Introduction
The deep sea lacks primary photosynthetic producers; it is often an oligotrophic environment compared with the shallow sea. In such oceanic deep environments, the sedimentation of marine snow (i.e. the mucus-laden remains and excretions of organisms living in the shallow sea) and the diurnal vertical movement of zooplankton provides a route for supplying photosynthetic products from the shallow sea to the deep sea (Wiebe et al., Reference Wiebe, Madin, Haury, Harbison and Philbin1979). Nutrients are also supplied to the deep sea through the influx of plant remains. There is constant runoff of seagrass from shallow-water communities to deeper ecosystems. During floods and storms, large amounts of seagrass flow from the communities and eventually sink to the deep-sea bottom (Menzies et al., Reference Menzies, Zaneveld and Pratt1967; Menzies & Rowe, Reference Menzies and Rowe1969). On these sea beds, abyssal sea urchins directly consume large amounts of eelgrass leaves at depth of 2500–4000 m; sea cucumbers ingest nutrients from the decomposed seagrass remains (Suchanek et al., Reference Suchanek, Williams, Ogden, Hubbard and Gill1985).
On deep-sea bottoms near land, there are characteristic inflows of terrestrial plant remains, such as fallen leaves and twigs, and occasional large pieces of wood. These terrigenous plant remains are decomposed to differing degrees by fungi, bacteria and other microorganisms; they are utilized by diverse benthic invertebrates (Sridhar, Reference Sridhar2012; Xu et al., Reference Xu, Pang and Luo2014). For example, Xylophaga bivalves bore into wood; after the death of these borers, the remaining holes are used by sipunculids, galatheoids and polychaetes belonging to Cirratulidae, Maldanidae, Capitellidae and Ampharetidae. Various sessile organisms (e.g. bivalves, bryozoans and barnacles) also use wood as a substrate (Wolff, Reference Wolff1979).
Terrestrial plant remains on deep-sea bottoms are also used by three described species of tube-bearing polychaetes belonging to the genus Anchinothria (Onuphidae), which live in portable tubes that are characteristically encrusted with submerged fallen leaves or twigs of land plants, spines of sea urchins or sponges, foraminiferas etc. (Table S1). While the use of fragments of fallen leaves/twigs as material for portable tubes is common among trichopteran larvae in inland waters (Thomas et al., Reference Thomas, Frandsen, Prendini, Zhou and Holzenthal2020), it is remarkable that deep-sea polychaetes have a similar lifestyle. Generally, polychaetes belonging to Onuphidae live on sea bottoms as settling semi-burrowers (Onuphis) or crawling tube bearers (Hyalinoeciinae: Anchinothria, Hyalinoecia, Leptoecia, Nothria). In Onuphinae, Diopatra is the most common leaf-utilizing species. Diopatra is a semi-burrowing species found in the intertidal and subtidal mud flat, which is known to utilize leaves, shells and seaweed to encrust the upper tube exposed on the sand (Paxton, Reference Paxton1998). As to Hyalinoeciinae, Leptoecia and Hyalinoecia worms live in rigid transparent tubes made of onuphic acid (Graham et al., Reference Graham, Kelly, Pautard and Wilson1965; Budaeva et al., Reference Budaeva, Pyataeva and Meissner2014), while Nothria worms live in portable tubes encrusted with components such as sand, stones and shell fragments (e.g. Fauchald, Reference Fauchald1982; Imajima, Reference Imajima1999). It is intriguing to consider how the use of fallen leaves/twigs as portable tube material evolved in an Onuphidae clade.
Anchinothria has been described in previous studies as morphologically sister to Nothria and Hyalinoecia (Budaeva et al., Reference Budaeva, Schepetov, Zanol, Neretina and Willassen2016). On the other hand, ecological and molecular phylogenetic knowledge of Anchinothria is limited due to its deep-sea habitat and rare occurrence which makes it difficult to collect. In the genus, A. cirrobranchiata is collected only in the seas around Japan, and known to have a tube with scattered pieces of leaves and wood (Table S1, Imajima, Reference Imajima1999). Because deep-sea life utilizing terrigenous plant remains is unique, it is intriguing to determine what plant species are used as tube materials, if the plant debris on portable tubes is also used as food, and when the utilization of land plant debris originated in the phylogeny of Onuphidae.
