INTRODUCTION
Pollack, Pollachius pollachius (Linnaeus, 1758) (Gadidae) is distributed widely in the NE Atlantic from northern Norway to North Africa (Cohen et al., Reference Cohen, Inada, Iwamoto and Scialabba1990). Pollachius pollachius is the largest of the eight gadoid species occurring in Galician waters (Bañon et al., Reference Bañon, Villegas-Ríos, Serrano, Mucientes and Arronte2010). This species is targeted mainly by the artisanal fleet, basically with gillnets and lines, in coastal areas, although it is occasionally caught as by-catch in offshore trawl fisheries (Rodriguez et al., Reference Rodriguez, Fariña, Velasco, Pérez and Acosta2011). Unlike other exploited gadoids, such as cod, Gadus morhua and saithe, Pollachius virens, the biology of P. pollachius remains poorly known. Previous reproductive studies were limited in scope and/or focused mainly on controlled experiments for aquaculture purposes (Suquet et al., Reference Suquet, Petton, Normant, Dosdat and Gaignon1996, Reference Suquet, Normant, Gaignon, Quéméner and Fauvel2005; Omnes et al., Reference Omnes, Sévère, Barone, Suquet, Buchet, Le Roux, Gaignon, Fostier, Fauvel, Basurco and Saroglia2002). In spite of its importance as a fishery resource throughout much of its distribution range (ICES, 2010), there is no assessment for P. pollachius in ICES Division VIIIc (where Galician waters are included). In these areas, management conservation is based on total allowable catch (TAC) which are based on landings trends. Recently, P. pollachius was included in a list of stock and species for which collecting data might be required to better evaluate stock status (ICES, 2010; Rodriguez et al., Reference Rodriguez, Fariña, Velasco, Pérez and Acosta2011). As a starting point, biological data are being collected for the species following a European commission decision (European Union, 2010). However, the information available to date is still insufficient to evaluate the P. pollachius stock status in Galician waters.
The capacity of a population to produce viable eggs and larvae is important in the recruitment process of exploited fish stocks (Solemdal, Reference Solemdal1997; Marteinsdottir & Steinarsson, Reference Marteinsdottir and Steinarsson1998; Kraus et al., Reference Kraus, Tomkiewicz and Köster2002). The understanding of these processes has acquired increasing notoriety over the past decade, after the validity of traditional indicators like spawning stock biomass began to be questioned (Marshall et al., Reference Marshall, Kjesbu, Yaragina, Solemdal and Ulltang1998). All these considerations that affect population dynamics are encompassed by the term stock reproductive potential, introduced by Trippel (Reference Trippel1999). Reproductive biology has a crucial role in fishery stock assessment of the productivity of exploited stocks. Its integration into advice and management strategies is expanding through the production of alternative indices of reproductive potential (Trippel, Reference Trippel1999; Morgan, Reference Morgan2008). Thus, improved estimates of population reproductive potential should lead to enhanced performance of the models used in some fisheries assessments (Marshall et al., Reference Marshall, Kjesbu, Yaragina, Solemdal and Ulltang1998). For example, several reproductive traits like sex-ratio, maturity or fecundity have been suggested to be included in sampling programmes for fisheries assessment (Tomkiewicz et al., Reference Tomkiewicz, Morgan, Burnett and Saborido-Rey2003; Lambert, Reference Lambert2008; Morgan, Reference Morgan2008). An accurate evaluation of the gonad development is crucial to describe the maturation process, the timing of reproduction or follicle recruitment process (Murua & Saborido-Rey, Reference Murua and Saborido-Rey2003; Lowerre-Barbieri et al., Reference Lowerre-Barbieri, Ganias, Saborido-Rey, Murua and Hunter2011b). Age/size at maturity is the most common and well-studied life history parameter in fish exploited populations, mainly used in stock assessment to get a better estimation of limit reference points (Morgan, Reference Morgan2008). Additionally, atresia levels have been used as a criterion for determination of fecundity strategies, i.e. determinate vs indeterminate (Greer Walker et al., Reference Greer Walker, Witthames and Bautista de los Santos1994; Alonso-Fernández et al., Reference Alonso-Fernández, Domínguez-Petit, Bao, Rivas and Saborido-Rey2008; Ganias, Reference Ganias2013). The timing and length of the spawning season have long been recognized as critical factors in the recruitment process, since they determine the likelihood of eggs and larvae to find suitable environmental conditions for their survival (Cushing, Reference Cushing1972; Mertz & Myers, Reference Mertz and Myers1994; Alonso-Fernández & Saborido-Rey, Reference Alonso-Fernández and Saborido-Rey2011). Traditionally, studies of reproductive biology have assumed that most of the variations in population reproductive success are attributable to maternal effects (Solemdal, Reference Solemdal1997). Thus, most of the literature on reproductive potential of fish populations has focused mainly on females (Murua & Saborido-Rey, Reference Murua and Saborido-Rey2003). In the last decades, however, there has been an emerging interest in male effects of reproductive potential (Trippel, Reference Trippel2003), the paternal effect, for which a basic knowledge of male reproductive biology is required.
