285 m.
ETYMOLOGY
The name refers to the shape of the prostomium, which in dorsal view resembles a platypus beak.
DESCRIPTION
Colour in alcohol uniform white or pale yellowish, chaetae light golden. Holotype with 17 chaetigers, 15 thoracic, 400 µm wide (Figure 1A). All specimens incomplete, most represented by short fragments of 16–19 segments, one posterior fragment with pygidium. Thoracic width up to 450 µm; 14–17 thoracic chaetigers with or without sharp border between body regions; 12–14 anterior chaetigers swollen, with lateral and dorsal inflations and wide intersegmental furrows; dorsal groove located mid-dorsally (Figures 1A & 2A). Segments becoming longer in the abdominal region (Figure 1A, B).
Prostomium triangular in lateral view, longer than it is wide, with widely rounded anterior margin, lateral margins almost parallel or converging at small angle (Figures 1A, C, D & 2A–D, F). Prostomial furrow more or less developed depending on muscle contraction. Posterior prostomial ring abruptly expanded, same length as peristomium, without mid-ventral notch (Figures 1C & 2A, B). One specimen with everted proboscis with 10 buccal tentacles. Nuchal organs not observed.
Branchial filament attached to midlength of chaetiger 3 (Figures 1B–D & 2A, B, D). In stained specimens, a triangular fold of epithelium extending from posterior border of chaetiger 2 to base of filament (Figure 1C).
Chaetiger 1 with uniramous parapodia, all subsequent segments bearing biramous parapodia; parapodial rami situated very close to each other (Figures 1B, D & 2D–F). All chaetae hirsute capillaries with smooth shafts, arranged in two indistinct rows. Noto- and neurochaetae in thorax of the same width, chaetae in anterior row are wider (shaft 6 µm, hirsute part 10 µm thick) and shorter than chaetae in posterior row (shaft 4 µm, hirsute part 6 µm thick) (Figures 1D & 2D–F). First chaetiger bearing 10–11 chaetae; in subsequent segments notopodia bearing 6–9 and neuropodia 9–11 chaetae. Abdominal chaetae few, slender (4 µm thick) capillaries in both rami; upper notochaetae and lower neurochaetae twice as long as remaining chaetae (Figure 2G).
Pygidium divided by vertical cleft, with three short simple cirri, no intercirral processes (Figure 1F).
Juveniles with 11–12 chaetigers, differing from adults by smaller number of thoracic chaetigers; prostomium shorter, separated from peristomium by wide furrow; branchial filament arising from segmental border between chaetigers 2 and 3 (Figure 1E).
DISTRIBUTION
South-east Atlantic Ocean, Congo deep-sea fan.
REMARKS
This species is most similar to Cossura brunnea Fauchald, Reference Fauchald1972 described from the Pacific Ocean between Mexico and Oregon. Also reported from the US Atlantic slope (New England to North Carolina), C. brunnea is found from depths of 1600 to 2200 m (Hilbig, Reference Hilbig, Blake, Hilbig and Scott1996). It is similar to C. platypus sp. nov. in the attachment of the branchial filament and the number of thoracic chaetigers. The main difference is the shape of the prostomium, which is broadly triangular and together with the peristomium forms an evenly tapering cone in C. brunnea. In C. platypus sp. nov. the prostomium is widely rounded anteriorly and the posterior prostomial ring is abruptly expanded. Another characteristic of C. brunnea is the dark pigmentation of the body, whereas