From the deep-sea bottom near land in the Japanese Archipelago, we collected the deep-sea polychaete Anchinothria cirrobranchiata, which uses fallen leaves of land plants as material for its portable tube. To address the above-mentioned questions, we observed the feeding biology of these polychaetes, investigated the composition of the materials encrusting their portable tubes, identified the plant species of the leaves on the tubes, and performed a molecular phylogenetic analysis of Onuphidae.
Materials and methods
Sample collection
Deep-sea benthic organisms were collected by trawling a dragnet at a depth of 200–300 m off the coast of Owase, Mie Prefecture (34°01′N 136°22′E), on 26 February 2017. Owase has a very high amount of precipitation and is surrounded by extensive evergreen forests. In addition to various benthic organisms, extensive terrigenous plant remains were collected in this bottom trawl. Eighteen Anchinothria cirrobranchiata were collected. The worms were identified based on Imajima (Reference Imajima1999); branchiae first present from chaetiger 11 and branched with five filaments (Figure 1A), bidentate pseudocomposite hooks are seen in chaetiger 1–4 (Figure 1B), scoop-shaped pectinate chaetae are seen from chaetiger 2 (Figure 1C) and subacicular hooks start from chaetiger 9–11 (Figure 1D). Three of the collected Anchinothria worms were used for ecological observations and the others were fixed in 70% ethanol for detailed observation of tube materials and in 99% ethanol for molecular phylogenetic analysis; the voucher is stored at Seto Marine Biological Laboratory (specimen number: SMBL-V0637).

Fig. 1. Ecology and morphology of Anchinothria cirrobranchiata. (A) dorsal view which shows branchiae. Branchiae start from chaetiger 11, and branched with five filaments from chaetiger 17 (arrowhead); (B) bidentate pseudocomposite hooks in chaetiger 1; (C) scoop-shaped pectinate chaetae in chaetiger 3; (D) subacicular hooks in chaetiger 9; (E) a tube cross-section, showing a thin film stretched inside a parchment-like layer; (F–G) tubes encrusted with fallen leaves and twigs; (H) A worm moving forward by moving the front of its body (arrowhead) out of the tube; (I) body filled with numerous eggs; (J) front view of a worm foraging for fallen leaves; Scale bars: (A) 1 mm; (B–D) 200 μm; (E) 1 mm; (F–H) 1 cm; (I, J) 1 mm.
Ecological observations and identification of the leaves used for tubes
One of the Anchinothria polychaetes collected was kept in an aquarium at Toba Aquarium with filtered natural seawater regulated to 10°C, and its biological photograph were taken and behaviour was recorded with TG-4 (OLYMPUS, Tokyo, Japan) at 1.0–2.5×. Two polychaetes were kept in Petri dishes with artificial seawater at a room temperature of 25 ± 3°C for observation.
The individual kept in an aquarium was provided autochthonous fallen leaves collected with polychaetes and its behaviour was observed. The individuals in the Petri dishes were provided fallen land leaves Quercus glauca, Quercus phillyraeoides, Zelkova serrata and Castanopsis sieboldii soaked overnight in 1 litre of seawater to prevent changes in salinity and runoff of secondary metabolites into the culturing seawater, and time-lapse photography was performed from a vertical position for 24 h. To determine the structure and constitution of tube materials, a single tube was cut into thin slices and its cross-sectional structure was observed under a microscope at 10×. We examined all materials in each portable tube for the 15 fixed specimens; we identified the plant species of all leaf fragments based on the morphology of nervure and leaf margin. The area of all tube-encrusting materials was measured using ImageJ v. 1.53 and the percentages of area of each material against total area of materials were calculated.
