INTRODUCTION
Hartmann-Schröder (Reference Hartmann-Schröder1958) proposed Hesionura for Hesionura fragilis Hartmann-Schröder, Reference Hartmann-Schröder1958. It was originally diagnosed by a rectangular, longer than wide, prostomium. Hartmann-Schröder (Reference Hartmann-Schröder1960) later proposed Eteonides also diagnosed by a longer than wide prostomium. These two genera are so similar that Hartmann-Schröder herself (Reference Hartmann-Schröder1963) regarded them as synonyms, but quite surprisingly, she preferred to use Eteonides over Hesionura. It was Hartman (Reference Hartman1965) who corrected the priority, since Hesionura is the senior synonym over Eteonides.
Hesionura, along with the genera Pseudomystides, Pelagobia, Eteone, Mysta and Hypereteone, belongs to the subfamily Eteoninae, a group defined by lacking a dorsal cirri on the second (Eteone, Mysta and Hypereteone) or third segment, and two or three pairs of tentacular cirri (Bergström, Reference Bergström1914; Hartmann-Schröder, Reference Hartmann-Schröder and Dahl1971). More recently, Pleijel (Reference Pleijel1991) included a series of other genera in Eteoninae: Pirakia, Pterocirrus, Eumida, Sige, Eulalia, Protomystides, Mystides, Galapagomystides, along with Pseudomystides, Hesionura and Eteone (with Mysta and Hypereteone synonymized with Eteone). This clade is supported by the presence of nuchal organs, with a latero-posterior protrusion in the prostomium, and the presence of dorsal cirri on the third segment.
Hesionura is currently characterized by well-developed segments all along the body, an elongated prostomium, usually much longer than wide, paired frontal antennae and palps, no median antenna, small eyes in the posterior-dorsal part of the prostomium and nuchal organs in a groove between the prostomium and the first segment. The proboscis has diffusely distributed, rounded or conical papillae. Three pairs of tentacular cirri are present on the first two segments. The tentacular cirri are cylindrical and have short cirrophores and long cirrostyles, except for the ventral pair, which is somewhat flattened. Dorsal cirri are absent on the third segment. Dorsal and ventral cirri are long, cylindrical or rounded. The parapodia are uniramous. The pre-chaetal lobes are symmetrical and bilobate. Chaetae are present from the third segment. The rostrum of the shaft may be unidentate, bidentate, tridentate, or quadridentate, whereas articles are short, strongly serrulated and present in various forms. The pygidium has one pair of long, cylindrical cirri (Pleijel, Reference Pleijel1991, Reference Pleijel1993a, Reference Pleijelb; Viéitez et al., Reference Viéitez, Alós, Parapar, Besteiro, Moreira, Núñez, Laborda and San Martín2004).
We observed one small tooth on the medial surface of each large tooth on the chaetal rostrum of all the new species (Figures 9C, 19A, 26C), a character previously unreported for the genus (note: these small teeth are not included when characterizing a chaeta as bidentate, tridentate or quadridentate). A median pygidial papilla had not been observed by Pleijel (Reference Pleijel1993a) or Viéitez et al. (Reference Viéitez, Alós, Parapar, Besteiro, Moreira, Núñez, Laborda and San Martín2004), although it was reported by Hartmann-Schröder (Reference Hartmann-Schröder1960) for Eteonides and subsequently in Hesionura shandongensis Zhao & Wu, Reference Zhao and Wu1991. In our study, a pygidial papilla was observed in Hesionura biformata sp. nov., H. variodentata sp. nov., H. carioca sp. nov. and H. lupina sp. nov. It is likely that pygidial papillae have passed unnoticed by other taxonomists because of their small size.
There are currently 13 described species in the genus, of which only Hesionura laubieri (Hartmann-Schröder, Reference Hartmann-Schröder1963) was already known from Brazil (Amaral et al., Reference Amaral, Nallin, Steiner, Forroni and Gomes-Filho2012). Besides redescribing H. laubieri based on newly collected material, we describe herein six new species, bringing the total in the genus to 19. This study is part of an ongoing extensive survey of the Phyllodocidae from southern and south-eastern Brazil.
MATERIALS AND METHODS
Collection of biological material
Altogether 251 individuals were examined, mostly from collections of living material made by the first author in 2009, 2010 and 2011 in the States of Rio de Janeiro, Paraná and Santa Catarina, in south-eastern and southern Brazil (Figure 1). Other examined specimens had previously been collected during the Habitats/Petrobrás and Revizee Score Sul projects. Additional material, used for comparisons, was examined at the Zoological Museum, Natural History Museum of Denmark (ZMUC) and at the Zoologische Museum of Hamburg. Holotypes are deposited at the Zoology Museum from Universidade Estadual de Campinas (ZUEC); paratypes are deposited at ZUEC and ZMUC.

Fig. 1. Collection sites in southern and south-eastern Brazil, off the States of Santa Catarina, São Paulo, Rio de Janeiro and Espírito Santo.
Animals were collected from sub-littoral estuarine soft bottoms to exposed oceanic beaches, and from shallow platform depths down to 98 m. Sediment samples were taken with a Petit Ponar grab and corers. Samples were sieved through 260 µm meshes to remove fine sediments. Live specimens from shallow sites were placed in plastic containers with seawater and transported to the lab in coolers with ice. Animals were kept in aquaria under room temperature, anaesthetized with menthol in seawater for more detailed morphological analyses and subsequently fixed in 4% formalin and preserved in 70% ethanol.
Identification routines
Semi-permanent slides were mounted with gel and glycerin (Amaral & Nonato, Reference Amaral and Nonato1987), for more detailed observations of the morphology of papillae, parapodia and chaetae. The same technique was used for whole organisms, in the case of smaller animals, which improved the visualization of the degree of fusion of the anterior segments and the distribution of tentacular cirri. The specimens were identified using stereoscopic and optical microscopes and, in some cases, scanning electron microscopy. Descriptions were prepared following the terminology adopted by Pleijel (Reference Pleijel1993a) for the family and are presented in the sequential order illustrated in Figures 2–5 and Table 1. Measures of median width included only parapodia, without considering the chaetae. For improved contrast in checking for papillae, Shirlastain A dye was used on some specimens.

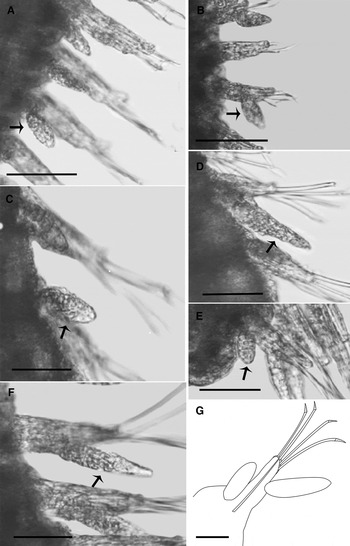
Fig. 2. Terminology used for the descriptions of Hesionura: (A, C) Dorsal view of H. laubieri; (B) Ventral view of H. laubieri; (A, B) Morphology of the prostomium and anterior segments of H. laubieri; (A) Prostomium elongated with frontal protuberance, where antennae and palps are located; (C) Undivided proboscis of H. laubieri (i.e. the anterior and posterior portions do not differ from one another). Formula for the tentacular cirri: (A, B) 1 + 2 + 0 = First dorsal tentacular cirri, second segment with dorsal and ventral tentacular cirri; 0 = Loss of dorsal cirri. Tentacular cirri morphology: (A–C) First and second pairs of dorsal tentacular cirri cylindrical; (B) Cylindrical ventral tentacular cirrus with inflated base and tapering distal end; (D) Ventral tentacular cirrus oval to elongated of H. longissima sp. nov. (arrow); (E) Rounded ventral tentacular cirrus of mature H. longissima sp. nov. (arrow); (F) Ventral tentacular cirrus with inflated base and tapering distal end of H. carioca sp. nov. (arrow). Scale bars: A–B, 0.05 mm; C, 0.06 mm; D–F, 0.05 mm.

Fig. 3. Terminology used for the descriptions of H. laubieri, H. ceciliae sp. nov., H. biformata sp. nov., H. variodentata sp. nov., H. carioca sp. nov., H. longissima sp. nov. and H. lupina sp. nov.; morphology of the parapodial dorsal cirri, in anterior view (arrow): (A) Dorsal cirrus cylindrical of H. laubieri (arrow); (B) Dorsal cirrus oval to elongated (arrow); (C) Dorsal cirrus cylindrical to rounded of H. longissima sp. nov. (arrow); (D) Dorsal cirrus cylindrical with rounded distal end of H. variodentata sp. nov. (arrow). Scale bars: A–D, 0.05 mm.