This study aimed to improve our knowledge of the reproductive biology of P. pollachius in an area in the southern part of its geographical range. Several important reproductive traits are described for the first time in a wild population of P. pollachius. Specifically, the objectives of this study were: (1) to describe the gamete development pattern and gonad organization; (2) to characterize the reproductive cycle and prevalence of atresia; and (3) to estimate the length at maturity for males and females in Galician waters. These results will help provide information for any future stock assessment.
MATERIALS AND METHODS
A total of 622 individuals were obtained on a monthly basis from the fish markets along the western coast of Galicia (Atlantic Iberian waters, ICES Division VIIIc, Figure 1) between November 2009 and October 2010, with the exception of August 2010 (Table 1). Fish were kept refrigerated until processing, with tissues sampled within 12 hours.

Fig. 1. Study area, the western coast of Galicia, showing locations of fish markets used to sample Pollachius pollachius.
Table 1. Monthly distribution of the sample size (number of individuals) and mean (±SD) fish total length (cm) and eviscerated weight (g) of Pollachius pollachius from the western coast of Galicia.

Data collection
The following parameters were recorded for each fish: total length (L T, ± 0.1 cm), total weight (W T, ± 0.01 g), eviscerated weight (W E, ± 0.01 g); sex; reproductive phase; gonad weight (W G, ± 0.01 g) and liver weight (W L, ± 0.01 g).
Gonads and livers were removed and three indices were estimated for each mature fish: gonadosomatic index (GSI), hepatosomatic index (HSI) and Fulton's condition factor (K).
 $$\eqalign{& \hbox{GSI} = 100^{*}W_{\rm G}/W_{\rm E}\cr & \hbox{HSI} = 100^{*}W_{\rm L}/W_{\rm E}\cr & {\rm K} = 100^{*}W_{\rm E}/L_{\rm T}^3}$$
$$\eqalign{& \hbox{GSI} = 100^{*}W_{\rm G}/W_{\rm E}\cr & \hbox{HSI} = 100^{*}W_{\rm L}/W_{\rm E}\cr & {\rm K} = 100^{*}W_{\rm E}/L_{\rm T}^3}$$Gonads from all specimens were fixed in 10% formalin, buffered with Na2HPO4*2H2O (0.046 M, molar concentration) and NaH2PO4*H2O (0.029 M) for histological processing. Central portions of the fixed gonads were extracted, dehydrated, embedded in paraffin, sectioned at 3 μm and stained with haematoxylin–eosin for microscopic analysis.
For each ovary, the follicles were classified into stages of development according to histological criteria: primary growth, cortical alveoli, vitellogenesis, germinal vesicle migration and hydration (West, Reference West1990; Tyler & Sumpter, Reference Tyler and Sumpter1996; Brown-Peterson et al., Reference Brown-Peterson, Wyanski, Saborido-Rey, Macewicz and Lowerre-Barbieri2011). Atretic follicles were detected in ovarian histological samples (presence–absence) in order to assess the portion of fish with atresia. Thus, only one of the three indices of atresia proposed by Kurita et al. (Reference Kurita, Meier and Kjesbu2003) was estimated; the prevalence of atresia, as the number of mature female pollack showing follicular atresia divided by the total number of mature females. In male pollack, testicular tissue germ cells were classified into the following stages of development: spermatogonia, spermatocysts, spermatids and spermatozoa (Grier, Reference Grier1981). This allowed each fish, males and females, to be classified in one of the following reproductive phases: immature, developing, spawning capable and correspondent sub-phases actively spawning, regressing and regenerating (Brown-Peterson et al., Reference Brown-Peterson, Wyanski, Saborido-Rey, Macewicz and Lowerre-Barbieri2011).