Molecular phylogenetic analysis
We performed direct polymerase chain reaction using Ampdirect® Plus reagent (Shimadzu) to amplify DNA from ethanol-preserved skin (3–5 mg dry weight) of A. cirrobranchiata. The tissue was dried, soaked in 50 μl lysis buffer (20 mM Tris-HCl at pH 8.0, 5 mM ethylenediaminetetraacetic acid, 400 mM NaCl, 3% sodium dodecyl sulphate, and 200 μmol ml−1 proteinase), and incubated at 60°C for 12 h. Polymerase chain reaction was used to amplify the 16S rRNA gene (~500 bp amplicon) using primers 16SarL and 16S-OnuR (Palumbi et al., Reference Palumbi, Martin, Romano, McMillan, Stice and Grabowski1991; Budaeva et al., Reference Budaeva, Schepetov, Zanol, Neretina and Willassen2016), and 18S rRNA gene (~1850 bp amplicon) using three primer pairs: 18e + 18L; 18F509 + 18R; 18F997 + 18D843 (Elwood et al., Reference Elwood, Olsen and Sogin1985; Hillis et al. Reference Hillis, Ammerman, Dixon and de Sá1993; Struck et al., Reference Struck, Hessling and Purschke2002). The purified polymerase chain reaction products (≥50 ng μl-1) were directly sequenced (Eurofins, Japan) using the above primers. The resulting sequences were deposited in the DDBJ/EMBL/GenBank databases under the accession numbers listed in Table S2. With the obtained sequences, the phylogenetic position of Anchinothria were analysed based on the phylogenetic tree derived in Budaeva et al. (Reference Budaeva, Schepetov, Zanol, Neretina and Willassen2016). Our dataset includes the sequences of the representative OTUs of each major clade of Onuphinae used in this study. Eunicidae, which comes outside Onuphidae, was used as an outgroup. The cited sequences are listed in Table S2. These sequences were aligned using the Muscle package (Edgar, Reference Edgar2004) implemented in SeaView software (Galtier et al., Reference Galtier, Gouy and Gautier1996; Gouy et al., Reference Gouy, Guindon and Gascuel2010) with the default settings. We used Gblocks v. 0.91b (Castresana, Reference Castresana2000; Talavera & Castresana, Reference Talavera and Castresana2007) to eliminate any ambiguously aligned regions of each sequence. The 16S and 18S gene sequence lengths before and after Gblocks treatment were 448 and 443 bp, 1774 and 1636 bp, respectively. Bayesian and maximum likelihood phylogenetic analyses were performed on the combined 16S + 18S dataset using MrBayes v. 3.1.2 (Ronquist & Huelsenbeck, Reference Ronquist and Huelsenbeck2003) and RAxML v. 7.4.2 (Stamatakis, Reference Stamatakis2006) implemented in raxmlGUI v. 1.31 (Silvestro & Michalak, Reference Silvestro and Michalak2012). We selected the GTRGAMMA model for the RAxML analysis and used Kakusan4 software (Tanabe, Reference Tanabe2011) to select appropriate models for the MrBayes analysis. The model selected for the MrBayes analysis was GTR_GAMMA for each gene. Two independent runs of the Metropolis-coupled Markov chain Monte Carlo procedure were performed simultaneously for 2,000,000 generations. The convergence of MCMC was confirmed with Tracer v1.7.2.
Results
Materials of portable tubes
The portable tube of Anchinothria cirrobranchiata was long, straight, and dorsoventrally flattened; it comprised a thin inner film and parchment-like layer with accretion glued on it (Figure 1E). Each tube had an opening at each end. The accretion comprised sand grains and fragments of fallen leaves and twigs; the fragments of fallen leaves and twigs mechanically reinforced the tube (Figure 1F–G). Table 1 shows the percentages of materials glued on the portable tubes. These glued-on leaves were chartaceous, blackish, tough, and generally fragmented. All plants were evergreen tree species, except one deciduous tree species, Zelkova serrata.
Table 1. Percentage of materials glued on portable Anchinothria cirrobranchiata tubes (N = 15)

Moving and feeding behaviour
An Anchinothria worm bearing its portable tube advanced by protruding the anterior part of its body to a great extent from the tube, anchoring the body to the substrate with one to three parapodia, and drawing the posterior part of the body with the tube forward (Figure 1H). Anchinothria worms were also observed to turn anterior-posteriorly and up-down within their portable tube; for example, when they were moving in one direction and their head hit the aquarium wall, they turned around in the portable tube, changed their body direction to the opposite side, and continued moving. We also observed worms turning up and down in the tube while foraging, reversing their body orientation against the tube. Eggs were held in the body cavity of the exposed anterior part (Figure 1I). The average diameter of the eggs was 150 μm (±14 μm, N = 20).
The Anchinothria worms kept with autochthonous brownish fallen leaves in aquaria were observed to use their jaws to cut the leaves and glue them onto their tubes. The worms were also observed feeding on leaves of their own tube material by holding them with the first and second parapodia, then cutting them with their jaws and taking them in (Figure 1J). However, the worms did not feed on the fallen leaves collected on land.