Fig. 4. Terminology used for the descriptions of H. laubieri, H. ceciliae sp. nov., H. biformata sp. nov., H. variodentata sp. nov., H. carioca sp. nov., H. longissima sp. nov. and H. lupina sp. nov.; morphology of the ventral cirri, in ventral view: (A) Oval ventral cirrus oval of H. longissima sp. nov.; (B) Long and slender ventral cirrus of H. laubieri; (C) Conical ventral cirrus of median segments of H. ceciliae sp. nov.; (D) Dorsal cirrus cylindrical and abruptly tapered at the distal end of H. biformata sp. nov. Scale bars: A, 0.01 mm; B, 0.05 mm; C, 0.01 mm; D, 0.05 mm.

Fig. 5. Terminology used for the descriptions of H. laubieri, H. ceciliae sp. nov., H. biformata sp. nov., H. variodentata sp. nov., H. carioca sp. nov., H. longissima sp. nov. and H. lupina sp. nov.; chaetae and anal cirri: (A) Compound spinigerous chaetae of H. longissima sp. nov.; (B) Cirriform anal cirri; arrow indicates pygidial papilla of H. biformata sp. nov. (arrow). Scale bars: A, 0.05 mm; B, 0.01 mm.
Table 1. Diagnostic characters from H. laubieri and from the new species of Hesionura from southern and south-eastern Brazil.

Photographs were taken with a Canon PowerShot S45 digital camera with 4.0 MP resolution coupled to the stereoscopic and optical microscopes. Scanning electron microscope photographs were taken from fixed specimens previously transferred to 1% osmium in distilled water for 1 h. The specimens were dehydrated in an alcohol series of 50%, 60% and 70% for 5 min each, followed by 90% and 95% for 15 min each, kept in 100% alcohol and then transferred to a MEV BAL-TEC CPD030 (SEM) for critical point drying. The specimens were sputter coated with gold at a thickness of 102 Å in a SCD-030-Balzers FL9496 sputter coater and subsequently examined in a JSM6360LZ GOAL at the Electron Microscopy Center at the Federal University of Paraná. The scanning photographs taken at the Zoological Museum of the University of Copenhagen followed the same methodology, with the exception of the replacement of osmium by acetone and use of an alcohol series of 90%, 99% alcohol; 2:1.
SYSTEMATICS
Family Phyllodocidae Ørsted, Reference Ørsted1843
Genus Hesionura Hartmann-Schröder, Reference Hartmann-Schröder1958
(=Eteonides Hartmann-Schröder, Reference Hartmann-Schröder1960)
Hesionura laubieri (Hartmann-Schröder, Reference Hartmann-Schröder1963)
(Figures 2A–C & 6–9)
Eteonides laubieri Hartmann-Schröder, Reference Hartmann-Schröder1963, 228; Laubier, Reference Laubier1967, 93.
Hesionura laubieri. Westheide, Reference Westheide1974, 4–6. pl. 1; Amaral et al., Reference Amaral, Nallin and Steiner2010, 63; Reference Amaral, Nallin, Steiner, Forroni and Gomes-Filho2012, 74.

Fig. 6. Hesionura laubieri. Dorsal views of parapodia: (A) Parapodium 10, dorsal cirrus; (B) Parapodium 10, ventral cirrus (arrow); (C) Parapodium 43, ventral cirri; (D) Parapodium 45 with dorsal (left) and ventral (right) cirri, dorso-ventral view. Scale bars: 0.05 mm.

Fig. 7. Hesionura laubieri: (A–C) Dorsal views of parapodia; (A) Parapodium 90; (B) Parapodium 91; (C) Posterior segments of a specimen in reproductive stage; (D) Pygidium with anal cirri. Scale bars: A–C, 0.05 mm; D, 0.07 mm.

Fig. 8. Hesionura laubieri, paratypes: (A) Dorsal view of parapodia; (B) Ventral cirri from median segments, ventral view. Scale bars: 0.05 mm.

Fig. 9. Hesionura laubieri: (A) Supraacicular chaetae; (B) Posterior parapodium, antero-lateral view; (C) Bidentate chaeta; (D) Unidentate chaeta. Scale bars: A, 0.003 mm; B, 0.01 mm; C, D, 0.005 mm.
TYPE MATERIAL
Paratypes. 8 specimens. ZMH–14150. Zapallar, Valparaiso, Chile.
MATERIAL EXAMINED
27 individuals, length from 0.5 to 4.0 mm and number of segments from 27 to 88. Rio de Janeiro: Barra da Tijuca Beach, 23°00′51″S 43°22′23″W, 10 June 2010 (11 specimens, ZUEC–POL–14335); Barra da Tijuca Beach, 23°00′51″S 43°22′23″W, 10 June 2010 (5 specimens, ZMUC–POL–2421); Cabo Frio, 10 Feb 2011 (5 specimens, ZUEC–POL–16848). Santa Catarina: Estaleiro Beach, 27°01′49″S 48°34′48″W, 20 April 2009 (4 specimens, ZUEC–POL–16670). Estaleiro Beach, 27°01′49″S 48°34′48″W, 20 April 2009 (4 specimens, ZUEC–POL–16857); collected in coarse sand beaches. Continental shelf in the Campos Basin: Hab17 Foz30 R3, 21°34′12.552″S 40°25′32.724″W, 28 m, 23 July 2009 (1 specimen, ZUEC–POL–16838).
DIAGNOSIS
Four chaetae per parapodium throughout the body, two bidentate supracicular and two subacicular chaetae, one of them bidentate.
DESCRIPTION
Largest examined specimen incomplete, with 15 segments, 1.0 mm in length, 0.02 mm wide at median part of body, including parapodia and excluding chaetae. Body long, very slender and tapered at posterior end. Prostomium elongated and clearly longer than broad, antennae and palps inserted on a well-developed anterior protuberance (Figure 2A–C). Antennae and palps of similar size, conical and slender. Antennae and palps as long as prostomium. One pair of eyes at posterior margin of prostomium (Figure 2A). Undivided proboscis, diffusely covered with conical papillae (Figure 2C). Terminal ring of proboscis with larger, conical papillae (Figure 2C). First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophores and long cirrostyles; ventral tentacular cirri of segment 2 cylindrical, with inflated base and tapered distal end. First pair of tentacular cirri reaching segment 4. Dorsal and ventral tentacular cirri of segment 2 reaching segments 5 and 2, respectively (Figure 2A–C). Neuropodia and ventral cirri from segment 3. Dorsal cirri absent on segment 3. Normal dorsal cirri with well-developed cirrophores without dorsal extensions, from segment 4, symmetrical and cylindrical, anterior ones more slender than other dorsal cirri and 1.5 times as long as lobes. Dorsal cirri of median segments robust and longer than dorsal cirri of posterior segments. Neuropodia symmetrical, clearly shorter than ventral cirri, with light-brown acicula. Prechaetal lobes bilobate, symmetrical and rounded. Postchaetal lobes rounded. Ventral cirri horizontally oriented in relation to lobes, with inflated bases and tapered distal ends on anterior parapodia; median ventral cirri longer and more slender; more robust and shorter on posterior segments (Figures 6A, B & 7A–C). A similar pattern of cirri development and arrangement was also observed in paratypes (Figure 8A, B). Two supracicular and two subacicular spinigerous compound chaetae from segment 3. Rostrum of three uppermost chaetae bidentate and of lowermost one unidentate. Supracicular chaetae bidentate and subacicular chaetae unidentate in posterior parapodia, articles of varied shape and degrees of serration (Figure 9A–D). Pygidium with one pair of long, slender, cylindrical anal cirri; pygidial papilla not observed (Figure 7D).
COLOUR
Living specimens with bright green pigmentation. Mature animals show dark green pigmentation on the posterior segments (Figure 2C). When preserved, animals have an opaque pigmentation with brown parapodial cirri.
DISTRIBUTION
Atlantic Ocean, Brazil: Beaches in Santa Catarina, Rio de Janeiro and Bahia. Pacific Ocean: Valparaiso – Chile.
REMARKS
The morphological characteristics of the material from the Brazilian coast coincide well with the paratypes deposited in Valparaiso, Chile. However, the phylogenetic affinities between populations from the Atlantic and Pacific coasts of South America can only be confirmed through molecular analyses. Hesionura laubieri is the only species previously reported from Brazil (Westheide, Reference Westheide1974). Most of the regional reports probably refer to some of the other new species described herein. It differs from Hesionura ceciliae sp. nov. in having dorsal and ventral cirri longer than the parapodial lobes. The dorsal and ventral cirri of H. ceciliae sp. nov. are usually dark-brown, in contrast with those in H. laubieri, which are lighter in preserved animals. Hesionura laubieri differs from H. biformata sp. nov., as the ventral cirri in the latter are abruptly tapered in posterior parapodia. The dorsal and ventral cirri of H. laubieri are cylindrical and longer than the chaetae. They are cylindrical with inflated bases in Hesionura variodentata sp. nov. and cylindrical with round distal ends in H. carioca sp. nov. Hesionura laubieri shares long and slender parapodial cirri with H. augeneri (Friedrich, Reference Friedrich1937) and H. australiensis Hartmann-Schröder & Parker, Reference Hartmann-Schröder and Parker1990, but it differs from them by the number of teeth on the rostrum of chaetal shafts. It shares bidentate supracicular chaetae with H. ceciliae sp. nov. and H. serrata (Hartmann-Schröder, Reference Hartmann-Schröder1960), as well as four chaetae per parapodium with H. australiensis and H. variodentata sp. nov. It differs from Hesionura elongata (Southern, Reference Southern1914) by the absence of simple chaetae and the length of the parapodial cirri, which are half as long in H. laubieri. The absence of a pygidial papilla separates Hesionura laubieri from H. biformata sp. nov., H. variodentata sp. nov., H. lupina sp. nov. and H. shandongensis Zhao & Wu, Reference Zhao and Wu1991. All the species of Hesionura described herein share an elongated prostomium and long, cylindrical anal cirri.
Hesionura ceciliae sp. nov.
(Figures 10–12)