Cortical alveoli follicles (N = 110) were measured in histological sections of nine females to establish their threshold size, in order to identify the size above which is the stock of developing follicles in the ovaries (µm, mean ± SD = 218 ± 29). To study follicle size–frequency distribution and ovarian development organization of secondary growth follicles, the number and size of developing follicles (follicle development gonadotropin dependent phases: from cortical alveoli to final maturation) (Wallace & Selman, Reference Wallace and Selman1981; Lubzens et al., Reference Lubzens, Young, Bobe and Cerdà2010) were assessed in three actively spawning females. After being separated from the connective tissue (Lowerre-Barbieri & Barbieri, Reference Lowerre-Barbieri and Barbieri1993), follicles were counted and measured in whole mount samples (~50 mg from the central portion of the ovary) with image analysis software (Leica QWin) on a PC connected to a video camera (Leica DFC490) connected to a stereo microscope (Leica Z6 APOA).
Data analysis
All statistical analyses were conducted in R software (R Development Core Team, 2011). Only data from specimens where sex was confirmed histologically were used in the data analysis (304 females, 194 males).
The female reproductive cycle of P. pollachius was evaluated based on the monthly variation of the mean GSI values and frequencies of reproductive phases. Besides the GSI, other somatic indices can be useful to evaluate seasonal fluctuations in energy reserves related to gonadal development and reproductive activity (Henderson et al., Reference Henderson, Wong and Nepszy1996; Lambert & Dutil, Reference Lambert and Dutil2000; Alonso-Fernández & Saborido-Rey, Reference Alonso-Fernández and Saborido-Rey2012). The HSI and K were used as proxies of available energy reserves in liver and somatic tissue, respectively. To reduce the effect of the presence of different reproductive phases in each month and to properly depict the relationship of these indices with the reproductive cycle, the variation in the mean values of somatic indices between reproductive phases was assessed. Differences in GSI, HSI and K during the year and between reproductive phases were statistically evaluated using generalized least squares (GLS) models (Pinheiro et al., Reference Pinheiro, Bates, DebRoy and Sarkar2010) to account for heterogeneity problems (Zuur et al., Reference Zuur, Ieno, Walker, Saveliev and Smith2009).
Length at maturity (L 50%), defined as the body length at which 50% of the individuals were mature, was obtained by using generalized linear models (GLM) (McCullagh & Nelder, Reference McCullagh and Nelder1989) with a binomial family distribution. Sex of the individuals was included as a factor in the model to account for differences in maturation between males and females. L 50% estimations and their uncertainties were estimated employing the non-parametric bootstrap method (Efron, Reference Efron1979). In addition, a graphical comparison of maturity ogives parameters was made using the techniques of Kimura (Reference Kimura1980) for comparison of growth parameters. This technique consisted of generating 95% confidence ellipses around the parameter estimated by the GLM (Murdoch & Chow, Reference Murdoch and Chow2007). Non-overlapping confidence regions indicated differences in maturity ogive parameters.
RESULTS
Ovarian and testicular development organization
The ovaries of Pollachius pollachius are oval-shaped, with the two lobular ovaries partially fused, each one with a roughly hollow cylindrical form and a lamellar internal configuration. Folliculogenesis followed the process of follicle maturation common to other teleosts: primary growth, cortical alveoli, vitellogenesis, migratory nucleus (Wallace & Selman, Reference Wallace and Selman1981; Guraya, Reference Guraya1986; Tyler & Sumpter, Reference Tyler and Sumpter1996). Follicle maturation ends with hydration of follicles, leading then to ovulation (Lowerre-Barbieri et al., Reference Lowerre-Barbieri, Ganias, Saborido-Rey, Murua and Hunter2011b). The nucleus was not displaced by an oil droplet formation.
Pollachius pollachius exhibited an asynchronous development of secondary growth follicles, i.e. secondary growth follicles of all stages of development were present without a dominant population (Figure 2). This asynchrony was reflected by the follicle size–frequency distribution in the spawning capable phase, with no gap among primary growth and developing follicles (Figure 3). There was a continuous follicle size–frequency distribution, with the exception of such ovaries with hydrated follicles, which had a separate mode of large hydrated follicles. Several modes were present within the continuous follicle size–frequency distribution.