Molecular phylogenetic analysis
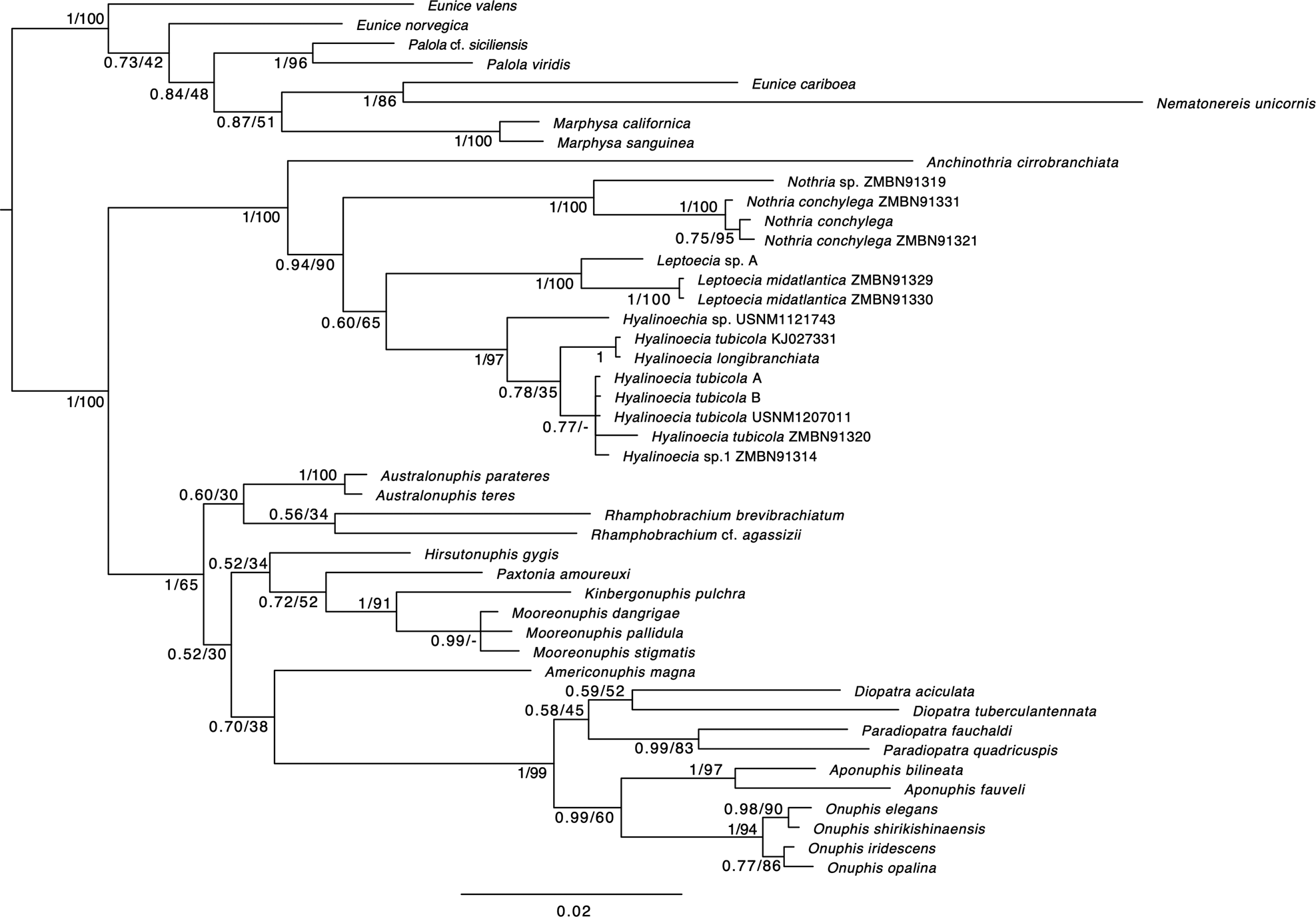
The phylogenetic tree derived from Bayesian method is shown in Figure 2. The tree topology was consistent with the ML and Bayesian methods except for specimens inside Hyalinoecia and Mooreonuphis. Hialinoeciinae including Hyalinoecia, Leptoecia, Nothria and Anchinothria was monophyletic (Bayesian posterior probability: PP = 1, bootstrap support value: BS = 100). Within this clade, Anchinothria was sister to the remaining genera, which were also monophyletic (PP = 0.94, BS = 90). Each of Nothria, Leptoecia and Hyalinoecia was monophyletic (Figure 2).