C. platypus sp. nov. is uniformly pale. The anterior and posterior chaetae are more similar in thickness in C. platypus sp. nov. than in C. brunnea. Cossura coasta recorded in West African waters differs from C. platypus sp. nov. in its conical prostomium and the presence of only short coarse chaetae in the neuropodia, while C. platypus sp. nov. has long and short chaetae in both rami; the number of thoracic chaetigers is unknown for this species. In the first description of C. coasta anal cirri are shown as distally branched (Kitamori, Reference Kitamori1960); however, in specimens from South Africa, they are simple (Day, Reference Day1967). It could mean that another, undescribed cossurid species probably inhabits African waters. Cossura rostrata Fauchald, Reference Fauchald1972 has similar distribution of short and long chaetae; it differs from C. platypus sp. nov. in the conical shape of the prostomium and 19–21 thoracic chaetigers followed by 10–15 intermediary segments of similar shape, while in C. platypus sp. nov. the segments become longer in the abdominal region. Cossura candida Hartman, Reference Hartman1955 differs from C. platypus sp. nov. in the conical shape of the prostomium and 24–35 thoracic chaetigers. The description of C. delta Reish, Reference Reish1958 is quite short, without indication of the number of thoracic chaetigers; however, supposedly it could be 18–25, because curved limbate chaetae which are limited to the thoracic region in distribution are present in 18–25 chaetigers. In C. delta the prostomium is conical with thickened chaetae present in neuropodia only, whereas in C. platypus sp. nov. they present in both rami. Cossura keablei Zhadan, Reference Zhadan2015, described from Fiji, is similar in the shape of the prostomium, but the branchial filament is attached to the anterior part of chaetiger 3, the posterior prostomial ring has a midventral notch, and this species has 22–26 thoracic chaetigers. Cossura duplex Tamai, Reference Tamai1986 is similar in the number and shape of thoracic chaetigers but differs in the conical shape of the prostomium and the coarse, flattened thoracic neurochaetae. In C. bansei Hilbig, Reference Hilbig, Blake, Hilbig and Scott1996 the branchial filament arises from the posterior border of chaetiger 3, sometimes appearing to originate on chaetiger 4; this species has a conical prostomium and 20–31 thoracic chaetigers.
Family COSSURIDAE Day, 1963
Genus Cossura Webster & Benedict, Reference Webster and Benedict1887
Cossura flabelligera sp. nov.
(Figures 3–5)

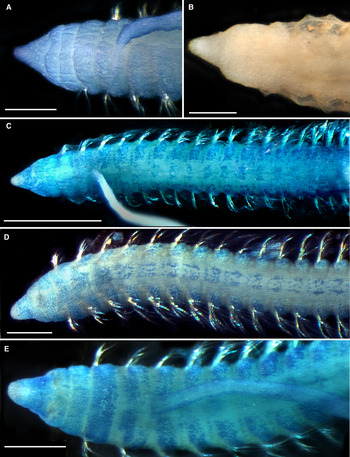
Fig. 3. Cossura flabelligera sp. nov., A–C, mucous sheath removed; (A) holotype, (MNHN-IA-TYPE 1784), dorsal view; (B) paratype, (MNHN-IA-TYPE 1785), dorsal view, methylene blue staining; (C) paratype, (MNHN-IA-TYPE 1787), lateral view; (D) paratype, (MNHN-IA-TYPE 1786), dorsal view, covered with mucous sheath. Scale bars: A, 1 mm; B, 500 µm; C, 200 µm; D, 300 µm.

Fig. 4. Cossura flabelligera sp. nov., A, C–E, mucous sheath removed, methylene blue staining; (A) paratype, (MNHN-IA-TYPE 1785), anterior end, dorsal view; (B) paratype, (MNHN-IA-TYPE 1786), anterior end, dorsal view; (C) paratype, (MNHN-IA-TYPE 1787), dorsal view; (D) paratype, (MNHN-IA-TYPE 1787), ventral view; (E) paratype, (MNHN-IA-TYPE 1787), anterior end, dorsal view. Scale bars: A, B, D, E, 200 µm; C, 500 µm.