Fig. 2. Female reproductive phases of Pollachius pollachius. Follicle development stages and other gonad structures presented in the ovaries: PG, primary growth; CA, cortical alveoli; E-Vit, early vitelogenesis; A-Vit, advanced vitelogenesis; Hyd, hydrated follicle; POF, post ovulatory follicles; At, atretic follicles; Cys, cystic follicles; OW, ovary wall.
Fig. 3. Size–frequency distribution of follicles for three actively spawning females of Pollachius pollachius (total length of females is shown in the legend). Only developing follicles (>200 μm) are represented.
The histological structure of the testis of P. pollachius showed a lobular type. Spermatogonia were distributed randomly along the entire length of the tubule in reproductive active fish. Spermatogenesis occurred within germinal cysts formed by enveloping Sertoli cell cytoplasmic processes; within each cyst, the maturation of germ cells was synchronous (Grier, Reference Grier1980, Reference Grier1981) (Figure 4).
Fig. 4. Male reproductive phases of Pollachius pollachius. Spermatogenesis development stages present in the testis: Sc, spermatocyst; St, spermatid; Sz, spermatozoa; r-Sz, residual spermatozoa.
Reproductive cycle
Values of GSI for both sexes peaked in February. Accordingly, 100% of mature females were classified as spawning capable (including actively spawning) in that month (Figure 5). Reproductive phase frequencies also revealed some reproductive activity in the gonads of females from January to April (spawning capable frequencies, including actively spawning: 26.67% in January, 76.93% in March and 26.92% in April). The largest proportion of regressing females was found in April and May (7.69% and 15.0%, respectively). The prevalence of atresia was highest in April (34.62%) and May (25%), and in regressing individuals (Figure 6). Male reproductive cycle extended from January to May, with a peak of reproductive activity in February. The largest proportion of regressing males was found in June (50%).
Fig. 5. Frequency of reproductive phases for (A) female and (B) male Pollachius pollachius. White line represents variation of mean GSI (gonadosomatic index) value through the year. Numbers on bars represent the number of individuals. X axis labels indicate month and year of sampling.
Fig. 6. Prevalence of atresia in ovaries (A) along the year and (B) in different reproductive phases of Pollachius pollachius. Maturity phases are denoted as follows: Dev, developing; SC, spawning capable; AS, sub-phase actively spawning; Rgs, regressing and Rgn, regenerating. X axis labels indicate month and year of sampling.
Average values of GSI, HSI and K varied significantly during seasonal reproduction and sexes (Table 2). All the three indices dropped to minimum values in March/April in females, extending to May in the case of males (Figure 7). Maximum HSI levels occurred in October for females and November for males. K was at a maximum in November for both sexes.
Fig. 7. Sex-specific monthly and reproductive phases variation of somatic indices of Pollachius pollachius: (A) GSI vs month; (B) GSI vs reproductive phase; (C) HSI vs month; (D) HSI vs reproductive phase; (E) K vs month; and (F) K vs reproductive phase. GSI, gonadosomatic index; HIS, hepatosomoatic index; K, condition factor. Grey triangles and dashed lines represent females and black dots and solid line represent males. X axis labels indicate month and year of sampling.
Table 2. Seasonal variation of somatic indices of Pollachius pollachius from the western coast of Galicia. Summary of multiple general least squares (GLS). GSI, gonadosomatic index; HIS, hepatosomatic index; K, condition factor. Note that the symbol ‘:’ denotes interaction and df means degrees of freedom.

Variation of GSI, HSI and K among the different reproductive phases and sexes was also significant (Table 3). Maximum values of GSI were observed in actively spawning females and males (Figure 7), and minimum values in the regenerating phase. For the HSI, minimum values corresponded with the actively spawning sub-phase in both sexes and maximum values occurred in the developing phase (Figure 7). In the case of K, maximum value was found in the regressing phase for females and in the developing phase for males. The minimum was located in the spawning capable phase for males and in the actively spawning sub-phase in females (Figure 7).