Fig. 10. Hesionura ceciliae sp. nov.: (A) Prostomium and anterior end of holotype, dorsal view; (B) Ventral tentacular cirrus of holotype (arrow). Scale bars: 0.05 mm.

Fig. 11. Hesionura ceciliae sp. nov. Dorsal views of parapodia from the holotype: (A) Parapodium 6, with dorsal cirrus (arrow); (B) Parapodium 7, ventral cirri (arrow); (C) Parapodium 30, dorsal cirri (arrow); (D) Parapodium 30, ventral cirrus (arrow); (E) Posterior view of median parapodium. Scale bars: 0.05 mm.
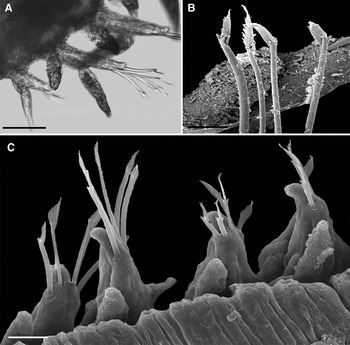
Fig. 12. Hesionura ceciliae sp. nov. Dorsal views: (A) Parapodium 59; (B) Bidentate chaetae from anterior parapodium; (C) Compound spinigerous chaetae in median parapodia. Scale bars: A, 0.05 mm; B, 0.006 mm; C, 0.01 mm.
TYPE MATERIAL
Holotype. Margin of continental shelf in Campos Basin, Brazil, Hab13 H04 R1, 21°42′53.723″S 40°10′15.249″W, 98 m; coarse sandy bottom, 9 March 2009 (ZUEC–POL–16876).
Paratypes. 40 paratypes, length from 1.0 to 4.0 mm and number of segments from 27 to 96. Continental shelf in Campos Basin, Brazil: Hab13 I01 R2, 21°11′0.839″S 40°28′27.312″W, 26 m, 5 March 2009 (9 paratypes, ZUEC–POL–16883); Hab13 Foz16 R1, 21°12′14.127″S 40°42′26.060″W, 15 m, 7 March 2009 (3 paratypes, ZUEC–POL–17011); Hab13 Foz01 R2, 21°17′25.307″S 40°54′7.308″W, 15 m, 8 March 2009 (2 paratypes, ZUEC–POL–16898); Hab13 H02 R3, 21°44′18.9″S, 40°17′14.4″W, 49 m, 9 March 2009 (1 paratype, ZMUC–POL–2406); Hab13 Foz18 R2, 21°33′54.457″S 40°42′55.682″W, 22 m, 10 March 2009 (3 paratype, ZUEC–POL–17008); Hab13 Foz18 R3, 21°33′53.089″S 40°42′55.610″W, 21 m, 10 March 2009 (1 paratype, ZUEC–POL–17007); Hab13 Foz19 R1, 21°44′42.879″S, 40°43′9.193″W, 22 m, 11 March 2009 (1 paratype, ZMUC–POL–2407); Hab13 Foz30 R2, 21°34′12.572″S 40°25′32.411″W, 29 m, 13 March 2009 (1 paratype, ZUEC–POL–16875); Hab13 Foz43 R1, 22°12′18.796″S 40°14′39.451″W, 97 m, 15 March 2009 (2 paratypes, ZUEC–POL–16921); Hab16 H02 R2, 21°44′19.591″S 40°17′15.669″W, 50 m, 8 July 2009 (1 paratype, ZUEC–POL–17010); Hab16 H02 R3, 21°44′19.6″S 40°17′15.608″W, 49 m, 9 July 2009 (1 paratype, ZUEC–POL–16877); Hab17 Foz29 R01, 21°24′45.072″S 40°25′17.287″W, 32 m, 20 July 2009 (1 paratype, ZUEC–POL–16884); Hab17 Foz18 R3, 21°33′54.6″S, 40°2′53.9″W, 23 m, 20 July 2009 (3 paratypes, ZMUC–POL–2408); Hab17 Foz18 R2, 21°33′54.232″S, 40°42′55.434″W, 23 m, 20 July 2009 (1 paratype, ZMUC–POL–2409); Hab17 I01 R2, 21°11′1.370″S 40°28′29.103″W, 26 m, 21 July 2009 (3 paratypes, ZUEC–POL–16957); Hab17 I01 R3, 21°11′1.081″S 40°28′28.858″W, 26 m, 21 July 2009 (1 paratype, ZUEC–POL–16916); Hab17 Foz16 R1, 21°12′15.019″S 40°42′26.207″W, 16 m, 22 July 2009 (1 paratype, ZUEC–POL–16903); Hab17 Foz24 R2, 21°50′20.997″S 40°31′38.672″W, 28 m, 23 July 2009 (2 paratypes, ZUEC–POL–17006); Hab17 Foz30 R2, 21°34′13.655″S 40°25′31.571″W, 28 m, 23 July 2009 (1 paratype, ZUEC–POL–17009); Hab17 Foz23 R2, 21°01′9.4″S 40°31′55.3″W, 24 July 2009 (2 paratypes, ZMUC–POL–2410). Collected on substrates of coarse sand, between 22 and 98 m.
DIAGNOSIS
Ventral cirri on anterior and median segments conical, with tapered distal ends. Four bidentate chaetae throughout the body, except in the last segments, which have five chaetae of which the lowermost one is unidentate.
DESCRIPTION
Holotype a complete specimen with 57 segments, 3.0 mm in length, measuring 0.1 mm wide on median part of body including parapodia and excluding chaetae. Body long, very slender and tapered at posterior end. Prostomium elongated, much longer than wide, with antennae and palps inserted on a well-outlined anterior protuberance. Antennae and palps conical and slender, of similar size, longer than prostomium. One pair of eyes at posterior margin of prostomium (Figure 10A). Undivided proboscis, covered with diffusely distributed conical papillae. Terminal ring not observed. First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophores and long cirrostyles; ventral tentacular cirri of segment 2 flat and oval, with tapered distal end (Figure 10A). Tentacular cirri of first segment reaching setiger 4. Dorsal and ventral tentacular cirri of second segment reaching segments 5 and 2, respectively. Neuropodia and ventral cirri from third segment. Dorsal cirri absent on segment 3. Dorsal cirri symmetrical, with well-developed cirrophores, without dorsal extensions, present from segment 4. Dorsal cirri of anterior and median parapodia oval, cylindrical and slender on posterior segments. Lobes longer than dorsal cirri and shorter than ventral cirri. Prechaetal lobes bilobate, symmetrical and rounded. Postchaetal lobes rounded. Ventral cirri horizontally oriented in relation to lobes, from segment 3, conical, slender, with inflated bases on anterior segments; conical on median segments; cylindrical and slender on posterior segments (Figures 11A–D and 12A). Compound spinigerous chaetae from segment 3. Anterior and median parapodia with two supracicular and two subacicular bidentate chaetae. Posterior parapodia with five chaetae, two supracicular and three subacicular; four dorsalmost chaetae bidentate, lowermost chaeta unidentate. Articles of varied shape and with serrated outer edges (Figure 12B, C). Pygidium with one pair of cylindrical and slender anal cirri; pygidial papilla not observed.
COLOUR
Preserved specimens show a brownish pigmentation, more evident in parapodial cirri.
ETYMOLOGY
The species is named after Dr Cecília Amaral, one of the greatest contributors to the field of polychaete taxonomy in Brazil.
DISTRIBUTION
Atlantic Ocean, Brazil: Margin of continental shelf in Campos Basin.
REMARKS
Hesionura ceciliae sp. nov. shares the shape of the ventral tentacular cirri, oval with tapered distal ends, with H. australiensis. It differs from H. laubieri by the oval dorsal cirri and conical ventral cirri, with tapered distal ends, these being much shorter than the chaetae, whereas H. laubieri has cylindrical ventral cirri that are longer than the chaetae. It differs from H. elongata by the absence of simple chaetae and the shape of the parapodial cirri, which are cylindrical in the latter. It shares conical dorsal cirri with H. coineaui difficilis (Banse, Reference Banse1963). Hesionura ceciliae sp. nov. differs from H. biformata sp. nov. and H. australiensis, both of which have long, conical ventral cirri. The shape of the dorsal cirri and the presence of a bidentate chaeta in the uppermost position on anterior and medium parapodia separates Hesionura ceciliae sp. nov from H. variodentata sp. nov., H. carioca sp. nov., H. lupina sp. nov., H. coineaui (Laubier, Reference Laubier1962), H. shandongensis Zhao & Wu, Reference Zhao and Wu1991, H. augeneri (Friedrich, Reference Friedrich1937) and H. australiensis. Hesionura ceciliae sp. nov. shares the presence of five chaetae in posterior parapodia with H. biformata sp. nov., H. carioca sp. nov., H. longissima sp. nov., H. lupina sp. nov. and H. australiensis. Hesionura ceciliae sp. nov. differs from H. biformata sp. nov., H. variodentata sp. nov., H. lupina sp. nov. and H. shandongensis by the absence of a pygidial papilla.
Hesionura biformata sp. nov.
(Figures 13–16)