Fig. 5. Cossura flabelligera sp. nov., SEM, A–C, with mucous sheath, D–H, mucous sheath removed, (A) paratype, (MNHN-IA-TYPE 1787), lateral view; (B) same specimen, surface of mucous sheath close up view; (C) paratype, (MNHN-IA-TYPE 1787), anterior end, dorsal view; (D) paratype, (MNHN-IA-TYPE 1787), anterior end, lateral view, (E) same specimen, parapodia of chaetigers 2–5; (F) paratype, (MNHN-IA-TYPE 1787), parapodia of chaetigers 3–4; (G) paratype, (MNHN-IA-TYPE 1787), piece of abdomen, dorsal view; (H) same, close-up. Scale bars: A, G, 300 µm; B, D, 100 µm; C, E, F, 30 µm; H, 10 µm.
TYPE MATERIAL
Holotype: (MNHN-IA-TYPE 1784), RV ‘Pourquoi Pas?’, CONGOLOBE cruise, ROV ‘Victor 6000’, dive 491, 3.1.2012, 19:36:21, large blade cores, GCL2bis, layer 0–5 cm, fraction 1000 µm, location CoL C, 06°42.092′S 005°29.306′E, depth 4846 m. Paratypes: (MNHN-IA-TYPE 1785), 1 specimen, RV ‘Pourquoi Pas?’, CONGOLOBE cruise, ROV ‘Victor 6000’, dive 491, 3.1.2012, 19:36:21, large blade cores, GCL2bis, layer 0–5 cm, fraction 1000 µm, location CoL C, 06°42.092′S005 29.306′E, depth 4846 m; (MNHN-IA-TYPE 1786), 6 specimens, RV ‘Pourquoi Pas?’, CONGOLOBE cruise, ROV ‘Victor 6000’, dive 491, 3.1.2012, 04:56:23, large blade cores, GCL5, layer 5–10 cm, fraction 500 µm, location CoL C, 06°42.080′S 005°29.287′E, depth 4846 m; (MNHN-IA-TYPE 1787), 21 specimens, RV ‘Pourquoi Pas?’, CONGOLOBE cruise, ROV ‘Victor 6000’, dive 492, 5.1.2012, 02:58:38, large blade cores, GCL1, layer 5–10 cm, fraction 500 µm, location CoL B, 06°25.248′S 005°49.707′E, depth 4719 m.
ETYMOLOGY
The name refers to the polychaete genus Flabelligera having a thick mucous cover which resembles the most remarkable character of this cossurid species – the mucous sheath.
DESCRIPTION
Colour in alcohol uniformly white or pale yellowish, chaetae golden to brown (Figure 3A, C). Holotype 6.5 mm long, with 17 thoracic chaetigers, 340 µm wide, abdomen covered by a sheath (Figure 3A). All specimens incomplete, represented by short fragments with 18–40 chaetigers. Bodies of most specimens including chaetae covered by thick mucous sheath resembling tunic of flabelligerids (Figures 3D, 4B & 5A–C). Thoracic width up to 350 µm, 16–19 thoracic chaetigers. Thoracic segments not inflated in smaller specimens, slightly inflated in larger ones, with wide intersegmental furrows and middorsal groove (Figures 3B & 4C, E). Segments becoming longer and thicker in the abdominal region (Figure 3A).
Prostomium conical, with rounded tip, about as long as wide; lateral margins straight to slightly concave, posterior prostomial ring same length as peristomium, without midventral notch (Figures 3D, 4A–E & 5C). Nuchal organs not seen. Branchial filament arising from posterior part of chaetiger 2 (Figures 3B, C, 4A, C, E & 5D).
Chaetiger 1 with uniramous parapodia, all subsequent segments with biramous parapodia; chaetae emerging from low lobes; parapodial rami situated very close to each other. All chaetae hirsute capillaries with smooth shafts, arranged in two distinct rows; anterior chaetae in each thoracic segment thicker and shorter than posterior chaetae (Figures 3C & 5D–F). First chaetiger bearing three thicker anterior and four thinner posterior chaetae. In other segments, four neurochaetae of anterior row notably thicker (shaft 9 µm, hirsute part 11 µm thick) than four anterior notochaetae (shaft 6 µm, hirsute part 8 µm thick); posterior chaetae 4 µm thick, five chaetae in each ramus (Figures 3C & 5D–F). Abdominal chaetae 3–5 long slender capillaries in both rami, 2.5–3 µm thick (Figure 5G, H). Pygidium not found.