Table 3. Variation of somatic indices in relation to reproductive phase of Pollachius pollachius from the western coast of Galicia. Summary of multiple generalized least squares (GLS). GSI, gonadosomatic index; HIS, hepatosomoatic index; K, condition factor. Note that the symbol ‘:’ denotes interaction and df means degrees of freedom.

Length at sexual maturity
Maturity ogives for males and females displayed a similar shape (Figure 8). Nonetheless, estimates of L50% revealed significant differences between males and females, being 36.1 cm and 47.1 cm, respectively. The smallest mature fish found were a 26.8 cm male and a 42 cm female and the largest immature fish were a 47.3 cm male and a 66 cm female. Variation in maturity ogives was due mainly to the intercept of the relationship (Table 4; Figure 8). L50% for the sexes combined was 42.3 cm.
Fig. 8. Sex-specific and sex-combined plots of maturity ogives for Pollachius pollachius. The 95% confidence elliptic regions for α (intercept) and β (slope) estimations are shown in the secondary panel.
Table 4. Sex-specific and sex-combined maturity ogives of Pollachius pollachius from the western coast of Galicia. Summary of multiple generalized linear models (GLM). Also, L 50% estimation with correspondent bootstrap uncertainty parameters are provided for females, males and sexes combined.

DISCUSSION
Pollachius pollachius is a widely distributed species on European coasts and although there are reports of P. pollachius landings in Spanish waters (Rodriguez et al., Reference Rodriguez, Fariña, Velasco, Pérez and Acosta2011), little biological information is available for this species in the southern parts of its geographical range. Therefore, the present study presents the first data for reproductive biology of this species in Galician waters, which will be informative for any future stock assessment and fishery management (Marshall et al., Reference Marshall, O'Brien, Tomkiewicz, Koster, Kraus, Marteinsdottir, Morgan, Saborido-Rey, Blanchard, Secor, Wright, Mukhina and Bjornsson2003; Morgan, Reference Morgan2008; Lowerre-Barbieri et al., Reference Lowerre-Barbieri, Ganias, Saborido-Rey, Murua and Hunter2011b), but also for aquaculture purposes (Suquet et al., Reference Suquet, Petton, Normant, Dosdat and Gaignon1996; Omnes et al., Reference Omnes, Sévère, Barone, Suquet, Buchet, Le Roux, Gaignon, Fostier, Fauvel, Basurco and Saroglia2002).
Proper knowledge of gametogenesis represents a main goal in comprehending fish reproductive strategies (Murua & Saborido-Rey, Reference Murua and Saborido-Rey2003). Thus, an accurate evaluation of ovarian development is crucial to describe the maturation process, the timing of reproduction or the follicle recruitment (Lowerre-Barbieri et al., Reference Lowerre-Barbieri, Brown-Peterson, Murua, Tomkiewicz, Wyanski and Saborido-Rey2011a). Histological examination of the gonads revealed that P. pollachius presented asynchronous development of secondary growth follicles, i.e. follicles of all stages of development were present without dominant cohorts. This was confirmed by the continuous follicle-size distribution which is a distinct evidence of asynchronous development of secondary growth follicles (Wallace & Selman, Reference Wallace and Selman1981; Murua & Saborido-Rey, Reference Murua and Saborido-Rey2003). Asynchronous development of secondary growth follicles is usually associated with species of indeterminate fecundity (Murua & Saborido-Rey, Reference Murua and Saborido-Rey2003). However, determinate fecundity strategies have been proposed in certain asynchronous species including Atlantic mackerel, Scomber scombrus (Greer Walker et al., Reference Greer Walker, Witthames and Bautista de los Santos1994), brown meager, Sciaena umbra (Grau et al., Reference Grau, Linde and Grau2009) and bib, Trisopterus luscus (Alonso-Fernández et al., Reference Alonso-Fernández, Domínguez-Petit, Bao, Rivas and Saborido-Rey2008). However, further research on follicle recruitment process is required to accurately determine the fecundity type of P. pollachius. Although this study did not address egg production, histological examination of the ovaries indicated that P. pollachius is a batch spawner. Presence of post-ovulatory follicles in the spawning capable females and the presence of several modes in the developing follicle pool are clear evidences of batch spawning type (Murua & Motos, Reference Murua and Motos2006). This fact is in agreement with previous studies of P. pollachius spawning type in captivity (Suquet et al., Reference Suquet, Petton, Normant, Dosdat and Gaignon1996; Omnes et al., Reference Omnes, Sévère, Barone, Suquet, Buchet, Le Roux, Gaignon, Fostier, Fauvel, Basurco and Saroglia2002).