Fig. 2. Bayesian/ML tree of the combined 16S and 18S rRNA sequences. The numbers at the nodes are Bayesian posterior probability values and maximum likelihood bootstrap support values. See Table S2 for the list of accession numbers.
Discussion
Utilization of fallen leaves and twigs
The leaves and twigs glued on tubes presumably contribute to reinforcing the tube, and they serve as both camouflage and portable food. The dominant species belonged to two genera of Fagaceae, Quercus and Castanopsis, both of which are dominant genera in the natural forests around Owase. These results suggest that the life of Anchinothria is supported by the nearby terrestrial evergreen forest ecosystem via the substantial influx of forest litter caused by high precipitation.
The behavioural observations that the worms turn up and down, and reverse their body orientation against the tube suggested that the tube does not have a dorsoventral or anterior–posterior orientation. The moving behaviour with portable tube has been observed in detail in Hyalinoecia artifex (Meyer et al., Reference Meyer, Wagner, Ball, Turner, Young and Van Dover2016), and Anchinothria cirrobranchiata was found to move and change the body direction in a similar method. On the other hand, A. cirrobranchiata has thick tubes, unlike Hyalinoecia which builds fitted tubes.
From the observation in aquarium tanks, it is supposed that Anchinothria use fallen leaves deposited in the deep sea not only as tube-encrusting material but also as food. As to fallen leaves on land, it is possible that the small size of the water system in the Petri dish stressed the worms and prevented them from feeding. To determine whether Anchinothria will feed on undecayed fallen leaves, it is necessary to conduct another test in the same environment in which they fed on decayed fallen leaves.
Reproductive ecology
The eggs of A. cirrobranchiata were small-sized similar to many of tube-bearing onuphid species (Giangrande, Reference Giangrande1997). Some Onuphid species, such as Notonuphis antarctica, are known to have eggs larger than 1 mm and nurse in adult tubes, but species with eggs as small as 200 μm are generally known to spawn eggs (Paxton, Reference Paxton1986). From this knowledge, it is assumed that A. cirrobranchiata spawn its eggs.
Evolution of tube-bearing life
The phylogenetic tree suggests that Anchinothria is sister to other Hyalinoeciinae genera: Hyalinoecia, Leptoecia and Nothria, which were also monophyletic. This result contradicts the morphological assumption in previous research that the subfamily Hyalinoeciinae was divided into two groups: Hyalinoechia group including Hyalinoecia, Leptoecia and Hyalospinifera, and Nothria group including Nothria and Anchinothria (Budaeva et al., Reference Budaeva, Schepetov, Zanol, Neretina and Willassen2016). Molecular phylogenetic trees do not provide well-supported estimates of phylogenetic relationships among four genera due to low support values in both previous and present studies. Further taxon sampling and analysis with more genetic markers may resolve their relationships.
Onuphinae have developed non-portable tubes encrusted with sediment materials such as shell fragments, algae, fallen leaves, sponge rods etc. (Fauchald, Reference Fauchald1982; Paxton Reference Paxton1986), and in the Hyalinoeciinae, the accretion of sediment material onto portable tubes is only seen in the Nothria and Anchinothria. Nothria species build tubes with shells and gravel fragments, but Anchinothria has a diverse selection of tube-encrusting materials (Table S1). It is hoped that phylogenetic analysis of Anchinothria species which utilize a variety of case-encrusting materials will elucidate the evolutionary path of case-encrusting materials selection and the ecology and morphology acquired in the use of each material.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315422000807.
Data
Gene sequences analysed during this study are deposited to GenBank.
Acknowledgements
We thank all the staff of the Toba aquarium for supporting our sampling.
Author contributions
LY designed the study; LY and TM carryied out the study; LY and MK analysed the data; LY, TM and MK interpreted the findings and wrote the article.
Financial support
This work is supported by Japan Ministry of Education, Culture, Science, Sports, and Technology Grant-in-Aid for Scientific Research (15H02420, 20H03321; https://www.jsps.go.jp/english/index. html) for Makoto Kato and Grants-in-Aid for Scientific Research Grant number 18J22890 for Luna Yamamori.
Conflict of interest
The authors declare none.
Ethical standards
Not applicable.