Fig. 13. Hesionura biformata sp. nov.: (A) Prostomium and anterior end of holotype, dorsal view; (B) Ventral view of partially everted proboscis in the holotype; ventral tentacular cirrus indicated by arrow. Scale bars: 0.05 mm.

Fig. 14. Hesionura biformata sp. nov. Dorsal view of parapodia from the holotype: (A) Parapodium 5, ventral and dorsal cirrus (arrows); (B) Parapodium 25, dorsal cirrus (arrow); (C) Parapodium 25, ventral cirrus (arrow); (D) Parapodium 45, dorsal cirrus (arrow); (E) Parapodium 47, ventral cirrus (arrow); (F) Posterior view of median parapodium. Scale bars: 0.05 mm.

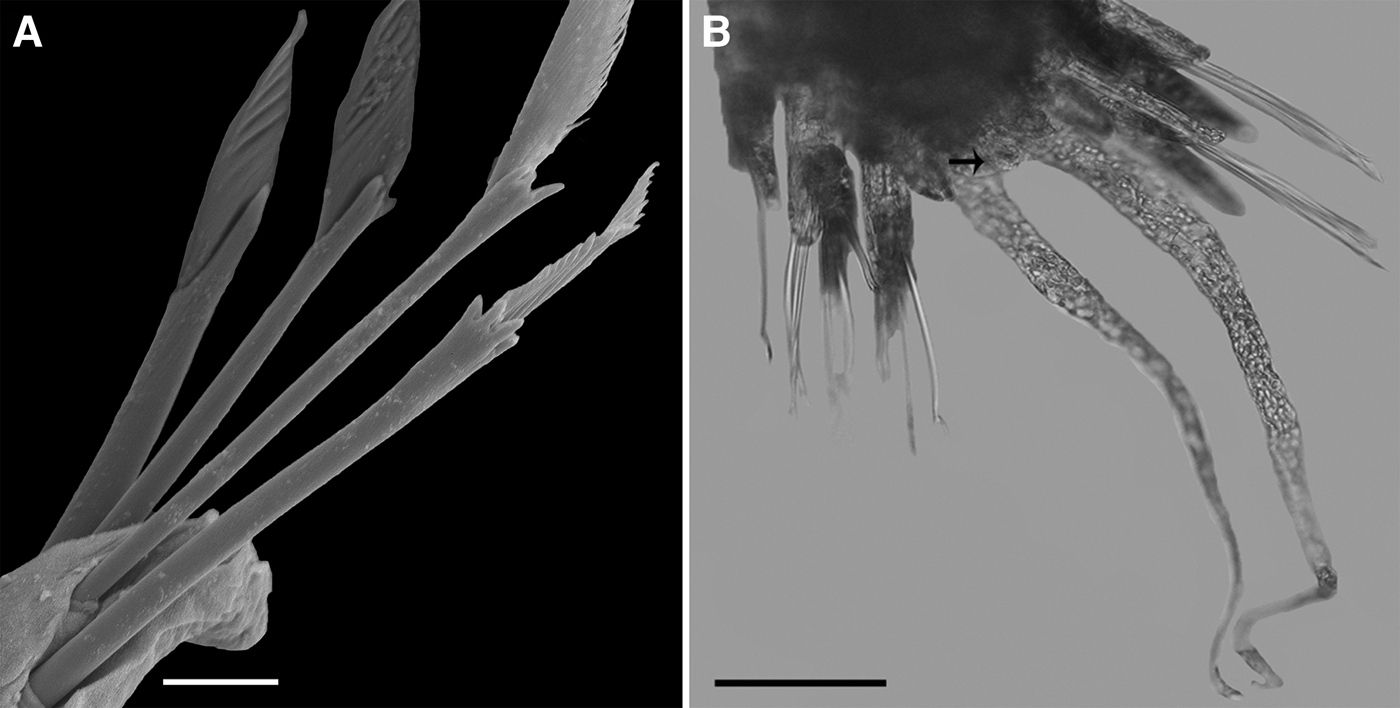
Fig. 15. Hesionura biformata sp. nov.: (A) Anterior parapodia, dorsal view, showing tridentate uppermost chaeta in each neuropodium, other chaetae bidentate; (B, C) Median parapodia, dorsal views, showing tridentate uppermost chaeta, two bidentate middle chaetae, and lowermost chaeta with one rounded tooth; (D) End of shaft and article of tridentate chaeta; (E) End of shaft and article of unidentate chaeta. Scale bars: A–B, 0.01 mm; C, 0.03 mm; D, 0.01 mm, E, 0.01 mm.