In males the three major phases composing spermatogenesis were present in P. pollachius: mitotic proliferation of spermatogonia, meiosis of spermatocytes, and spermiogenesis, the restructuring of spermatids into flagellated spermatozoa (Schulz & Miura, Reference Schulz and Miura2002). Testicular tissue is of the lobular type (Takashima & Hibiya, Reference Takashima and Hibiya1995), i.e. spermatogonia are randomly distributed along the entire length of the lobule in reproductive active fish, which is equivalent to the unrestricted spermatogonial type (Grier, Reference Grier1980, Reference Grier1981). Spermatogenesis is cystic in this species, thus, developing germ cells are enclosed within germinal cysts formed by enveloping Sertoli cells.
This study represented the first approach in Atlantic Iberian waters to the annual reproductive cycle of P. pollachius. Some females with reproductive activity were detected from January to May, with a peak of spawning in February (more than 70% of mature females in active spawning condition), as confirmed by both histology and GSI mean values variation. In captive fish from wild populations, previous estimations of female spawning season are relatively far from our results, with estimated durations of 92 days at 9.5–11.8°C (Suquet et al., Reference Suquet, Petton, Normant, Dosdat and Gaignon1996), 57 days at 9.0–11.5°C (Omnes et al., Reference Omnes, Sévère, Barone, Suquet, Buchet, Le Roux, Gaignon, Fostier, Fauvel, Basurco and Saroglia2002) and 65 days at 8.0°C (Suquet et al., Reference Suquet, Normant, Gaignon, Quéméner and Fauvel2005). Our estimation of the spawning season duration (around 4 months) was shorter than that of other gadoids in Spanish waters, such as T. luscus (Alonso-Fernández et al., Reference Alonso-Fernández, Domínguez-Petit, Bao, Rivas and Saborido-Rey2008), but closer to other gadoids such as Norway pout Trisopterus esmarkii, poor cod Trisopterus minutus and whiting Merlangius merlangus, with reported spawning seasons of around 3–4 months (Cooper, Reference Cooper1983).
In spite of the observed differences in the mean GSI values of males and females, male reproductive cycle followed the same seasonality as females but extended until the end of May. Longer duration of male reproductive season may guarantee in some way the reproductive success of the species. This is the case of other exploited fish species like cod, G. morhua (Hutchings & Myers, Reference Hutchings and Myers1993), anglerfish, Lophiomus setigerus (Yoneda et al., Reference Yoneda, Tokimura, Fujita, Takeshita, Takeshita, Matsuyama and Matsuura1998) and winter flounder, Pleuronectes americanus (Sonya & Burton, Reference Sonya and Burton1999). An extended duration of spawning capable activity in males compared to those of females is usually a characteristic of batch spawning fish, in which spermatogenesis is a continuous process and testis never reach the regressing condition (Pavlov et al., Reference Pavlov, Emel'yanova, Novikov, Jakobse, Fogarty, Megrey and Moksness2009). Nevertheless, this is not exactly the case of P. pollachius since a relatively high proportion of regressing males were found just at the end of the spawning season. This is an evidence of ending of the spermatogenesis.
The highest levels of follicular atresia in females were found at the end of the spawning season, a typical feature of species with asynchronous development of secondary growth follicles (Murua & Saborido-Rey, Reference Murua and Saborido-Rey2003). Thus, the annual peak of prevalence of atresia seems to mark the end of the female spawning season in the population. This is concurrent with maximum levels of atresia in the regressing females, i.e. resorption of unreleased oocytes (Pavlov et al., Reference Pavlov, Emel'yanova, Novikov, Jakobse, Fogarty, Megrey and Moksness2009).