Tip of prostomium unstained in methylene blue, solid stripes on dorsal side of chaetigers 1–3, lateral and median patches on dorsal sides, smaller lateral and larger median patches on ventral sides of other chaetigers (Figure 4C–E).
DISTRIBUTION
South-east Atlantic Ocean, Congo deep-sea fan.
REMARKS
Cossura flabelligera sp. nov. is similar to C. longocirrata Webster & Benedict, Reference Webster and Benedict1887, which according to Fournier & Petersen (Reference Fournier and Petersen1991), is an arctic-boreal species distributed in the North Atlantic. Cossura flabelligera sp. nov. is similar in the position of the branchial filament, the shape of the prostomium, the number of thoracic chaetigers, and the shape and distribution of the chaetae. The main difference distinguishing C. flabelligera sp. nov. from C. longocirrata is the thick mucous sheath covering the entire body including the chaetae. Moreover, the noto- and neurochaetae are similar in thickness in C. longocirrata but the neurochaetae are thicker in C. flabelligera sp. nov. Cossura heterochaeta Orensanz, Reference Orensanz1976 is similar to C. flabelligera sp. nov. in its conical prostomium, position of the branchial filament and presence of thickened anterior chaetae, but there are two types of anterior thickened chaetae in C. heterochaeta – with and without an arista – whereas in C. flabelligera sp. nov., they uniformly have aristas. In addition, C. flabelligera sp. nov. has more anterior segments (16–19 instead of 12–14 as in C. heterochaeta). Cossura coasta has a branchial filament arising from chaetiger 3 and only coarse thick chaetae in the neuropodia, but in C. flabelligera sp. nov., the branchial filament arises from chaetiger 2, and the neuropodia contain both thick and slender chaetae. Some cossurid species produce mucous continuously along the entire length of the body, resulting in the presence of a partial mucous coating that may be encrusted with sediment particles but probably does not represent a proper tube. Except for some mucous among the chaetae, preserved cossurids are typically free of any surrounding or attached tubelike structures (Hilbig, Reference Hilbig, Blake, Hilbig and Scott1996). The Flabelligera-like tunic of Cossura flabelligera sp. nov. seems to be a character unique for this species.
ACKNOWLEDGEMENTS
We thank the captain and crew of the RV ‘Pourquoi Pas?’, the team of the ROV ‘Victor’, and the chief scientists of the cruises, Karine Olu (IFREMER Centre Bretagne, Institut Carnot Ifremer-EDROME Laboratoire Environnement Profond) and Christophe Rabouille (Laboratoire des Sciences du Climat et de l'Environnement, CNRS), for collecting and providing us with the material. We thank Patrick Briand, Philippe Noël, Olivier Mouchel and Jozée Sarrazin for their help on board during the WACS cruise and Manuella Rabiller, Erwan Guillon, Lenaick Menot for faunal sorting and identifications of the polychaete families. We are grateful to Tarik Meziane and Laure Corbari (MNHN) for their help with deposition of type material and Andrew Gebruk (Shirshov Institute of Oceanology, Moscow) for involving us in this project. Two anonymous reviewers improved the manuscript.
FINANCIAL SUPPORT
This study was supported by grants 15-04-05875 and 16-04-00343 from the Russian Foundation of Basic Research. The scanning electron microscopy investigations were supported by the Russian Scientific Foundation, grant 14-50-00029. The electron microscopy was performed at the User Facilities Center of M.V. Lomonosov Moscow State University.