Fig. 16. Hesionura biformata sp. nov. Posterior end with anal cirri and pygidial papilla (arrow), ventral view. Scale bar: 0.05 mm.
TYPE MATERIAL
Holotype. Margin of continental shelf in Campos Basin, Brazil, Hab13 Foz13 R2, 21°33′54.457″S 42°42′55.682″W, 21 m, sandy bottom, 10 March 2009 (ZUEC–POL–17028).
Paratypes. 20 paratypes, length from 2.0 to 3.2 mm and number of segments from 45 to 47. Continental shelf in Campos Basin, Brazil: Hab11 G2 R3, 21°59′3.751″S 40°25′10.170″W, 52 m, 25 February 2009 (1 paratype, ZUEC–POL–17339); Hab11 D02 R1, 22°12′52.897″S 40°51′12.067″W, 52 m, 26 February 2009 (2 paratypes, ZUEC–POL–17023); Hab11 D02 R3, 22°12′53.401″S 40°51′12.488″W, 52 m, 26 February 2009 (1 paratype, ZUEC–POL–17033); Hab11 F01 R3, 21°57′16.168″S 40°37′59.616″W, 26 m, 26 February 2009 (1 paratype, ZUEC–POL–17034); Hab13 H02 R2, 21°44′19.348″S 40°17′15.581″W, 49 m, 9 March 2009 (3 paratypes, ZUEC–POL–17031); Hab13 Foz21 R1, 22°6′21.254″S 40°43′39.365″W, 47 m, 12 March 2009 (2 paratypes, ZUEC–POL–17025); Hab13 Foz21 R1, 22°6′21.254″S 40°43′39.365″W, 47 m, 12 March 2009 (1 paratype, ZMUC–POL–2411); Hab13 Foz21 R3, 22°6′21.902″S 40°43′39.494″W, 47 m, 12 March 2009 (1 paratype, ZUEC–POL–17032); Hab16 F02 R2, 22°3′41.650″S 40°24′9.684″W, 55 m, 6 July 2009 (3 paratypes, ZUEC–POL–17024); Hab16 F02 R3, 22°3′41.650″S 40°24′9.944″W, 55 m, 6 July 2009 (2 paratypes, ZUEC–POL–17029); Hab17 Foz10 R2, 21°55′25.779″S 40°49′12.029″W, 21 m, 18 July 2009 (1 paratype, ZUEC–POL–17030); Hab17 Foz10 R3, 21°55′25.907″S 40°49′12.706″W, 21 m, 18 July 2009 (1 paratypes, ZUEC–POL–17026); Hab17 Foz30 R3, 21°34′12.552″S 40°25′32.724″W, 28 m, 23 July 2009 (1 paratype, ZUEC–POL–17022). Collected in sandy substrate, between 21 and 55 m.
DIAGNOSIS
Ventral cirri on median parapodia more than 1.5 times as long as the neuropodial lobes, abruptly tapering on posterior segments. Posterior parapodia with one bidentate and two unidentate subacicular chaetae.
DESCRIPTION
Holotype a complete specimen with 65 segments, 3.0 mm in length, measuring 0.1 mm wide at median part of body, including parapodia and excluding chaetae. Body long, very slender and tapered at posterior end. Prostomium elongated, much longer than wide, with antennae and palps inserted on a well-defined anterior protuberance. Antennae and palps conical and slender, of similar size, half as long as prostomium. One pair of eyes at posterior margin of prostomium (Figure 13A). Undivided proboscis covered with diffusely distributed conical papillae. Terminal ring not observed (Figure 13B). First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophore and long cirrostyle (Figure 13A); ventral tentacular cirri of segment 2 with inflated base, tapering at distal end (Figure 13B). Tentacular cirri of first segment reaching setiger 4. Dorsal and ventral tentacular cirri of segment 2 reaching segments 5 and 2, respectively (Figure 13A). Neuropodia and ventral cirri present from third segment. Dorsal cirri absent on segment 3. Dorsal cirri symmetrical, with well-developed cirrophores, without dorsal extensions, from segment 4. Dorsal cirri of anterior parapodia cylindrical, with rounded distal ends on median segments and oval on posterior segments. Parapodial lobes longer than dorsal cirri and shorter than ventral cirri, with light-brown acicula. Prechaetal lobes bilobate and symmetrical. Postchaetal lobes rounded. Ventral cirri horizontally oriented in relation to lobes, from segment 3, cylindrical on anterior and median segments, slender and abruptly tapered on posterior segments (Figure 14A–E). Compound spinigerous chaetae from segment 3. Rostrum of chaetal shaft with one to three teeth. Anterior parapodia with two supracicular and two subacicular chaetae, uppermost one tridentate, the others bidentate. Median parapodia with most dorsal chaeta tridentate, second and third chaetae bidentate, and lowermost one unidentate. Posterior parapodia with two supracicular and three subacicular chaetae, three uppermost ones bidentate, two ventralmost ones unidentate; article with serrated outer edge (Figure 15A–D). Pygidium with one pair of cylindrical anal cirri; pygidial papilla present (Figure 16).
COLOUR
Preserved specimens are brownish, with pigment more concentrated on the dorsal cirri and at the base of the ventral cirri.
DISTRIBUTION
Atlantic Ocean, Brazil: Margin of continental shelf in Campos Basin.
ETYMOLOGY
The name of this species means ‘of two shapes’, specifically referring to the cylindrical ventral cirri that become abruptly filiform in posterior segments.
REMARKS
Hesionura biformata sp. nov. differs from H. laubieri and H. elongata by the length of the ventral cirri, which although long do not exceed the length of the chaetae, and by the presence of five chaetae in posterior parapodia, with the three dorsalmost ones being bidentate and the two ventralmost ones being unidentate. Hesionura biformata sp. nov. shares the oval shape of the dorsal cirri on posterior segments with that in H. carioca sp. nov. and H. mystidoides Hartmann-Schröder, Reference Hartmann-Schröder1963. It differs from H. variodentata sp. nov., H. lupina sp. nov. and H. shandongensis by having median ventral cirri cylindrical and slender, abruptly tapered on posterior segments. It shares the presence of cylindrical and slender ventral cirri on anterior segments with Hesionura fragilis Hartmann-Schröder, Reference Hartmann-Schröder1958. The presence of cylindrical dorsal cirri with distal rounded ends on anterior parapodia and the number of teeth on the chaetal rostra separates H. biformata sp. nov. from H. ceciliae sp. nov. As in H. variodentata sp. nov., H. carioca sp. nov., H. lupina sp. nov. and H. australiensis, the uppermost chaeta is tridentate on anterior and median parapodia.
Hesionura variodentata sp. nov.
(Figures 17–19)

Fig. 17. Hesionura variodentata sp. nov. Prostomium and anterior end of holotype, ventral view; tentacular cirri of segment 1 and ventral tentacular cirri of segment 2 (arrow) in focus. Scale bar: 0.04 mm.

Fig. 18. Hesionura variodentata sp. nov. Dorsal view of parapodia of the holotype: (A) Parapodium 7, dorsal cirrus (arrow); (B) Parapodium 7, ventral cirrus (arrow); (C) Parapodium 35, dorsal cirrus (arrow); (D) Parapodium 35, cylindrical ventral cirrus (arrow); (E) Parapodium 70, dorsal cirrus (arrow); (F) Parapodium 67, ventral cirrus (arrow); (G) Posterior view of median parapodium. Scale bars: A–D; G, 0.05 mm; E, 0.02 mm; F, 0.03 mm.