In ecological studies and in fishery science, body condition factor is widely used to account for the energy reserves stored within individual fish (Marshall et al., Reference Marshall, Yaragina, Lambert and Kjesbu1999). In some temperate species, condition factor fluctuates throughout the year with no clear seasonal pattern (Méndez & González, Reference Méndez and González1997; Domínguez-Petit et al., Reference Domínguez-Petit, Saborido-Rey and Medina2010). In the case of P. pollachius, the values observed along the year were quite variable but an increase was evident by the end of the spawning season in the values of HSI and K, likely related to active feeding during resting period. Population asynchronicity in spawning activity may make difficult the ecological study of the dynamic of energy allocation and mobilization in a temporal basis (Domínguez-Petit et al., Reference Domínguez-Petit, Saborido-Rey and Medina2010). Therefore, it was deemed convenient to analyse condition factor based on female reproductive phases, rather than based on a temporal pattern. In our study, only HSI followed a clear pattern of variation along the reproductive phases in females and males. This may be related to the role of the liver as a deposit of lipids and glycogen. Actually, the lowest HSI values were observed in individuals with fully developed gonads (actively spawning) whereas females with gonads at the beginning of the developmental process (developing) presented the highest values. This pattern may reflect the cycle of energy reserves and its relationship with gonad development and reproductive activity. The importance of liver lipid reserves in reproductive activity has also been described in other gadoid species such as G. morhua or T. luscus (Marshall et al., Reference Marshall, Yaragina, Lambert and Kjesbu1999; Alonso-Fernández & Saborido-Rey, Reference Alonso-Fernández and Saborido-Rey2012). In the case of P. pollachius additional effort is required to properly investigate the energy allocation strategy with biochemical data to better understand the relationship between energy reserves and reproduction.
Age/size at maturity is the most common and well-studied life history parameter in fish exploited populations (Trippel, Reference Trippel1995; Morgan, Reference Morgan2008), as it is used to inform on several stock assessment methods to get a better estimation of limit reference points. Traditionally, estimates of spawning stock biomass (SSB) assumed that all fish mature at the same age or use a single invariant maturity ogive (Morgan, Reference Morgan2008). However, the present study estimated significantly different L 50% for females (47.1 cm) and males (36.1 cm). Not considering these differences in future SSB estimations may have important consequences from a quantitative perspective (Morgan, Reference Morgan2008). In a previous study with captive individuals, Omnes et al. (Reference Omnes, Sévère, Barone, Suquet, Buchet, Le Roux, Gaignon, Fostier, Fauvel, Basurco and Saroglia2002) estimated an age at maturity of two years (around 0.7 kg) and three years (around 1.1 kg) for males and females, respectively. Although these previous studies were limited in sample size and captivity conditions are not comparable with wild conditions (i.e. continuous feeding in captivity), our results present similar trend especially for females (values in weight obtained from length–weight relationships, see Supplementary Material). L 50% is known to be a highly plastic parameter that changes under external pressure, due to anthropogenic or environmental factors (Trippel, Reference Trippel1995; Domínguez-Petit et al., Reference Domínguez-Petit, Korta, Saborido-Rey, Murua, Saínza and Piñeiro2008). Shifts in maturation at smaller sizes and younger ages can have important consequences on stock reproductive potential, affecting egg production (Kraus et al., Reference Kraus, Tomkiewicz and Köster2002) and egg and larval viability (Trippel et al., Reference Trippel, Kjesbu, Solemdal, Chamber and Trippel1997a). However, a long maturation data set is required to detect and validate those changes at a population level. To our knowledge there is no historical maturity data set available for P. pollachius in any stock. Worryingly, for a related exploited species such as P. virens, a dramatic decrease in L 50% of 16 cm for females and 9 cm for males was recorded in less than two decades in the NW Atlantic (Trippel et al., Reference Trippel, Morgan and Frechet1997b). Therefore, we strongly recommend monitoring this parameter to significantly improve the estimation of stock reproductive potential and its integration in stock assessment.
The present study sets the first reproductive knowledge for P. pollachius in Galician waters. This information will provide a baseline to improve the reproductive potential estimations and to assess the stock status. Further research should be focused on egg production estimations and age-based demographic analysis to complement present information and provide a better proxy of stock productivity.
ACKNOWLEDGEMENTS
The authors would like to thank to M. Muñoz (University of Gerona) for helpful histological advice. We are also deeply grateful to D. Domínguez, M. Fabeiro, M. Guiance and P. Varela for technical support and assistance with histological work and to F. Pinto for language revision.
FINANCIAL SUPPORT
This research was supported by the Xunta de Galicia, under the project ARTEVIGO (09MMA022402PR). David Villegas-Rios was funded by a FPU scholarship from the Ministry of Economy and Competitiveness.
Supplementary material and methods
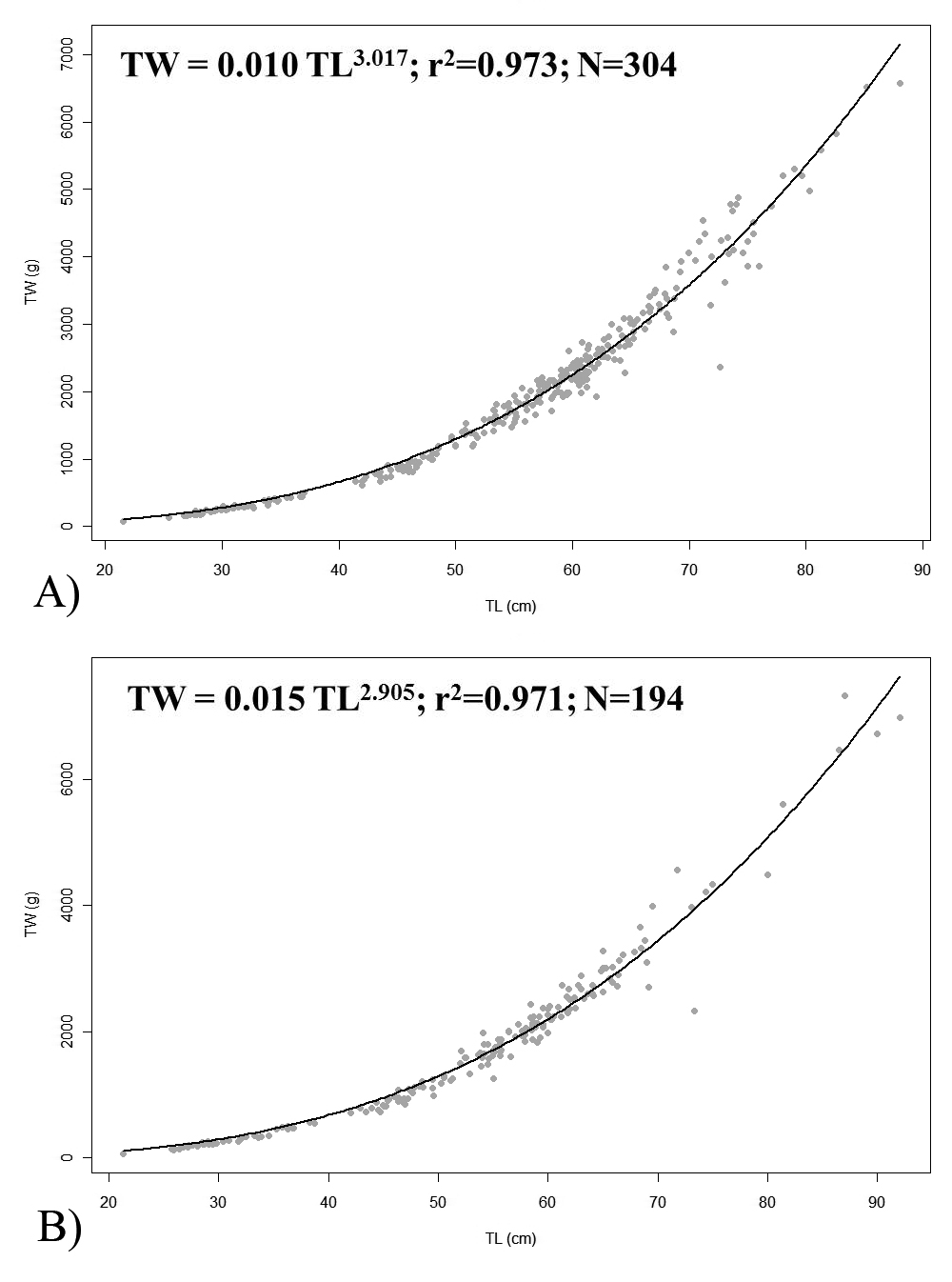
The following supplementary material, referred to in this paper, can be found online at journals.cambridge.org/mbi: length–weight relationships of (A) male and (B) female Pollachius pollachius from the present study sampling. Total weight is represented by TW (in g) and total length by TL (in cm).