Fig. 19. Hesionura variodentata sp. nov.: (A) Dorsalmost chaeta from anterior parapodium; (B) Four compound spinigerous chaetae from posterior parapodium; (C) Pygidium with anal cirri and scar from pygidial papilla, dorsal view. Scale bars: A–B, 0.02 mm; C, 0.01 mm.
TYPE MATERIAL
Holotype. Margin of continental shelf in Campos Basin, Brazil, Hab13 I01 R2, 21°11′0.839″S 40°28′27.312″W, 26 m, 5 March 2009 (ZUEC–POL–17035).
Paratypes. A total of 15 paratypes, length from 1.2 to 4.5 mm and number of segments from 47 to 94. Continental shelf in Campos Basin, Brazil: Hab13 I01 R2, 21°11′0.839″S 40°28′27.312″W, 26 m, 5 March 2009 (4 paratypes, ZUEC–POL–17036); Hab13 H01 R2, 21°43′22.206″S 40°31′52.366″W, 26 m, 9 March 2009 (2 paratypes, ZMUC–POL–2412), Hab13 B01 R3, 22°41′47.098″S 41°53′46.452″W, 30 m, 16 March 2009 (7 paratypes, ZUEC–POL–17037); Hab17 Foz16 R1, 21°12′15.019″S 40°42′26.207″W, 16 m, 22 July 2009 (1 paratype, ZUEC–POL–17038); Hab17 Foz16 R3, 21°12′14.606″S 40°42′27.627″W, 15 m, 22 July 2009 (1 paratype, ZUEC–POL–17039). Collected in sandy substrates between 15 and 30 m.
DIAGNOSIS
Ventral tentacular cirri conical. Anterior and median parapodia with uppermost chaeta tridentate and other three chaetae bidentate. Posterior parapodia with four chaetae, the three dorsalmost bidentate, the ventralmost one unidentate.
DESCRIPTION
Holotype a complete specimen with 65 segments, 4.5 mm long and 0.08 mm wide in median part of body, including parapodia and excluding chaetae. Body long, very slender and tapered at posterior end. Prostomium elongated, longer than wide, with antennae and palps inserted on a well-defined anterior protuberance (Figure 17). Antennae and palps cylindrical and slender, of similar size, half as long as prostomium. One pair of eyes at posterior margin of prostomium (Figure 17). Undivided proboscis covered with diffusely distributed conical papillae. Terminal ring not observed. First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophore and long cirrostyle; ventral tentacular cirri of segment 2 with inflated base and distally tapered. Tentacular cirri of first segment reaching segment 4. Dorsal and ventral tentacular cirri of segment 2 reaching segments 5 and 3, respectively. Neuropodia and ventral cirri preset from third segment. Dorsal cirri absent on segment 3. Dorsal cirri symmetrical, with well-developed oval cirrophore, without dorsal extension, from segment 4. Dorsal cirri oval, on median segments longer than on anterior and posterior segments. Parapodial lobes symmetrical, longer than dorsal cirri and shorter than ventral cirri, with light-brown acicula. Prechaetal lobe bilobate, symmetrical and rounded. Postchaetal lobe rounded. Ventral cirri horizontally oriented in relation to lobes, present from segment 3, conical on anterior segments, cylindrical with inflated base on median segments, elongate on posterior segments (Figure 18A–D). Compound spinigerous chaetae from segment 3. Rostrum of chaetal shaft with one to three teeth. All neuropodia with two supracicular and two subacicular chaetae; dorsalmost chaeta tridentate, the other three bidentate on anterior and median segments; on posterior segments three dorsalmost chaetae bidentate, lowermost chaeta unidentate. Articles of varied shape, with serrated outer edge (Figure 19A–B). Pygidium with one pair of cylindrical anal cirri. Pygidial papilla present (Figure 19C).
COLOUR
Preserved specimens are opaque.
DISTRIBUTION
Atlantic Ocean, Brazil: Margin of continental shelf in Campos Basin.
ETYMOLOGY
The name of this species refers to the varying dentition patterns in chaetae from posterior parapodia.
REMARKS
The presence of a median pygidial papilla has previously been reported only for Hesionura shandongensis, but it is herein also described in H. variodentata sp. nov., H. biformata sp. nov., H. carioca sp. nov. and H. lupina sp. nov. Hesionura variodentata sp. nov. differs from H. laubieri, H. augeneri and H. australiensis in the shape of the parapodial cirri. As in H. laubieri and in H. longissima sp. nov., only four chaetae are found in all neuropodia. Hesionura variodentata sp. nov. differs from H. elongata by the shape of the ventral cirri and by the absence of simple chaetae, as well as the number of teeth on the chaetal rostra, a character which also differentiates it from H. ceciliae sp. nov. It shares the presence of a tridentate dorsalmost supracicular chaeta in anterior and median parapodia with H. biformata sp. nov., H. carioca sp. nov., H. lupina sp. nov. and H. australiensis, but differs from them in having only four chaetae in posterior parapodia. Hesionura variodentata sp. nov. differs from H. longissima sp. nov. in the shape of the parapodial cirri and in the number of chaetal teeth in anterior and median parapodia (see Table 1).
Hesionura carioca sp. nov.
(Figures 20–22)

Fig. 20. Hesionura carioca sp. nov. (A) Anterior end, anterolateral view showing tentacular cirri of right side of body (ventral tentacular cirrus of segment 2 indicataed by arrow); (B) Modified ventral cirri in mature specimen. Scale bars: 0.05 mm.

Fig. 21. Hesionura carioca sp. nov. Dorsal view of parapodia of the holotype: (A) Anterior parapodia, dorsal cirri oval (arrow); (B) Anterior parapodia, ventral cirri oval (arrow); (C, D) Median and posterior parapodia, dorsal cirri oval (arrow); (E) Posterior view of median parapodium. Scale bars: 0.05 mm.

Fig. 22. Hesionura carioca sp. nov.: (A) Parapodia with oval cirri, holotype; (B) Chaetae of anterior parapodium; (C) Pygidium with pair of anal cirri, holotype. Scale bars: A, 0.03 mm; B, 0.05 mm; C, 0.5 mm.
TYPE MATERIAL
Holotype. Margin of continental shelf in Campos Basin, Brazil, Hab17 Foz16 R1, 21°12′15.019″S 40°42′26.207″W, 16 m, sandy bottom, 22 July 2009 (ZUEC–POL–17040).
Paratypes. 7 paratypes, length from 4.0 to 6.0 mm and number of segments from 70 to 90. Continental shelf in Campos Basin, Brazil: Hab13 Foz13 R2, 21°33′54.457″S 40°42′55.682″W, 21 m, 10 March 2009 (4 paratypes, ZUEC–POL–17042); Hab17 Foz18 R3, 21°33′54.611″S 40°42′53.901″W, 23 m, 20 July 2009 (1 paratype, ZUEC–POL–17041); Hab13 Foz18 R3, 21°33′54.611″S, 40°42′53.901″W, 23 m, 20 July 2009 (2 paratypes, ZMUC–POL–2413). Collected in sandy substrate, 21–23 m.
DIAGNOSIS
Ventral tentacular cirri cylindrical and relatively elongate with tapered distal end. Dorsal cirri oval. Posterior parapodia with one tridentate, three bidentate and one unidentate chaetae.
DESCRIPTION
Holotype a complete specimen with 68 segments, 5.0 mm long, and 0.09 mm wide at median part of body, including parapodia and excluding chaetae. Body long, very slender and tapered at posterior end. Prostomium elongated, longer than wide, antennae and palps inserted on well-delineated anterior protuberance. Antennae and palps conical and slender, of similar size, about as long as prostomium. One pair of eyes situated at posterior margin of prostomium. Undivided proboscis covered with diffusely distributed, conical papillae. Terminal ring of proboscis with conical papillae. First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophore and long cirrostyle; ventral tentacular cirri of segment 2 with inflated base, distally tapered. First pair of tentacular cirri reaching segment 3. Dorsal and ventral tentacular cirri of segment 2 reaching segments 3 and 5, respectively (Figure 20A, B). Neuropodia and ventral cirri present from segment 3. Dorsal cirri absent on segment 3. Dorsal cirri symmetrical and oval, with well-developed cirrophores, without dorsal extensions, from segment 4. Dorsal cirri more elongate on anterior neuropodia, more robust on median neuropodia, and more rounded on posterior segments. Parapodial lobes longer than dorsal and shorter than ventral cirri, with light-brown acicula. Prechaetal lobes bilobate, rounded. Postchaetal lobes rounded. Ventral cirri horizontally oriented in relation to lobes from segment 3 on, with round distal end, on anterior segments cylindrical, on median segments cylindrical and slender, on posterior segments more rounded (Figure 21A–D). The parapodial cirri modified to rounded during the reproductive stage (Figure 21D). Compound spinigerous chaetae from segment 3. Rostrum of chaetal shaft with one to three teeth. Anterior and median parapodia with two supracicular and two subacicular chaetae, dorsalmost chaeta tridentate, the other three bidentate. Posterior parapodia with five chaetae, two supracicular and three subacicular, dorsalmost chaeta tridentate, lowermost chaeta unidentate, the three others bidentate. Articles of varied shape, with serrated outer edge (Figure 22A). Pygidium with one pair of long cylindrical anal cirri. Pygidial papilla present (Figure 22B).
COLOUR
Preserved specimens are opaque.
DISTRIBUTION
Atlantic Ocean, Brazil: margin of the continental shelf in Campos Basin.
ETYMOLOGY
The species name honours the people from the beautiful city of Rio de Janeiro.
REMARKS
Hesionura carioca sp. nov., and H. longissima sp. nov. share the rounded shape of the parapodial cirri, modified during the reproductive stage. Hesionura carioca sp. nov. differs from H. laubieri and H. elongata by the presence of long, cylindrical ventral tentacular cirri, the shape and length of the dorsal and ventral cirri, as well as by the number of teeth on the rostrum of chaetae in anterior and medium parapodia. As in Hesionura biformata sp. nov., H. variodentata sp. nov., H. lupina sp. nov. and H. coineaui, the dorsalmost chaeta is tridentate on anterior and median neuropodia. Hesionura carioca sp. nov. shares the presence of a pygidial papilla with H. variodentata sp. nov., H. biformata sp. nov., H. lupina sp. nov. and H. shandongensis. Hesionura carioca sp. nov. differs from H. ceciliae sp. nov. in the shape of the parapodial cirri and in the number of teeth on the dorsalmost chaeta throughout the body (tridentate and bidentate, respectively).
Hesionura longissima sp. nov.
(Figures 23–26)

Fig. 23. Hesionura longissima sp. nov.: (A) Anterior end, dorsal view, paratype; (B) Prostomium, dorsal view; (C) Anterior segments, ventral view, note ventral tentacular cirrus at the right; (D) Anterior segments, ventral view, note tentacular cirrus of segment 1 at the left. (B–D), holotype. Scale bars: A, 0.05 mm; B, 0.2 mm; C–D, 0.05 mm.

Fig. 24. Hesionura longissima sp. nov. Holotype: (A) Anterior parapodia, dorsal cirri (arrow); (B) Anterior parapodia, ventral cirri (arrow); (C) Median parapodia, dorsal cirri (arrow); (D–F) Median and posterior parapodia, ventral cirri (arrow). Scale bars: A–F, 0.05 mm.

Fig. 25. Hesionura longissima sp. nov. (A) Anterior parapodia, dorsal cirri; (B) Anterior parapodium, ventral cirrus; (C) Median parapodia, dorsal and ventral cirri; (D) Posterior parapodium, dorsal and ventral cirri. DC, Dorsal cirrus; VC, Ventral cirrus. Scale bars: A, 0.02; B–D, 0.05 mm.

Fig. 26. Hesionura longissima sp. nov. (A) Posterior view of median parapodium; (B) Compound spinigerous chaetae; (C) Quadridentate supracicular compound chaeta. Scale bars: A, 0.05 mm; C–B, 0.01 mm.
TYPE MATERIAL
Holotype. Margin of continental continental shelf in Campos Basin, Brazil, Hab17 Foz25 R1, 21°39′32.551″S 40°31′124.888″W, 28 m, 23 July 2009 (ZUEC–POL–17044).
Paratypes. A total of 53 paratypes, length from 4.0 to 21 mm and number of segments from 80 to 198. Continental shelf in Campos Basin, Brazil: Hab11 F02 R1, 22°3′41.155″S 40°24′9.910″W, 56 m, 25 February 2009 (1 paratype, ZUEC–POL–17049); Hab13 I01 R3, 21°11′0.915″S 40°28′27.114″W, 26 m, 5 March 2009 (17 paratypes, ZUEC–POL–17045); Hab13 H01 R3, 21°43′22.256″S 40°31′52.229″W, 26 m, 9 March 2009 (7 paratypes, ZUEC–POL–17053); Hab13 Foz24 R3, 21°50′20.898″S 40°31′39.721″W, 27 m, 13 March 2009 (10 paratypes, ZUEC–POL–17054); Hab13 Foz31 R1, 21°45′2.021″S 40°25′45.245″W, 26 m, 13 March 2009 (1 paratype, ZMUC–POL–2414); Hab16 Foz31 R3, 21°45′2.157″S 40°25′45.051″W, 25 m, 9 July 2009 (1 paratype, ZUEC–POL–17046); Hab17 Foz03 R1, 21°28′2.071″S 40°56′16.956″W, 15 m, 20 July 2009 (2 paratypes, ZMUC–POL–2415); Hab17 Foz18 R1, 21°33′54.340″S 40°42′55.771″W, 23 m, 20 July 2009 (2 paratypes, ZUEC–POL–17051); Hab17 Foz29 R1, 21°24′45.072″S 40°25′17.287″W, 26 m, 21 July 2009 (1 paratype, ZUEC–POL–17043); Hab17 Foz04 R3, 22°23′22.230″S, 40°34′59.116″W, 110 m, 25 July 2009 (2 paratypes, ZMUC–POL–2416); Hab17 I01 R1, 21°11′0.342″S 40°28′28.313″W, 26 m, 21 July 2009 (1 paratype, ZUEC–POL–17050); Hab17 Foz24 R1, 21°50′21.049″S 40°31′37.376″W, 28 m, 23 July 2009 (5 paratypes, ZUEC–POL–17052); Hab17 Foz24 R2, 21°50′20.997″S 40°31′38.672″W, 28 m, 23 July 2009 (1 paratype, ZUEC–POL–17047); Hab17 Foz30 R2, 21°34′13.655″S 40°25′31.571″W, 28 m, 23 July 2009 (1 paratype, ZUEC–POL–17048); Hab17 D04 R1, 22°23′ 22.230″S 40°34′59.116″W, 110 m, 25 July 2009 (1 paratype, ZMUC–POL–2417). In very thick sandy bottoms, between 15 and 110 m.
DIAGNOSIS
Parapodia with dorsalmost supracicular chaeta quadridentate. Adults with long body, up to 21 mm, usually with about 200 segments. Dorsal and ventral cirri rounded in reproductive stages.
DESCRIPTION
Holotype a complete specimen, with 198 segments and 21 mm in length, 0.29 mm wide at median part of body, including parapodia and excluding chaetae. Body long, very slender and tapered at posterior end. Prostomium elongated, antennae and palps inserted on well-defined anterior protuberance. Antennae and palps conical and slender, of similar size, half as long as prostomium. One pair of eyes on median region of prostomium (Figure 23B). Undivided proboscis covered with diffusely distributed conical papillae. Terminal ring of proboscis with conical papillae (Figure 23D). First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophore and long cirrostyle; ventral tentacular cirri of segment 2 oval to elongate. Tentacular cirri of first segment reaching segment 4. Dorsal and ventral tentacular cirri of segment 2 reaching segments 5 and 3, respectively (Figure 23A). Neuropodia and ventral cirri present from segment 3. Dorsal cirri absent on segment 3. Dorsal cirri symmetrical and oval, with well-developed cirrophores, without dorsal extensions, from segment 4. Median dorsal cirri longer than anterior and posterior ones. In sexually mature animals, dorsal cirri of anterior parapodia oval, on median and posterior segments rounded, median ones more robust than posterior ones. Parapodial lobes longer than dorsal cirri and similar length ventral cirri in reproductive stage, with light-brown acicula. Parapodial lobes longer than dorsal and shorter than ventral cirri in reproductive stage, which is a unique feature in Hesionura. Prechaetal lobes bilobate, rounded. Postchaetal lobes rounded. Ventral cirri symmetrical and oval, horizontally oriented in relation to lobes, present from segment 3, more robust on median segments and thinner on posterior segments (Figures 24A–F & 25A–D). Compound spinigerous chaetae from segment 3. Rostrum of chaetal shaft with one to four teeth. Two supracicular chaetae, dorsalmost one quadridentate, second one tridentate, and two subacicular bidentate chaetae, with exception of last 15 segments, which have the three dorsalmost chaetae bidentate and the ventralmost unidentate. Articles with serrated outer edge (Figure 26A, B). Pygidium with one pair of cylindrical anal cirri; pygidial papilla not observed.
COLOUR
Preserved specimens are opaque.
DISTRIBUTION
Atlantic Ocean, Brazil: Margin of continental shelf in Campos Basin.
ETYMOLOGY
The name of this species refers to the exceptional length of specimens in relation to congeneric species.
REMARKS
Hesionura longissima sp. nov. displays rounded dorsal and ventral tentacular cirri in mature animals. Such modifications are shared with H. carioca sp. nov. (Figures 24C & 25B–D). This type of epitoky was previously unknown for Hesionura. Hesionura longissima differs from other species of the genus by the presence of oval to rounded dorsal and ventral cirri and the presence of a quadridentate chaeta in the dorsalmost position throughout the body, except in the last 20 segments, as well as by the length of the body, which may reach 21 mm in adults. It clearly differs from Hesionura elongata, which has capillary chaetae and fusiform dorsal cirri, and from H. coineaui, H. variodentata sp. nov., H. serrata and H. ceciliae sp. nov. in the shape of the ventral cirri. The presence of five chaetae in the posterior parapodia separates H. longissima sp. nov. from H. laubieri, H. australiensis and H. variodentata sp. nov., all of which have four chaetae in posterior parapodia. Hesionura longissima sp. nov. also differs from H. variodentata sp. nov., H. carioca sp. nov. and H. shandongensis by the absence of a pygidial papilla.
Hesionura lupina sp. nov.
(Figures 27–29)

Fig. 27. Hesionura lupina sp. nov. Anterior end, holotype: (A) Dorsal view of the prostomium; (B) Dorsolateral view, ventral tentacular cirrus visible on left side of animal. Scale bars: A, 0.1 mm; B, 0.07 mm.

Fig. 28. Hesionura lupina sp. nov. Dorsal views of the parapodia in the holotype: (A) Anterior parapodia with dorsal cirri in focus; (B) Anterior parapodia with ventral cirri in focus; (C) Median parapodia with dorsal cirri in focus; (D) Median parapodia with ventral cirri in focus. Scale bars: 0.05 mm.

Fig. 29. Hesionura lupina sp. nov.: (A) Posterior parapodia with dorsal and ventral cirri, dorsal view, holotype; (B) Posterior view of median parapodium (C) Chaetae from posterior parapodium; (D) Anal cirrus, holotype (one anal cirrus lost). Scale bars: A–B, 0.05 mm; C, 0.01 mm; D, 0.2 mm.
TYPE MATERIAL
Holotype. Margin of continental shelf in Campos Basin, Brazil, Hab13 Foz19 R2, 21°44′42.969″S 40°43′8.347″W, 21 m, 11 March 2009 (ZUEC–POL–17061).
Paratypes. 75 paratypes, length from 2.4 to 4.2 mm and number of segments from 42 to 88. Continental shelf in Campos Basin, Brazil: Hab11 D02 R1, 22°12′52.897″S 40°51′12.067″W, 52 m, 26 February 2009 (2 paratypes, ZUEC–POL–17070); Hab13 Foz29 R3, 21°24′43.736″S 40°25′18.618″W, 33 m, 7 March 2009 (1 paratype, ZUEC–POL–17074); Hab13 Foz18 R2, 21°33′54.457″S 40°42′55.682″W, 22 m, 10 March 2009 (5 paratypes, ZUEC–POL–17076); Hab13 Foz18 R2, 21°33′54.457″S 40°42′55.682″W, 22 m, 10 March 2009 (3 paratypes, ZUEC–POL–17073); Hab13 Foz18 R3, 21°33′53.089″S 40°42′55.610″W, 21 m, 10 March 2009 (7 paratypes, ZUEC–POL–17075); Hab13 Foz19 R1, 21°44′42.879″S 40°43′9.193″W, 22 m, 11 March 2009 (4 paratypes, ZUEC–POL–17067); Hab13 Foz10 R2, 21°55′25.160″S 40°49′12.284″W, 21 m, 12 March 2009 (1 paratype, ZUEC–POL–17071); Hab13 Foz30 R3, 21°34′12.597″S 40°25′31.241″W, 29 m, 13 March 2009 (13 paratypes, ZUEC–POL–17068); Hab13 Foz31 R2, 21°45′2.154″S 40°25′44.878″W, 26 m, 13 March 2009 (1 paratype, ZUEC–POL–17062); Hab13 Foz31 R2, 21°45′2.154″S 40°25′44.878″W, 26 m, 13 March 2009 (1 paratype, ZUEC–POL–17065); Hab13 Foz41 R1, 21°45′14.261″S 40°14′7.911″W, 67 m, 14 March 2009 (1 paratype, ZUEC–POL–17064); Hab16 G02 R1, 21°59′3.657″S 40°25′11.070″W, 52 m, 6 July 2009 (1 paratype, ZUEC–POL–17069); Hab16 Foz31 R1, 21°45′2.327″S, 40°25′45.451″W, 27 m, 9 July 2009 (15 paratypes, ZMUC–POL–2418); Hab17 Foz13 R3, 21°28′21.692″S 40°48′34.086″W, 22 m, 20 July 2009 (1 paratype, ZUEC–POL–17078); Hab17 Foz18 R2, 21°33′54.232″S 40°42′55.434.280″W, 23 m, 20 July 2009 (1 paratype, ZUEC–POL–17063); Hab13 Foz27 R3, 21°28′2.200″S 40°31′0.169″W, 30 m, 22 July 2009 (11 paratypes, ZMUC–POL–2419); Hab13 Foz25 R1, 21°39′31.133″S 40°56′21.516″W, 28 m, 23 July 2009 (3 paratypes, ZMUC–POL–2420); Hab17 Foz24 R3, 21°50′20.765″S 40°31′38.459″W, 28 m, 23 July 2009 (1 paratype, ZUEC–POL–17072); Hab17 Foz25 R1, 21°39′32.551″S 40°31′24.888″W, 28 m, 23 July 2009 (3 paratypes, ZUEC–POL–17066). Collected in sandy bottoms, between 21 and 67 m.
DIAGNOSIS
Ventral tentacular cirri of segment 2 cylindrical with inflated base and tapered distal end. Ventral cirri cylindrical and elongate, on median and posterior segments robust, median ones more elongate than posterior ones. Posterior parapodia with one tridentate and four bidentate chaetae.
DESCRIPTION
Holotype a complete female with 134 segments and 4.0 mm in length, 0.03 mm wide at median part of body, including parapodia and excluding chaetae. Body long, cylindrical and tapered at posterior end. Prostomium elongated, antennae and palps inserted on well-defined anterior protuberance. Antennae and palps conical and slender, of similar size, half as long as prostomium. One pair of eyes at posterior margin of prostomium (Figure 27A). Undivided proboscis covered with diffusely distributed, conical papillae. Terminal ring of proboscis with conical papillae. First segment visible dorsally. Tentacular cirri of segment 1 and dorsal tentacular cirri of segment 2 cylindrical, with short cirrophore and long cirrostyle; ventral tentacular cirri of segment 2 with inflated base and tapered distal end. Tentacular cirri of first segment reaching segment 4. Dorsal and ventral tentacular cirri of segment 2 reaching segments 1 and 4, respectively (Figure 27A, B). Neuropodia and ventral cirri present from segment 3. Dorsal cirri symmetrical and cylindrical, with well-developed cirrophores without dorsal extensions, from segment 4, more robust on anterior segments, shorter on median segments, and with more rounded distal end on posterior sements. Parapodial lobes longer than dorsal cirri and shorter than ventral cirri, with light-brown acicula. Prechaetal lobes bilobate and rounded. Postchaetal lobes rounded. Ventral cirri horizontally oriented in relation to parapodial lobes, with rounded distal end, cylindrical and more robust on anterior segments, longer and more slender on posterior segments (Figure 28A–D). Compound spinigerous chaetae from segment 3. Shafts of chaetae with two or three teeth. Anterior and median parapodia with two supracicular and two subacicular chaetae, dorsalmost one tridentate, other three bidentate. Posterior parapodia with five chaetae, two supracicular and three subacicular, dorsalmost chaeta tridentate, other four bidentate; articles with serrated outer edge (Figure 29C). Pygidium with one pair of cylindrical, long anal cirri (Figure 29D). Pygidial papilla present (Figure 29B).
COLOUR
Preserved specimens are opaque.
DISTRIBUTION
Atlantic Ocean, Brazil: Margin of continental shelf in Campos Basin.
ETYMOLOGY
This species is named after our friend and colleague, the late José María Orensanz (affectionately known as Lobo, sea-wolf).
REMARKS
Hesionura lupina sp. differs from H. carioca sp. nov. and H. longissima sp. nov. in the shape of the parapodial cirri, which are rounded in the reproductive stages in the latter. It shares a similar shape of ventral tentacular cirri with H. longissima sp. nov. and H. mystidoides, but differs from them by the cylindrical dorsal cirri. Hesionura lupina sp. nov. shares cylindrical and elongate dorsal and ventral cirri with H. laubieri and H. elongata. Having the uppermost chaeta tridentate in anterior parapodia is a character shared with H. biformata sp. nov., H. variodentata sp. nov., H. carioca sp. nov. and H. coineaui. Hesionura lupina sp. nov., H. biformata sp. nov. and H. carioca sp. nov. have a pygidial papilla similar to that of H. shandongensis. The number of teeth on the chaetal rostrum in anterior and posterior parapodia is similar to those in H. carioca sp. nov., except for the lowermost chaeta in posterior parapodia, which in H. lupina sp. nov. is bidentate and in H. carioca sp. nov. unidentate.
ACKNOWLEDGEMENTS
We thank the Brazilian National Research Council (CNPq), CAPES, Centro de Estudos do Mar from Universidade Federal do Paraná (UFPR), and the Graduate Program in Zoology (UFPR), the Scanning Microscopy Laboratory (Biocel) and Abaline (Association of Boatmen-PR, Paraná), for their financial and logistical support. Fredrik Pleijel of the Tjärnö Marine Biological Laboratory, Gottenburg University, was most helpful to Veronica Oliveira during her stay in Sweden. The first author owes special thanks to Majken Them Tøttrup and Andreas Altenburger (ZMUC), for their attention and support. Our thanks to many colleagues who donated material from the Revizee South Score and Habitat/Petrobrás Projects, and to the Museum of Natural History (Sweden), and the Zoologisches Museum of Hamburg (Germany) for the loan of comparative material.